
Nanomaterials and Coatings for Managing Antibiotic-Resistant Biofilms


Abstract
:1. Introduction
2. Anti-Virulence Approaches for Managing Biofilm Infections
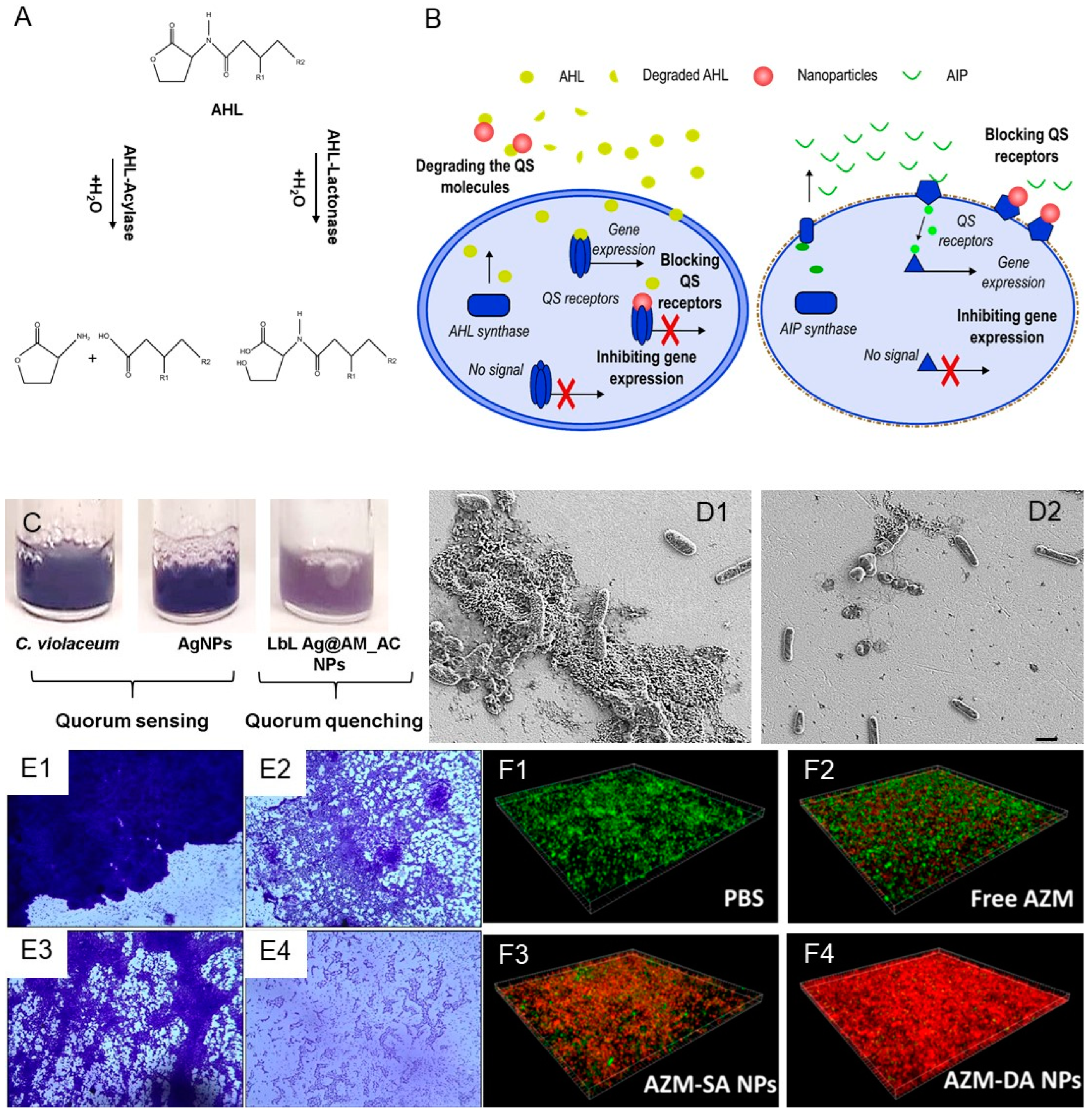
2.1. Nano-Formulated Quorum-Sensing Inhibitors
2.2. Quorum-Quenching Enzyme NPs
2.3. Metal NPs as QS Inhibitors
2.4. Mimicks of QS Signals and Inhibitors

3. Antifouling, Antiadhesive, and Biofilm-Dispersing Nanoactives
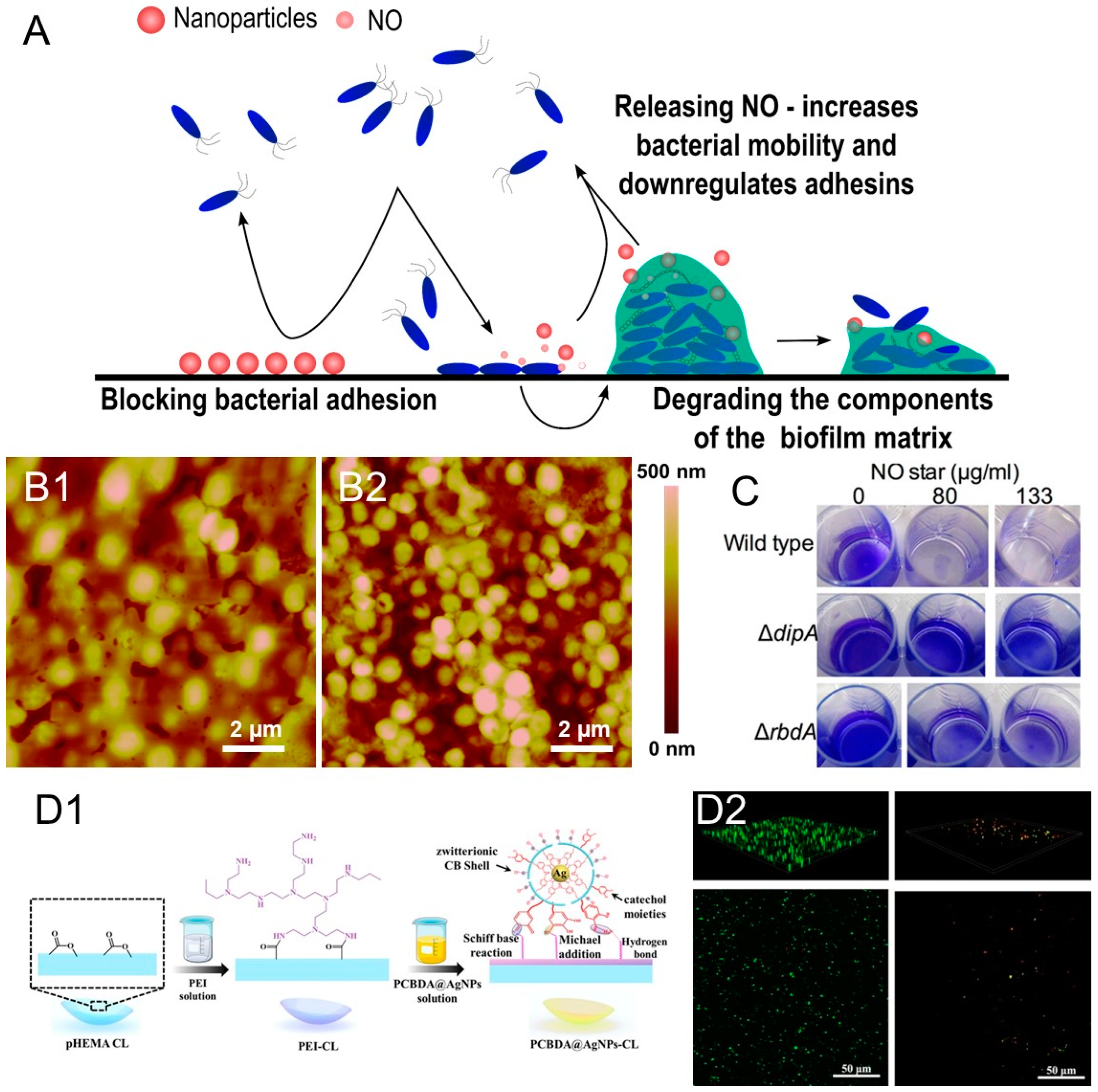
3.1. Nitric Oxide Donors Loaded in Nanocarriers
3.2. Zwitterionic Materials
3.3. Nanoformulated Matrix-Degrading Enzymes

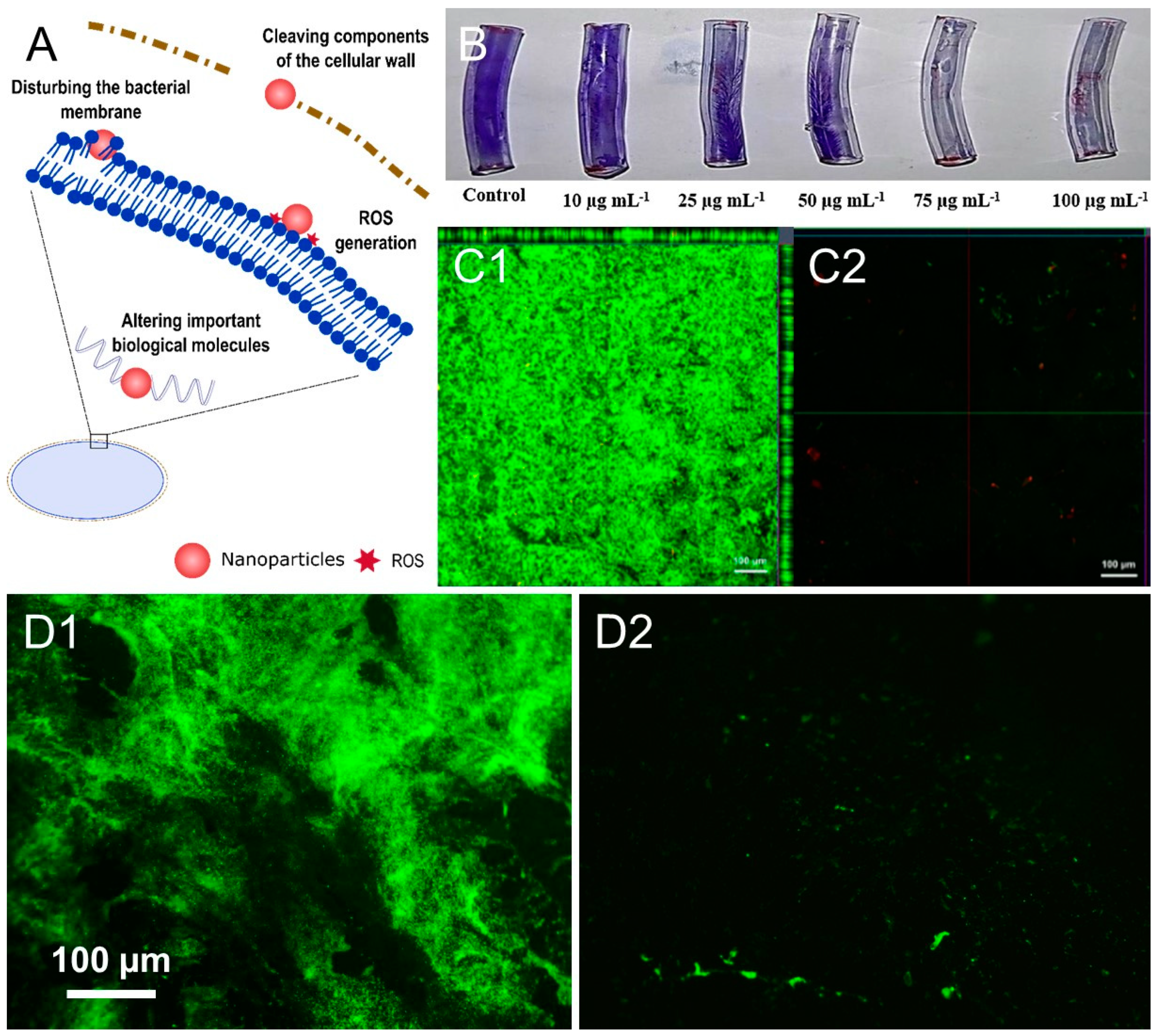
4. Antimicrobial Nanoactives
4.1. Antimicrobial Peptides
4.2. Non-Peptide Mimics of AMPs
4.3. Marine-Derived Antibacterial Lipids
4.4. Bactericidal Enzymes
4.5. Metal and Metal Oxide Nanoparticles


{kind=link}
{kind=link}
{kind=link}
| Strategy | Actives | Nanoparticle | Antibiofilm Activity | Application | Ref. |
|---|---|---|---|---|---|
| Quorum sensing inhibitors | Quorum sensing inhibitors | Gold-silica-cinnamaldehyde | S. aureus | Colloidal suspension | [38] |
| Acid-poly (lactic-co-glycolic acid)-azithromycin | P. aeruginosa | Colloidal suspension | [44] | ||
| Eugenol nanoemulsion | E. coli | Nanocomposite hydrogel | [48] | ||
| Quorum-quenching enzymes | Silver-aminocellulose-acylase | P. aeruginosa | Colloidal suspension | [33] | |
| Acylase-gentamicine | [28] | ||||
| Gold-lactonase | [56] | ||||
| Metal and metal oxides | Gold, nickel oxide, tellurium and selenium | P. aeruginosa | Colloidal suspension | [59,60,62] | |
| Mimics of QS machinery | CAI-1 (autoinducer) | V. cholera | Colloidal suspension | [64] | |
| Molecularly imprinted NPs | P. aeruginosa | [66] | |||
| Anti-adhesion | NO donors | NO-releasing silica NPs | P. aeruginosa, E. coli, S. aureus, S. epidermidis | Colloidal suspension | [70] |
| Polymeric stars-spermine | P. aeruginosa | Colloidal suspension | [71] | ||
| Silver-NO donor | P. aeruginosa, E. coli, S. aureus, K. pneumoniae | NP-containing PVA/PEG-films | [73] | ||
| Zwitterions | PSBMA-silver, PCBDA-silver | P. aeruginosa, E. coli, S. aureus | Membranes and gauzes coatings | [77,80] | |
| Matrix degrading enzymes | Silver-amylase | S. aureus, E. coli | Colloidal suspension | [34] | |
| Zinc oxide-amylase | Catheters coatings | [31] | |||
| Shellac NPs-protease | S. aureus | Colloidal suspension | [88] | ||
| Bactericidal | Peptides | RBRBR-chitosan, IVFK-silver | S. aureus, E. coli | Colloidal suspension | [95,96] |
| Polymyxin B-PSBMA | P. aeruginosa | Catheters coatings | [100] | ||
| Antibacterial enzymes | Lysozyme-magnetite-chitosan | S. aureus, P. aeruginosa | Colloidal suspension | [121] | |
| Lysozyme-polydopamine | E. coli | Hydrogel dressing | [122] | ||
| Cellobiose dehydrogenase | S. aureus | Surface coatings | [123] | ||
| Metal and metal oxides | Silver | S. aureus, E. coli, P. aeruginosa | Catheters and implants coatings | [125,126,127] | |
| Silver-chitosan | S. aureus, E. coli | Coatings | [30] | ||
| Selenium | S. aureus, P. aeruginosa and S. typhi | Colloidal suspension | [141] | ||
| Zinc oxide-chitosan-gallic acid | S. aureus | Contact lenses coatings | [149] |
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flemming, H.C.; Wingender, J.; Szewzyk, U.; Steinberg, P.; Rice, S.A.; Kjelleberg, S. Biofilms: An emergent form of bacterial life. Nat. Rev. Microbiol. 2016, 14, 563–575. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Inaba, T.; Kiyokawa, T.; Obana, N.; Yawata, Y.; Nomura, N. Environmental factors that shape biofilm formation. Biosci. Biotechnol. Biochem. 2016, 80, 7–12. [Google Scholar] [CrossRef] [PubMed]
- De Kievit, T.R.; Iglewski, B.H. Bacterial Quorum Sensing in Pathogenic Relationships. Infect. Immun. 2000, 68, 4839–4849. [Google Scholar] [CrossRef]
- Winstanley, C.; Fothergill, J.L. The role of quorum sensing in chronic cystic fibrosis Pseudomonas aeruginosa infections. FEMS Microbiol. Lett. 2009, 290, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Rand, J.D.; Danby, S.G.; Greenway, D.L.A.; England, R.R. Increased expression of the multidrug efflux genes acrAB occurs during slow growth of Escherichia coli. FEMS Microbiol. Lett. 2002, 207, 91–95. [Google Scholar] [CrossRef]
- Wright, G.D. Molecular mechanisms of antibiotic resistance. Chem. Comm. 2011, 47, 4055–4061. [Google Scholar] [CrossRef]
- Olsen, I. Biofilm-specific antibiotic tolerance and resistance. Eur. J. Clin. Microbiol. Infect. Dis. 2015, 34, 877–886. [Google Scholar] [CrossRef]
- Høiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef]
- Vuong, C.; Voyich, J.M.; Fischer, E.R.; Braughton, K.R.; Whitney, A.R.; Deleo, F.R.; Otto, M. Polysaccharide intercellular adhesin (PIA) protects Staphylococcus epidermidis against major components of the human innate immune system. Cell. Microbiol. 2004, 6, 269–275. [Google Scholar] [CrossRef]
- Malaekeh-nikouei, B.; Sedigheh, B.; Bazzaz, F.; Mirhadi, E.; Sadat, A.; Khameneh, B. The role of nanotechnology in combating biofilm-based antibiotic resistance. J. Drug Deliv. Sci. Technol. 2020, 60, 101880. [Google Scholar] [CrossRef]
- Madsen, J.S.; Burmølle, M.; Hansen, L.H.; Sørensen, S.J. The interconnection between biofilm formation and horizontal gene transfer. FEMS Immunol. Med. Microbiol. 2012, 65, 183–195. [Google Scholar] [CrossRef] [PubMed]
- Davies, D. Understanding Biofilm Resistance To Antibacterial Agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Macià, M.D.; del Pozo, J.L.; Díez-Aguilar, M.; Guinea, J. Microbiological diagnosis of biofilm-related infections. Enferm. Infecc. Microbiol. Clin. 2018, 36, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Elnagdy, S.; Raptopoulos, M.; Kormas, I.; Pedercini, A.; Wolff, L.F. Local Oral Delivery Agents withAnti-BiofilmProperties for the Treatment of Periodontitis and Peri-Implantitis. A Narrative Review. Molecules 2021, 26, 5661. [Google Scholar] [CrossRef] [PubMed]
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic treatment of biofilm infections. Apmis 2017, 125, 304–319. [Google Scholar] [CrossRef]
- Lynch, A.S.; Robertson, G.T. Bacterial and Fungal Biofilm Infections. Annu. Rev. Med. 2014, 59, 415–428. [Google Scholar] [CrossRef]
- Percival, S.L.; Suleman, L.; Vuotto, C.; Donelli, G. Healthcare-associated infections, medical devices and biofilms: Risk, tolerance and control. J. Med. Microbiol. 2015, 64, 323–334. [Google Scholar] [CrossRef]
- Guzella Carvalho, L.; Alfonso Alvim, M.M.; Luiz Fabri, R.; Morais Apolonio, A.C. Staphylococcus aureus biofilm formation in Minas Frescal cheese packaging. Dairy Technol. 2021, 74, 575–580. [Google Scholar] [CrossRef]
- Zimmermann, K.; Li, S.; Mohseni, M. Biological Activity Offsets Diffusion Limitations of Biofilms in Biological Ion Exchange Water Filters. ACS EST Water 2022, 2, 2441–2449. [Google Scholar] [CrossRef]
- Barravecchia, I.; DeCesari, C.; Pyankova, O.V.; Scebba, F.; Carlo, M.; Vecchione, A.; Tavanti, A.; Tedeschi, L.; Angeloni, D. Pitting Corrosion Within Bioreactors for Space Cell-Culture Contaminated by Paenibacillus glucanolyticus, a Case Report. Microgravity-Sci. Technol. 2018, 30, 309–319. [Google Scholar] [CrossRef]
- Stipanicev, M.; Turcu, F.; Esnault, L.; Schweitzer, E.W.; Kilian, R.; Basseguy, R. Corrosion behavior of carbon steel in presence of sulfate-reducing bacteria in seawater environment. Electrochim. Acta 2013, 113, 390–406. [Google Scholar] [CrossRef]
- Bockmühl, D.P. Laundry hygiene—How to get more than clean. J. Appl. Microbiol. 2017, 122, 1124–1133. [Google Scholar] [CrossRef] [PubMed]
- Clatworthy, A.E.; Pierson, E.; Hung, D.T. Targeting virulence: A new paradigm for antimicrobial therapy. Nat. Chem. Biol. 2007, 3, 541–548. [Google Scholar] [CrossRef]
- Wu, H.; Moser, C.; Wang, H.Z.; Høiby, N.; Song, Z.J. Strategies for combating bacterial biofilm infections. Int. J. Oral Sci. 2014, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Xia, G.; Shi, C.; Wan, J.; Liu, L.; Chen, Y.; Wu, Y.; Zhang, W.; Zhou, M.; He, H.; et al. Therapeutic strategies against bacterial biofilms. Fundam. Res. 2021, 1, 193–212. [Google Scholar] [CrossRef]
- Blaser, M. Stop the killing of beneficial bacteria. Nature 2011, 476, 393–394. [Google Scholar] [CrossRef] [PubMed]
- Rigo, S.; Cai, C.; Gunkel-Grabole, G.; Maurizi, L.; Zhang, X.; Xu, J.; Palivan, C.G. Nanoscience-Based Strategies to Engineer Antimicrobial Surfaces. Adv. Sci 2018, 5, 1700892. [Google Scholar] [CrossRef]
- Ivanova, K.; Ivanova, A.; Hoyo, J.; Pérez-Rafael, S.; Tzanov, T. Nano-Formulation Endows Quorum Quenching Enzyme-Antibiotic Hybrids with Improved Antibacterial and Antibiofilm Activities against Pseudomonas aeruginosa. Int. J. Mol. Sci. 2022, 23, 7632. [Google Scholar] [CrossRef]
- Zhang, L.; Gu, F.X.; Chan, J.M.; Wang, A.Z.; Langer, R.S.; Farokhzad, O.C. Nanoparticles in Medicine: Therapeutic Applications and Developments. Clin. Pharmacol. Ther. 2008, 83, 761–769. [Google Scholar] [CrossRef]
- Francesko, A.; Ivanova, K.; Hoyo, J.; Pérez-Rafael, S.; Petkova, P.; Fernandes, M.M.; Heinze, T.; Mendoza, E.; Tzanov, T. Bottom-up Layer-by-Layer Assembling of Antibacterial Freestanding Nanobiocomposite Films. Biomacromolecules 2018, 19, 3628–3636. [Google Scholar] [CrossRef]
- Ivanova, A.; Ivanova, K.; Perelshtein, I.; Gedanken, A.; Todorova, K.; Milcheva, R.; Dimitrov, P.; Popova, T.; Tzanov, T. Sonochemically engineered nano-enabled zinc oxide/amylase coatings prevent the occurrence of catheter-associated urinary tract infections. Mater. Sci. Eng. C 2021, 131, 112518. [Google Scholar] [CrossRef]
- Soule, L.D.; Chomorro, N.P.; Chuong, K.; Mellott, N.; Hammer, N.; Hankenson, K.D.; Chatzistavrou, X. Sol−Gel-Derived Bioactive and Antibacterial Multi-Component Thin Films by the Spin-Coating Technique. ACS Biomater. Sci. Eng. 2020, 5549–5562. [Google Scholar] [CrossRef]
- Ivanova, A.; Ivanova, K.; Tied, A.; Heinze, T.; Tzanov, T. Layer-By-Layer Coating of Aminocellulose and Quorum Quenching Acylase on Silver Nanoparticles Synergistically Eradicate Bacteria and Their Biofilms. Adv. Funct. Mater. 2020, 30, 2001284. [Google Scholar] [CrossRef]
- Ferreres, G.; Bassegoda, A.; Hoyo, J.; Torrent-Burgués, J.; Tzanov, T. Metal-Enzyme Nanoaggregates Eradicate Both Gram-Positive and Gram-Negative Bacteria and Their Biofilms. ACS Appl. Mater. Interfaces 2018, 10, 40434–40442. [Google Scholar] [CrossRef]
- Ivanova, K.; Ivanova, A.; Ramon, E.; Hoyo, J.; Sanchez-Gomez, S.; Tzanov, T. Antibody-Enabled Antimicrobial Nanocapsules for Selective Elimination of Staphylococcus aureus. ACS Appl. Mater. Interfaces 2020, 12, 35918–35927. [Google Scholar] [CrossRef]
- Da Silva Filho, P.M.; Andrade, A.L.; Cruz Lopes, J.B.A.; de Azevedo Pinheiro, A.; Alve de Vesconcelos, M.; da Cruz Fonseca, S.G.; de Lopes, F.L.G.; Silva Sousa, E.H.; Teixera, E.H.; Longhinotti, E. The biofilm inhibition activity of a NO donor nanosilica with enhanced antibiotics action. Int. J. Pharm. 2021, 610, 121220. [Google Scholar] [CrossRef]
- Niu, C.; Afre, S.; Gilbert, E.S. Subinhibitory concentrations of cinnamaldehyde interfere with quorum sensing. Lett. Appl. Microbiol. 2006, 43, 489–494. [Google Scholar] [CrossRef]
- Ramasamy, M.; Lee, J.; Jintae, L. Development of gold nanoparticles coated with silica containing the antibiofilm drug cinnamaldehyde and their effects on pathogenic bacteria. Int. J. Nanomed. 2017, 12, 2813–2828. [Google Scholar] [CrossRef]
- Subhaswaraj, P.; Barik, S.; Macha, C.; Venkata, P. Anti quorum sensing and anti biofilm efficacy of cinnamaldehyde encapsulated chitosan nanoparticles against Pseudomonas aeruginosa PAO1. LWT Food Sci. Technol. 2018, 97, 752–759. [Google Scholar] [CrossRef]
- Ouyang, J.; Sun, F.; Feng, W.; Sun, Y.; Qiu, X.; Xiong, L.; Liu, Y.; Chen, Y. Quercetin is an effective inhibitor of quorum sensing, biofilm formation and virulence factors in Pseudomonas aeruginosa. J. Appl. Microbiol. 2016, 120, 966–974. [Google Scholar] [CrossRef] [Green Version]
- Tran, T.; Hadinoto, K. A Potential Quorum-Sensing Inhibitor for Bronchiectasis Therapy: Quercetin—Chitosan Nanoparticle Complex Exhibiting Superior Inhibition of Biofilm Formation and Swimming Motility of Pseudomonas aeruginosa to the Native Quercetin. Int. J. Mol. Sci. 2021, 22, 1541. [Google Scholar] [CrossRef]
- Yu, L.; Shang, F.; Chen, X.; Ni, J.; Yu, L.; Zhang, M. The anti-biofilm effect of silver- nanoparticle-decorated quercetin nanoparticles on a multi-drug resistant Escherichia coli strain isolated from a dairy cow with mastitis. PeerJ 2018, 6, e5711. [Google Scholar] [CrossRef]
- Tateda, K.; Comte, R.; Pechere, J.-C.; Köhler, T.; Yamaguchi, K.; van Delden, C. Azithromycin Inhibits Quorum Sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2001, 45, 1930–1933. [Google Scholar] [CrossRef]
- Kłodzinska, S.N.; Wan, F.; Jumaa, H.; Sternberg, C.; Rades, T.; Nielsen, H.M. Utilizing nanoparticles for improving anti-biofilm effects of azithromycin: A head-to-head comparison of modified hyaluronic acid nanogels and coated poly (lactic-co-glycolic acid) nanoparticles. J. Colloid Interface Sci. 2019, 555, 595–606. [Google Scholar] [CrossRef]
- Gao, Y.; Wang, J.; Chai, M.; Li, X.; Deng, Y.; Jin, Q.; Ji, J. Size and Charge Adaptive Clustered Nanoparticles Targeting the Biofilm Microenvironment for Chronic Lung Infection Management. ACS Nano 2020, 14, 5686–5699. [Google Scholar] [CrossRef]
- Ashrafudoulla, M.; Rahaman, F.; Ha, A.J.; Hong, S.; Ha, S. Antibacterial and antibio film mechanism of eugenol against antibiotic resistance Vibrio parahaemolyticus. J. Food Microbiol. 2020, 91, 103500. [Google Scholar] [CrossRef]
- Lou, Z.; Letsididi, K.S.; Yu, F.; Pei, Z.; Wang, H. Inhibitive Effect of Eugenol and Its Nanoemulsion on Quorum Sensing–Mediated Virulence Factors and Biofilm Formation by Pseudomonas aeruginosa. J. Food Prot. 2019, 82, 379–389. [Google Scholar] [CrossRef]
- Prateeksha; Barik, S.K.; Singh, B.N. Nanoemulsion-loaded hydrogel coatings for inhibition of bacterial virulence and biofilm formation on solid surfaces. Sci. Rep. 2019, 9, 6520. [Google Scholar] [CrossRef]
- Antunes, J.C.; Tavares, T.D.; Teixeira, M.A.; Teixeira, M.O.; Homem, C.; Amorim, M.T.P.; Felgueiras, H.P. Eugenol-Containing Essential Oils Loaded onto Chitosan/Polyvinyl Alcohol Blended Films and Their Ability to Eradicate Staphylococcus aureus or Pseudomonas aeruginosa from Infected Microenvironments. Pharmaceutics 2021, 13, 195. [Google Scholar] [CrossRef]
- Nabawy, A.; Marie, J.; Schmidt-malan, S.; Park, J.; Li, C.; Huang, R.; Fedeli, S.; Nath, A.; Patel, R.; Rotello, V.M. Dual antimicrobial-loaded biodegradable nanoemulsions for synergistic treatment of wound biofilms. J. Control. Release 2022, 347, 379–388. [Google Scholar] [CrossRef]
- Sethuram, L.; Thomas, J.; Mukherjee, A.; Chandrasekaran, N. Eugenol micro-emulsion reinforced with silver nanocomposite electrospun mats for wound dressing strategies. Mater. Adv. 2021, 2, 2971–2988. [Google Scholar] [CrossRef]
- Sio, C.F.; Otten, L.G.; Cool, R.H.; Diggle, S.P.; Braun, P.G.; Bos, R.; Daykin, M.; Ca, M.; Williams, P.; Quax, W.J. Quorum Quenching by an N-Acyl-Homoserine Lactone Acylase from Pseudomonas aeruginosa PAO1. Infect. Immun. 2006, 74, 1673–1682. [Google Scholar] [CrossRef]
- Lee, B.; Yeon, K.; Shim, J.; Kim, S.; Lee, C.; Lee, J.; Kim, J. Effective Antifouling Using Quorum-Quenching Acylase Stabilized in Magnetically-Separable Mesoporous Silica. Biomacromolecules 2014, 15, 1153–1159. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, L.; Li, Q. A bioactive poly (vinylidene fluoride)/graphene oxide@acylase nanohybrid membrane: Enhanced anti-biofouling based on quorum quenching. J. Membr. Sci. 2018, 547, 110–122. [Google Scholar] [CrossRef]
- Anandan, K.; Rai, R. Microbial Pathogenesis Quorum quenching activity of AiiA lactonase KMMI17 from endophytic Bacillus thuringiensis KMCL07 on AHL-mediated pathogenic phenotype in Pseudomonas aeruginosa. Microb. Pthogenes. 2019, 132, 230–242. [Google Scholar] [CrossRef]
- Gupta, K.; Chhibber, S. Biofunctionalization of Silver Nanoparticles With Lactonase Leads to Altered Antimicrobial and Cytotoxic Properties. Front. Mol. Biosci. 2019, 6, 63. [Google Scholar] [CrossRef]
- Vinoj, G.; Pati, R.; Sonawane, A. In Vitro Cytotoxic Effects of Gold Nanoparticles Coated with Functional Acyl Homoserine Lactone Lactonase Protein from Bacillus licheniformis and Their Antibiofilm Activity against Proteus Species. Antimicrob Agents Chemother. 2014, 59, 763–771. [Google Scholar] [CrossRef]
- Singh, B.R.; Singh, B.N.; Singh, A.; Khan, W.; Naqvi, A.H. Mycofabricated biosilver nanoparticles interrupt Pseudomonas aeruginosa quorum sensing systems. Sci. Rep. 2015, 5, 13719. [Google Scholar] [CrossRef]
- Qais, F.A.; Ahmad, I.; Altaf, M.; Alotaibi, S.H. Biofabrication of Gold Nanoparticles Using Capsicum annuum Extract and Its Antiquorum Sensing and Antibio fi lm Activity against Bacterial Pathogens. ACS Omega 2021, 6, 16670–16682. [Google Scholar] [CrossRef]
- Maruthupandy, M.; Nadar, G.; Quero, F.; Li, W. Anti-quorum sensing and anti-biofilm activity of nickel oxide nanoparticles against Pseudomonas aeruginosa. J. Environ. Chem. Eng. 2020, 8, 104533. [Google Scholar] [CrossRef]
- Singh, N.; Paknikar, K.M.; Rajwade, J. Gene expression is influenced due to ‘nano’ and ‘ionic’ copper in pre-formed Pseudomonas aeruginosa biofilms. Environ. Res. 2019, 175, 367–375. [Google Scholar] [CrossRef]
- Gómez-Gómez, B.; Arregui, L.; Serrano, S.; Santos, A.; Pérez-Corona, T.; Madrid, Y. Selenium and tellurium-based nanoparticles as interfering factors in quorum sensing-regulated processes: Violacein production and bacterial biofilm formation. Metallomics 2019, 11, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Gómez-gómez, B.; Arregui, L.; Serrano, S.; Santos, A.; Pérez-corona, T.; Madrid, Y. Unravelling mechanisms of bacterial quorum sensing disruption by metal-based nanoparticles. Sci. Total Environ. 2019, 696, 133869. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.D.; Spiegel, A.C.; Hurley, A.; Perez, L.J.; Maisel, K.; Ensign, L.M.; Hanes, J.; Bassler, B.L.; Semmelhack, M.F.; Prud, R.K. Modulating Vibrio cholerae Quorum-Sensing-Controlled Communication Using Autoinducer-Loaded Nanoparticles. Nano Lett. 2015, 15, 2235–2241. [Google Scholar] [CrossRef]
- Piletska, E.V.; Stavroulakis, G.; Karim, K.; Whitcombe, M.J.; Chianella, I.; Sharma, A.; Eboigbodin, K.E.; Robinson, G.K.; Piletsky, S.A. Attenuation of vibrio fischeri quorum sensing using rationally designed polymers. Biomacromolecules 2010, 11, 975–980. [Google Scholar] [CrossRef]
- Lopez, J.G.; Piletska, E.V.; Whitcombe, M.J.; Czulak, J.; Piletsky, S.A. Application of molecularly imprinted polymer nanoparticles for degradation of the bacterial autoinducer N-hexanoyl homoserine lactone. Chem. Comm. 2019, 55, 2664–2667. [Google Scholar] [CrossRef] [PubMed]
- Fa, S.; Zhao, Y. Synthetic nanoparticles for selective hydrolysis of bacterial autoinducers in quorum sensing. Bioorg. Med. Chem. Lett. 2019, 29, 978–981. [Google Scholar] [CrossRef] [PubMed]
- Barraud, N.; Schleheck, D.; Klebensberger, J.; Webb, J.S.; Hassett, D.J.; Rice, S.A.; Kjelleberg, S. Nitric Oxide Signaling in Pseudomonas aeruginosa Biofilms Mediates Phosphodiesterase Activity, Decreased Cyclic Di-GMP Levels, and Enhanced Dispersal. J. Bacteriol. 2009, 191, 7333–7342. [Google Scholar] [CrossRef]
- Weller, R.B. Nitric oxide-containing nanoparticles as an antimicrobial agent and enhancer of wound healing. J. Investig. Dermatol. 2009, 129, 2335–2337. [Google Scholar] [CrossRef]
- Hetrick, E.M.; Shin, J.H.; Paul, H.S.; Schoenfisch, M.H. Anti-biofilm efficacy of nitric oxide-releasing silica nanoparticles. Biomaterials 2009, 30, 2782–2789. [Google Scholar] [CrossRef] [Green Version]
- Duong, H.T.T.; Jung, K.; Kutty, S.K.; Agustina, S.; Adnan, N.N.M.; Basuki, J.S.; Kumar, N.; Davis, T.P.; Barraud, N.; Boyer, C. Nanoparticle (Star Polymer) Delivery of Nitric Oxide Effectively Negates Pseudomonas aeruginosa Biofilm Formation. Biomacromolecules 2014, 15, 2583–2589. [Google Scholar] [CrossRef] [PubMed]
- Slomberg, D.L.; Lu, Y.; Broadnax, A.D.; Hunter, R.A.; Carpenter, A.W.; Schoen, M.H. Role of Size and Shape on Biofilm Eradication for Nitric Oxide-Releasing Silica Nanoparticles. ACS Appl. Mater. Interfaces 2013, 5, 9322–9329. [Google Scholar] [CrossRef] [PubMed]
- Rolim, W.R.; Pieretti, J.C.; Reno, L.S.; Lima, B.A.; Nascimento, M.H.M.; Ambrosio, F.N.; Lombello, C.B.; Brocchi, M.; De Souza, A.C.S.; Seabra, A.B. Antimicrobial Activity and Cytotoxicity to Tumor Cells of Nitric Oxide Donor and Silver Nanoparticles Containing PVA/PEG Films for Topical Applications. ACS Appl. Mater. Interfaces 2019, 11, 6589–6604. [Google Scholar] [CrossRef] [PubMed]
- Urzedo, A.L.; Gonçalves, M.C.; Nascimento, M.H.M.; Lombello, C.B.; Nakazato, G.; Seabra, A.B. Cytotoxicity and Antibacterial Activity of Alginate Hydrogel Containing Nitric Oxide Donor and Silver Nanoparticles for Topical Applications ̧. ACS Biomater. Sci. Eng. 2020, 6, 2117–2134. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Gao, K.; Zhou, L.; Jiao, Z.; Wu, M.; Cao, J.; You, X.; Cai, Z.; Su, Y.; Jiang, Z. Zwitterionic materials for antifouling membrane surface construction. Acta Biomater. 2016, 40, 142–152. [Google Scholar] [CrossRef]
- Neoh, K.G.; Li, M.; Kang, E.-T.; Chiong, E.; Tambyah, A. Surface modification strategies for combating catheter-related complications: Recent advances and challenges. J. Mater. Chem. B 2017, 5, 2045–2067. [Google Scholar] [CrossRef]
- Liu, C.; Faria, A.F.; Elimelech, M. Mitigation of Biofilm Development on Thin-Film Composite Membranes Functionalized with Zwitterionic Polymers and Silver Nanoparticles. Environ. Sci. Technol. 2017, 51, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Xin, X.; Li, P.; Zhu, Y.; Shi, L.; Yuan, J.; Shen, J. Mussel-Inspired Surface Functionalization of PET with Zwitterions and Silver Nanoparticles for the Dual-Enhanced Antifouling and Antibacterial Properties. Langmuir 2019, 35, 1788–1797. [Google Scholar] [CrossRef]
- Ma, L.; Li, K.; Xia, J.; Chen, C.; Liu, Y.; Lang, S.; Yu, L.; Liu, G. Commercial soft contact lenses engineered with zwitterionic silver nanoparticles for effectively treating microbial keratitis. J. Colloid Interface Sci. 2022, 610, 923–933. [Google Scholar] [CrossRef]
- Xiang, J.; Zhu, R.; Lang, S.; Yan, H.; Liu, G.; Peng, B. Mussel-inspired immobilization of zwitterionic silver nanoparticles toward antibacterial cotton gauze for promoting wound healing. Chem. Eng. J. 2021, 409, 128291. [Google Scholar] [CrossRef]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, K.; Fernandes, M.M.; Francesko, A.; Mendoza, E.; Guezguez, J.; Burnet, M.; Tzanov, T. Quorum-Quenching and Matrix-Degrading Enzymes in Multilayer Coatings Synergistically Prevent Bacterial Biofilm Formation on Urinary Catheters. ACS Appl. Mater. Interfaces 2015, 7, 27066–27077. [Google Scholar] [CrossRef] [PubMed]
- Abeleda, H.E.P.; Javier, A.P.; Murillo, A.Q.M.; Baculi, R.Q. Alpha-amylase conjugated biogenic silver nanoparticles as innovative strategy against biofilm-forming multidrug resistant bacteria. Biocatal. Agric. Biotechnol. 2020, 29, 101784. [Google Scholar] [CrossRef]
- Lamppa, J.W.; Griswold, K.E. Alginate Lyase Exhibits Catalysis-Independent Biofilm Dispersion and Antibiotic Synergy. Antimicrob. Agents Chemother. 2013, 57, 137–145. [Google Scholar] [CrossRef]
- Li, S.; Wang, Y.; Li, X.; Lee, B.S.; Jung, S.; Lee, M. Enhancing the Thermo-Stability and Anti-Biofilm Activity of Alginate Lyase by Immobilization on Low Molecular Weight Chitosan Nanoparticles. Int. J. Mol. Sci. 2019, 20, 4565. [Google Scholar] [CrossRef]
- Kumar, K.; Tripathi, M.; Pandey, N.; Kumar, A. Alginate lyase immobilized chitosan nanoparticles of ciprofloxacin for the improved antimicrobial activity against the biofilm associated mucoid P. aeruginosa infection in cystic fibrosis. Int. J. Pharm. 2019, 563, 30–42. [Google Scholar] [CrossRef]
- Wan, B.; Zhu, Y.; Tao, J.; Zhu, F.; Chen, J.; Li, L.; Zhao, J.; Wang, L.; Sun, S.; Yang, Y.; et al. Alginate Lyase Guided Silver Nanocomposites for Eradicating Pseudomonas aeruginosa from Lungs. ACS Appl. Mater. Interfaces 2020, 12, 9050–9061. [Google Scholar] [CrossRef]
- Weldrick, P.J.; Hardman, M.J. Smart active antibiotic nanocarriers with protease surface functionality can overcome biofilms of resistant bacteria. Mater. Chem. Front. 2021, 5, 961–972. [Google Scholar] [CrossRef]
- Weldrick, P.J.; Hardman, M.J.; Paunov, V.N. Enhanced Clearing of Wound-Related Pathogenic Bacterial Biofilms Using Protease-Functionalized Antibiotic Nanocarriers. ACS Appl. Mater. Interfaces 2019, 11, 43902–43919. [Google Scholar] [CrossRef]
- Li, W.; Geng, X.; Liu, D.; Li, Z. Near-Infrared Light-Enhanced Protease-Conjugated Gold Nanorods As A Photothermal Antimicrobial Agent For Elimination Of Exotoxin And Bio fi lms. Int. J. Nanomed. 2019, 14, 8047–8058. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Ashish, P.; Agrawal, K.; Anjum, M.; Tripathi, M.; Pandey, N. Dnase-I functionalization of ciprofloxacin-loaded chitosan nanoparticles overcomes the biofilm-mediated resistance of Pseudomonas aeruginosa. Appl. Nanosci. 2020, 10, 563–575. [Google Scholar] [CrossRef]
- Tasia, W.; Lei, C.; Cao, Y.; Ye, Q.; He, Y.; Xu, C. Enhanced eradication of bacterial biofilms with DNase I-loaded silver-doped mesoporous silica nanoparticles. Nanoscale 2020, 12, 2328. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.-Y.; Chou, W.; Lin, C.; Sung, C.T.; Alalaiwe, A.; Yang, S. Facile Biofilm Penetration of Cationic Liposomes Loaded with DNase I/Proteinase K to Eradicate Cutibacterium acnes for Treating Cutaneous and Catheter Infections. Int. J. Nanomedicine 2021, 16, 8121–8138. [Google Scholar] [CrossRef] [PubMed]
- Ivanova, K.; Ramon, E.; Hoyo, J. Innovative Approaches for Prevention of Clinically Relevant Biofilms: Current Trends and Future Prospects. Curr. Top. Med. Chem. 2017, 17, 1889–1914. [Google Scholar] [CrossRef]
- Almaaytah, A.; Mohammed, G.K.; Abualhaijaa, A.; Al-balas, Q. Development of novel ultrashort antimicrobial peptide nanoparticles with potent antimicrobial and antibiofilm activities against multidrug- resistant bacteria. Drug Des. Devel. Ther. 2017, 11, 3159–3170. [Google Scholar] [CrossRef]
- Seferji, K.A.; Susapto, H.H.; Khan, B.K.; Rheman, Z.U.; Abbas, M.; Emwas, A.; Hauser, C.A.E. Green Synthesis of Silver-Peptide Nanoparticles Generated by the Photoionization Process for Anti-Biofilm Application. ACS Appl. Bio Mater. 2021, 4, 8522–8535. [Google Scholar] [CrossRef]
- Zhang, P.; Wu, S.; Li, J.; Bu, X.; Dong, X.; Chen, N.; Li, F.; Zhu, J.; Sang, L.; Zeng, Y.; et al. Dual-sensitive antibacterial peptide nanoparticles prevent dental caries. Theranostics 2022, 12, 4818–4833. [Google Scholar] [CrossRef]
- Yu, Q.; Deng, T.; Lin, F.; Zhang, B.; Zink, J.I. Supramolecular Assemblies of Heterogeneous Mesoporous Silica Nanoparticles to Co-deliver Antimicrobial Peptides and Antibiotics for Synergistic Eradication of Pathogenic Biofilm. ACS Nano 2020, 14, 5926–5937. [Google Scholar] [CrossRef]
- Gupta, A.; Landis, R.F.; Li, C.; Schnurr, M.; Das, R.; Lee, Y.; Yazdani, M.; Liu, Y.; Kozlova, A.; Rotello, V.M. Engineered Polymer Nanoparticles with Unprecedented Antimicrobial Efficacy and Therapeutic Indices against Multidrug- Resistant Bacteria and Biofilms. JACS 2018, 140, 12137–12143. [Google Scholar] [CrossRef]
- Ivanova, A.; Ivanova, K.; Tzanov, T. Simultaneous Ultrasound-Assisted Hybrid Polyzwitterion/Antimicrobial Peptide Nanoparticles Synthesis and Deposition on Silicone Urinary Catheters for Prevention of Biofilm-Associated Infections. Nanomaterials 2021, 11, 3143. [Google Scholar] [CrossRef]
- Tan, P.; Fu, H.; Ma, X. Design, optimization, and nanotechnology of antimicrobial peptides: From exploration to applications. Nano Today 2021, 39, 101229. [Google Scholar] [CrossRef]
- Wnorowska, U.; Fiedoruk, K.; Piktel, E.; Prasad, S.V.; Sulik, M.; Janion, M.; Daniluk, T.; Savage, P.B.; Bucki, R. Nanoantibiotics containing membrane-active human cathelicidin LL-37 or synthetic ceragenins attached to the surface of magnetic nanoparticles as novel and innovative therapeutic tools: Current status and potential future applications. J. Nanobiotechnol. 2020, 18, 3. [Google Scholar] [CrossRef]
- Chmielewska, S.J.; Skłodowski, K.; Depciuch, J.; Deptuła, P.; Piktel, E.; Fiedoruk, K.; Kot, P.; Paprocka, P.; Fortunka, K.; Wollny, T.; et al. Bactericidal properties of rod-, peanut-, and star-shaped gold nanoparticles coated with ceragenin CSA-131 against multidrug-resistant bacterial strains. Pharmaceutics 2021, 13, 425. [Google Scholar] [CrossRef] [PubMed]
- Tokajuk, J.; Deptuła, P.; Chmielewska, S.J.; Skłodowski, K.; Mierzejewska, Ż.A.; Grądzka-Dahlke, M.; Tołstoj, A.; Daniluk, T.; Paprocka, P.; Savage, P.B.; et al. Ceragenin CSA-44 as a Means to Control the Formation of the Biofilm on the Surface of Tooth and Composite Fillings. Pathogens 2022, 11, 491. [Google Scholar] [CrossRef] [PubMed]
- Epand, R.F.; Pollard, J.E.; Wright, J.O.; Savage, P.B.; Epand, R.M. Depolarization, bacterial membrane composition, and the antimicrobial action of ceragenins. Antimicrob. Agents Chemother. 2010, 54, 3708–3713. [Google Scholar] [CrossRef] [PubMed]
- Paprocka, P.; Durnaś, B.; Mańkowska, A.; Skłodowski, K.; Król, G.; Zakrzewska, M.; Czarnowski, M.; Kot, P.; Fortunka, K.; Góźdź, S.; et al. New β-Lactam Antibiotics and Ceragenins—A Study to Assess Their Potential in Treatment of Infections Caused by Multidrug-Resistant Strains of Pseudomonas aeruginosa. Infect. Drug Resist. 2021, 14, 5681–5698. [Google Scholar] [CrossRef] [PubMed]
- Paprocka, P.; Mańkowska, A.; Skłodowski, K.; Król, G.; Wollny, T.; Lesiak, A.; Głuszek, K.; Savage, P.B.; Durnaś, B.; Bucki, R. Bactericidal Activity of Ceragenin in Combination with Ceftazidime, Levofloxacin, Co-Trimoxazole, and Colistin against the Opportunistic Pathogen Stenotrophomonas maltophilia. Pathogens 2022, 11, 621. [Google Scholar] [CrossRef]
- Hoppens, M.A.; Sylvester, C.B.; Qureshi, A.T.; Scherr, T.; Czapski, D.R.; Duran, R.S.; Savage, P.B.; Hayes, D. Ceragenin mediated selectivity of antimicrobial silver nanoparticles. ACS Appl. Mater. Interfaces 2014, 6, 13900–13908. [Google Scholar] [CrossRef]
- Nagant, C.; Pitts, B.; Stewart, P.S.; Feng, Y.; Savage, P.B.; Dehaye, J.P. Study of the effect of antimicrobial peptide mimic, CSA-13, on an established biofilm formed by Pseudomonas aeruginosa. Microbiologyopen 2013, 2, 318–325. [Google Scholar] [CrossRef]
- Niemirowicz, K.; Durnaś, B.; Tokajuk, G.; Piktel, E.; Michalak, G.; Gu, X.; Kułakowska, A.; Savage, P.B.; Bucki, R. Formulation and candidacidal activity of magnetic nanoparticles coated with cathelicidin LL-37 and ceragenin CSA-13. Sci. Rep. 2017, 7, 4610. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Jennings, J.D.; Snarr, J.; Chaudhary, V.; Pollard, J.E.; Savage, P.B. Optimization of ceragenins for prevention of bacterial colonization of hydrogel contact lenses. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6217–6223. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.L.; Sinclair, K.D.; Jeyapalina, S.; Bloebaum, R.D. Characterization of a novel active release coating to prevent biofilm implant-related infections. J. Biomed. Mater. Res. Part B Appl. Biomater. 2013, 101 B, 1078–1089. [Google Scholar] [CrossRef]
- Hashemi, M.M.; Holden, B.S.; Taylor, M.F.; Wilson, J.; Coburn, J.; Hilton, B.; Nance, T.; Gubler, S.; Genberg, C.; Deng, S.; et al. Antibacterial and antifungal activities of poloxamer micelles containing ceragenin CSA-131 on ciliated tissues. Molecules 2018, 23, 596. [Google Scholar] [CrossRef] [PubMed]
- Stefanović, O.D. Synergistic activity of antibiotics and bioactive plant extracts: A study against gram-positive and gram-negative bacteria. In Bacterial Pathogenesis and Antibacterial Control; IntechOpen: London, UK, 2018; pp. 23–48. [Google Scholar]
- Alves, E.; Dias, M.; Lopes, D.; Almeida, A.; Domingues, M.D.R.; Rey, F. Antimicrobial lipids from plants and marine organisms: An overview of the current state-of-the-art and future prospects. Antibiotics 2020, 9, 441. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Lawlor, K.C. Antibacterial activity of long-chain polyunsaturated fatty acids against Propionibacterium acnes and Staphylococcus aureus. Mar. Drugs 2013, 11, 4544–4557. [Google Scholar] [CrossRef] [PubMed]
- Cheow, W.S.; Chang, M.W.; Hadinoto, K. The roles of lipid in anti-biofilm efficacy of lipid-polymer hybrid nanoparticles encapsulating antibiotics. Colloids Surfaces A Physicochem. Eng. Asp. 2011, 389, 158–165. [Google Scholar] [CrossRef]
- Fazly Bazzaz, B.S.; Khameneh, B.; Zarei, H.; Golmohammadzadeh, S. Antibacterial efficacy of rifampin loaded solid lipid nanoparticles against Staphylococcus epidermidis biofilm. Microb. Pathog. 2016, 93, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Rozenbaum, R.T.; Su, L.; Umerska, A.; Eveillard, M.; Håkansson, J.; Mahlapuu, M.; Huang, F.; Liu, J.; Zhang, Z.; Shi, L.; et al. Antimicrobial synergy of monolaurin lipid nanocapsules with adsorbed antimicrobial peptides against Staphylococcus aureus biofilms in vitro is absent in vivo. J. Control. Release 2019, 293, 73–83. [Google Scholar] [CrossRef]
- Xu, C.; He, Y.; Li, Z.; Nor, Y.A.; Ye, Q. Nanoengineered hollow mesoporous silica nanoparticles for the delivery of antimicrobial proteins into biofilms. J. Mater. Chem. B 2018, 6, 1899–1902. [Google Scholar] [CrossRef]
- Spirescu, V.A.; Niculescu, A.; Slave, S.; Birca, A.C.; Dorcioman, G.; Grumezescu, V.; Holban, A.M.; Oprea, O.-C.; Vasile, B.S.; Grumezescu, A.M.; et al. Anti-Biofilm Coatings Based on Chitosan and Lysozyme Functionalized Magnetite Nanoparticles. antibiotics 2021, 10, 1269. [Google Scholar] [CrossRef]
- Wang, Y.; Lv, Q.; Chen, Y.; Xu, L.; Feng, M.; Xiong, Z.; Li, J.; Ren, J.; Liu, J.; Liu, B. Bilayer hydrogel dressing with lysozyme- enhanced photothermal therapy for biofilm eradication and accelerated chronic wound repair. Acta Pharm. Sin. B 2022. [Google Scholar] [CrossRef]
- Lipovsky, A.; Thallinger, B.; Perelshtein, I.; Ludwig, R.; Sygmund, C.; Nyanhongo, G.S.; Guebitz, G.M.; Gedanken, A. Ultrasound coating of polydimethylsiloxanes with antimicrobial enzymes. J. Mater. Chem. B 2015, 3, 7014–7019. [Google Scholar] [CrossRef]
- Tan, Y.; Ma, S.; Leonhard, M.; Moser, D.; Ludwig, R. Co-immobilization of cellobiose dehydrogenase and deoxyribonuclease I on chitosan nanoparticles against fungal/bacterial polymicrobial biofilms targeting both biofilm matrix and microorganisms. Mater. Sci. Eng. C 2020, 108, 110499. [Google Scholar] [CrossRef] [PubMed]
- Roe, D.; Karandikar, B.; Bonn-savage, N.; Gibbins, B.; Roullet, J. Antimicrobial surface functionalization of plastic catheters by silver nanoparticles. J. Antimicrob. Chemother. 2008, 61, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Lewisoscar, F.; Nithya, C.; Vismaya, S.; Arunkumar, M.; Thajuddin, N. In vitro analysis of green fabricated silver nanoparticles (AgNPs) against Pseudomonas aeruginosa PA14 biofilm formation, their application on urinary catheter. Prog. Org. Coat. 2021, 151, 106058. [Google Scholar] [CrossRef]
- Zepon, K.M.; Souza, M.; Witt, A.; Fetzner, A.; Dal, F.; Morisso, P.; Luiza, A.; Heriberto, J.; Faverzani, R.; Alberto, L. Polymer-based wafers containing in situ synthesized gold nanoparticles as a potential wound-dressing material. Mater. Sci. Eng. C 2020, 109, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Van Hengel, I.A.J.; Riool, M.; Fratila-apachitei, L.E.; Witte-bouma, J.; Farrell, E.; Zadpoor, A.A.; Zaat, S.A.J.; Apachitei, I. Selective laser melting porous metallic implants with immobilized silver nanoparticles kill and prevent bio fi lm formation by methicillin- resistant Staphylococcus aureus. Biomaterials 2017, 140, 1–15. [Google Scholar] [CrossRef]
- Tran, H.A.; Tran, P.A. In Situ Coatings of Silver Nanoparticles for Biofilm Treatment in Implant-Retention Surgeries: Antimicrobial Activities in Monoculture and Coculture. ACS Appl. Mater. Interfaces 2021, 13, 41435–41444. [Google Scholar] [CrossRef]
- Wickramasinghe, S.; Ju, M.; Milbrandt, N.B.; Tsai, Y.H.; Navarreto-lugo, M.; Visperas, A.; Klika, A.; Barsoum, W.; Higuera-rueda, C.A.; Samia, A.C.S. Photoactivated Gold Nanorod Hydrogel Composite Containing. ACS Appl. Nano Mater. 2020, 3, 5862–5873. [Google Scholar] [CrossRef]
- Abdulkareem, E.H.; Memarzadeh, K.; Allaker, R.P.; Huang, J.; Pratten, J.; Spratt, D. Anti-biofilm activity of zinc oxide and hydroxyapatite nanoparticles as dental implant coating materials. J. Dent. 2015, 43, 1462–1469. [Google Scholar] [CrossRef]
- Taylor, P.; Kulshrestha, S.; Khan, S.; Meena, R.; Singh, B.R.; Khan, A.U. A graphene/zinc oxide nanocomposite film protects dental implant surfaces against cariogenic Streptococcus mutans. Biofouling 2014, 30, 37–41. [Google Scholar] [CrossRef]
- Neethu, S.; Midhun, S.J.; Radhakrishnan, E.K.; Jyothis, M. Surface functionalization of central venous catheter with mycofabricated silver nanoparticles and its antibio fi lm activity on multidrug resistant Acinetobacter baumannii. Microb. Pthogenes. 2020, 138, 103832. [Google Scholar] [CrossRef] [PubMed]
- Kumari, P.; Bharti, A.; Agarwal, V.; Kumar, N. ZnO coating on silicon rubber urinary catheter to reduce the biofilm infections. Adv. Mater. Process. Technol. 2021, 8, 423–433. [Google Scholar] [CrossRef]
- Ballo, M.K.S.; Rtimi, S.; Pulgarin, C.; Hopf, N.; Berthet, A.; Kiwi, J.; Moreillon, P.; Entenza, J.M. In Vitro and In Vivo Effectiveness of an Innovative Silver-Copper Nanoparticle Coating of Catheters To Prevent Methicillin-Resistant Staphylococcus aureus Infection. Antimicrob. Agents Chemother. 2016, 60, 5349–5356. [Google Scholar] [CrossRef]
- Pérez-díaz, M.; Alvarado-gomez, E.; Magaña-aquino, M.; Sánchez-sánchez, R.; Velasquillo, C.; Gonzalez, C. Anti-biofilm activity of chitosan gels formulated with silver nanoparticles and their cytotoxic effect on human fi broblasts. Mater. Sci. Eng. C 2016, 60, 317–323. [Google Scholar] [CrossRef]
- Haidari, H.; Bright, R.; Garg, S.; Vasilev, K.; Cowin, A.J.; Kopecki, Z. Eradication of Mature Bacterial Biofilms with Concurrent Improvement in Chronic Wound Healing Using Silver Nanoparticle Hydrogel Treatment. Biomedicines 2021, 9, 1182. [Google Scholar] [CrossRef]
- Velázquez-velázquez, J.L.; Santos-flores, A.; Araujo-meléndez, J.; Sánchez-sánchez, R.; Velasquillo, C.; González, C.; Martínez-castañon, G.; Martinez-gutierrez, F. Anti-biofilm and cytotoxicity activity of impregnated dressings with silver nanoparticles. Mater. Sci. Eng. C 2015, 49, 604–611. [Google Scholar] [CrossRef]
- Khan, R.; Huang, K.; Jinzhong, Z.; Zhu, T. Exploration of the antibacterial and wound healing potential of a PLGA/silk fibroin based electrospun membrane loaded with zinc oxide nanoparticles. J. Mater. Chem. B 2021, 9, 1452–1465. [Google Scholar] [CrossRef]
- Gong, C.; Luo, Y.; Pan, Y. Novel synthesized zinc oxide nanoparticles loaded alginate-chitosan biofilm to enhanced wound site activity and anti-septic abilities for the management of complicated abdominal wound dehiscence. J. Photochem. Photobiol. B Biol. 2019, 192, 124–130. [Google Scholar] [CrossRef]
- Ullah, A.; Mirani, Z.A.; Binbin, S.; Wang, F.; Chan, M.W.H.; Aslam, S.; Yonghong, L.; Hasan, N.; Naveed, M.; Hussain, S.; et al. An elucidative study of the anti-biofilm effect of selenium nanoparticles (SeNPs) on selected biofilm producing pathogenic bacteria: A disintegrating effect of SeNPs on bacteria. Process Biochem. 2023, 126, 98–107. [Google Scholar] [CrossRef]
- Mirza, K.; Naaz, F.; Ahmad, T.; Manzoor, N.; Sardar, M. Development of Cost-Effective, Ecofriendly Selenium Nanoparticle-Functionalized Cotton Fabric for Antimicrobial and Antibiofilm Activity. Fermentation 2023, 9, 18. [Google Scholar] [CrossRef]
- Taylor, P.; Lewisoscar, F.; Mubarakali, D. One pot synthesis and anti-biofilm potential of copper nanoparticles ( CuNPs ) against clinical strains of Pseudomonas aeruginosa. Biofouling 2015, 31, 371–391. [Google Scholar] [CrossRef]
- Altaf, M.; Zeyad, T.; Hashmi, A.; Abdulaziz, M.; Almuzaini, M. Efective inhibition and eradication of pathogenic biofilms by titanium dioxide nanoparticles synthesized using Carum copticum extract. RSC Adv. 2021, 11, 19248–19257. [Google Scholar] [CrossRef] [PubMed]
- Rajivgandhi, G.; Ramachandran, G.; Chackaravarthi, G.; Kanisha, C.; Maruthupandy, M.; Quero, F.; Al-mekhlafi, F.A.; Wadaan, M.A.; Li, W. Preparation of antibacterial Zn and Ni substituted cobalt ferrite nanoparticles for efficient biofilm eradication. Anal. Biochem. 2022, 653, 114787. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.T.; Ahamed, M.; Al-khedhairy, A.; Musarrat, J. Biocidal effect of copper and zinc oxide nanoparticles on human oral microbiome and biofilm formation. Mater. Lett. 2013, 97, 67–70. [Google Scholar] [CrossRef]
- Seo, Y.; Hwang, J.; Lee, E.; Kim, Y.J.; Lee, K.; Park, C.; Choi, Y.; Jeon, H.; Choi, J. Engineering copper nanoparticles synthesized on the surface of carbon nanotubes for anti-microbial and anti-biofilm applications. Nanoscale 2018, 10, 15529–15544. [Google Scholar] [CrossRef]
- Salat, M.; Petkova, P.; Hoyo, J.; Perelshtein, I.; Gedanken, A.; Tzanov, T. Durable antimicrobial cotton textiles coated sonochemically with ZnO nanoparticles embedded in an in-situ enzymatically generated bioadhesive. Carbohydr. Polym. 2018, 189, 198–203. [Google Scholar] [CrossRef]
- Hoyo, J.; Ivanova, K.; Guaus, E.; Tzanov, T. Multifunctional ZnO NPs-chitosan-gallic acid hybrid nanocoating to overcome contact lenses associated conditions and discomfort. J. Colloid Interface Sci. 2019, 543, 114–121. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreres, G.; Ivanova, K.; Ivanov, I.; Tzanov, T. Nanomaterials and Coatings for Managing Antibiotic-Resistant Biofilms. Antibiotics 2023, 12, 310. https://doi.org/10.3390/antibiotics12020310
Ferreres G, Ivanova K, Ivanov I, Tzanov T. Nanomaterials and Coatings for Managing Antibiotic-Resistant Biofilms. Antibiotics. 2023; 12(2):310. https://doi.org/10.3390/antibiotics12020310
Chicago/Turabian StyleFerreres, Guillem, Kristina Ivanova, Ivan Ivanov, and Tzanko Tzanov. 2023. "Nanomaterials and Coatings for Managing Antibiotic-Resistant Biofilms" Antibiotics 12, no. 2: 310. https://doi.org/10.3390/antibiotics12020310
APA StyleFerreres, G., Ivanova, K., Ivanov, I., & Tzanov, T. (2023). Nanomaterials and Coatings for Managing Antibiotic-Resistant Biofilms. Antibiotics, 12(2), 310. https://doi.org/10.3390/antibiotics12020310