
Induction of Viable but Non-Culturable State in Clinically Relevant Staphylococci and Their Detection with Bacteriophage K

Abstract
:1. Introduction
2. Results
2.1. Biofilm Formation and Induction of VBNC State
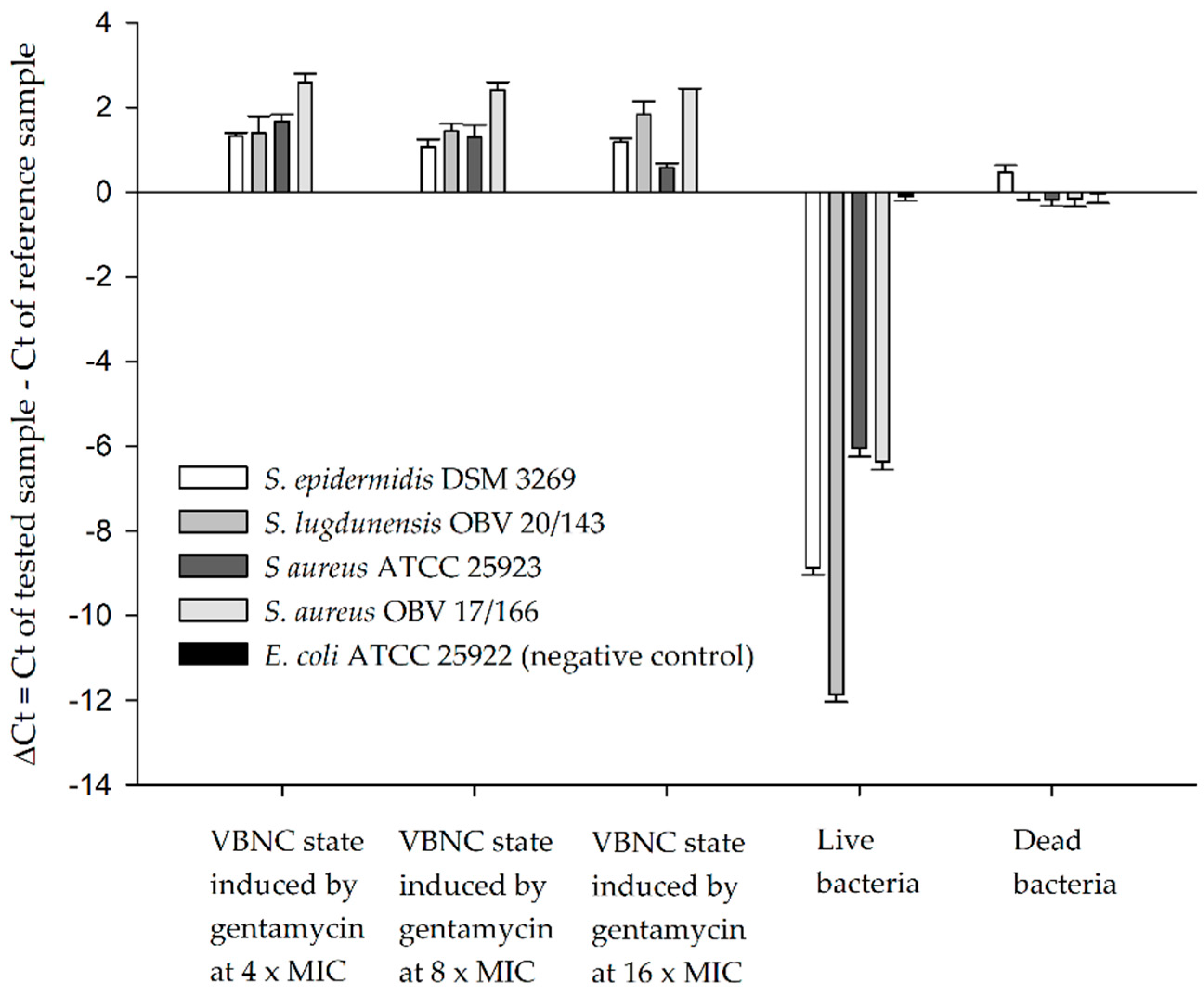
2.2. Detection of VBNC Bacteria with Bacteriophages and qPCR
3. Discussion
4. Materials and Methods
4.1. Bacteria and Bacteriophages
4.2. Biofilm Formation and Induction of VBNC State
4.3. Detection of VBNC Bacteria with Bacteriophages and qPCR
4.4. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abdelsattar, A.; Dawoud, A.; Makky, S.; Nofal, R.; Aziz, R.; El-Shibiny, A. Bacteriophages: From Isolation to Application. Curr. Pharm. Biotechnol. 2022, 23, 337–360. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Ross, R.P.; Meaney, W.; Fitzgerald, G.F.; Elbreki, M.F.; Coffey, A. Potential of the Polyvalent Anti-Staphylococcus Bacteriophage K for Control of Antibiotic-Resistant Staphylococci from Hospitals. Appl. Environ. Microbiol. 2005, 71, 1836–1842. [Google Scholar] [CrossRef]
- Estrella, L.A.; Quinones, J.; Henry, M.; Hannah, R.M.; Pope, R.K.; Hamilton, T.; Teneza-mora, N.; Hall, E.; Biswajit, B. Characterization of Novel Staphylococcus aureus Lytic Phage and Defining Their Combinatorial Virulence Using the OmniLog ® System. Bacteriophage 2016, 6, e1219440. [Google Scholar] [CrossRef]
- Šuster, K.; Podgornik, A.; Cör, A. Quick Bacteriophage-Mediated Bioluminescence Assay for Detecting Staphylococcus spp. in Sonicate Fluid of Orthopaedic Artificial Joints. New Microbiol 2017, 40, 190–196. [Google Scholar] [PubMed]
- Plota, M.; Sazakli, E.; Giormezis, N.; Gkartziou, F.; Kolonitsiou, F.; Leotsinidis, M.; Antimisiaris, S.G.; Spiliopoulou, I. In Vitro Anti-Biofilm Activity of Bacteriophage K (ATCC 19685-B1) and Daptomycin against Staphylococci. Microorganisms 2021, 9, 1853. [Google Scholar] [CrossRef] [PubMed]
- Šuster, K.; Podgornik, A.; Cör, A. An Alternative Molecular Approach for a Rapid and Specific Detection of Clinically Relevant Bacteria Causing Prosthetic Joint Infections with Bacteriophage K. New Microbiol. 2020, 43, 107–114. [Google Scholar]
- Šuster, K.; Cör, A. Fast and Specific Detection of Staphylococcal PJI with Bacteriophage-based Methods within 104 Sonicate Fluid Samples. J. Orthop. Res. 2022, 40, 1358–1364. [Google Scholar] [CrossRef] [PubMed]
- Goodman, S.B.; Gallo, J.; Gibon, E.; Takagi, M. Diagnosis and Management of Implant Debris-Associated Inflammation. Expert Rev. Med. Devices 2020, 17, 41–56. [Google Scholar] [CrossRef]
- Esteban, J.; Pérez-Jorge, C.; Pérez-Tanoira, R.; Gómez-Barrena, E. Microbiological Diagnosis of Prosthetic Joint Infection. In Infected Total Joint Arthroplasty: The Algorithmic Approach; Trebše, R., Ed.; Springer: London, UK, 2012; pp. 165–179. [Google Scholar]
- Berbari, E.F.; Marculescu, C.; Sia, I.; Lahr, B.D.; Hanssen, A.D.; Steckelberg, J.M.; Gullerud, R.; Osmon, D.R. Culture-Negative Prosthetic Joint Infection. Clin. Infect. Dis. 2007, 45, 1113–1119. [Google Scholar] [CrossRef]
- Malekzadeh, D.; Osmon, D.R.; Lahr, B.D.; Hanssen, A.D.; Berbari, E.F. Prior Use of Antimicrobial Therapy Is a Risk Factor for Culture-Negative Prosthetic Joint Infection. Clin. Orthop. 2010, 468, 2039–2045. [Google Scholar] [CrossRef]
- Kim, Y.-H.; Park, J.-W.; Kim, J.-S.; Kim, D.-J. The Outcome of Infected Total Knee Arthroplasty: Culture-Positive Versus Culture-Negative. Arch. Orthop. Trauma Surg. 2015, 135, 1459–1467. [Google Scholar] [CrossRef]
- Kalbian, I.; Park, J.W.; Goswami, K.; Lee, Y.-K.; Parvizi, J.; Koo, K.-H. Culture-Negative Periprosthetic Joint Infection: Prevalence, Aetiology, Evaluation, Recommendations, and Treatment. Int. Orthop. 2020, 44, 1255–1261. [Google Scholar] [CrossRef]
- Ayrapetyan, M.; Williams, T.; Oliver, J.D. Relationship between the Viable but Nonculturable State and Antibiotic Persister Cells. J. Bacteriol. 2018, 200, e00249-18. [Google Scholar] [CrossRef]
- Charani, E.; McKee, M.; Ahmad, R.; Balasegaram, M.; Bonaconsa, C.; Merrett, G.B.; Busse, R.; Carter, V.; Castro-Sanchez, E.; Franklin, B.D.; et al. Optimising Antimicrobial Use in Humans—Review of Current Evidence and an Interdisciplinary Consensus on Key Priorities for Research. Lancet Reg. Health Eur. 2021, 7, 100161. [Google Scholar] [CrossRef]
- Stewart, P.S.; Franklin, M.J. Physiological Heterogeneity in Biofilms. Nat. Rev. Microbiol. 2008, 6, 199–210. [Google Scholar] [CrossRef]
- Ayrapetyan, M.; Williams, T.C.; Oliver, J.D. Bridging the Gap between Viable but Non-Culturable and Antibiotic Persistent Bacteria. Trends Microbiol. 2015, 23, 7–13. [Google Scholar] [CrossRef]
- Pasquaroli, S.; Zandri, G.; Vignaroli, C.; Vuotto, C.; Donelli, G.; Biavasco, F. Antibiotic Pressure Can Induce the Viable but Non-Culturable State in Staphylococcus Aureus Growing in Biofilms. J. Antimicrob. Chemother. 2013, 68, 1812–1817. [Google Scholar] [CrossRef]
- Pasquaroli, S.; Citterio, B.; Cesare, A.; Amiri, M.; Manti, A.; Vuotto, C.; Biavasco, F. Role of Daptomycin in the Induction and Persistence of the Viable but Non-Culturable State of Staphylococcus Aureus Biofilms. Pathogens 2014, 3, 759–768. [Google Scholar] [CrossRef]
- Ramamurthy, T.; Ghosh, A.; Pazhani, G.P.; Shinoda, S. Current Perspectives on Viable but Non-Culturable (VBNC) Pathogenic Bacteria. Front. Public Health 2014, 2, 103. [Google Scholar] [CrossRef]
- Dong, K.; Pan, H.; Yang, D.; Rao, L.; Zhao, L.; Wang, Y.; Liao, X. Induction, Detection, Formation, and Resuscitation of Viable but Non-Culturable State Microorganisms. Compr. Rev. Food Sci. Food Saf. 2020, 19, 149–183. [Google Scholar] [CrossRef]
- Hyman, P.; Abedon, S.T. Practical Methods for Determining Phage Growth Parameters. In Bacteriophages: Methods and Protocols, Volume 1: Isolation, Characterization, and Interactions; Clokie, M.R.J., Kropinski, A.M., Eds.; Humana Press: Totowa, NJ, USA, 2009; pp. 175–202. ISBN 978-1-60327-164-6. [Google Scholar]
- Meile, S.; Kilcher, S.; Loessner, M.J.; Dunne, M. Reporter Phage-Based Detection of Bacterial Pathogens: Design Guidelines and Recent Developments. Viruses 2020, 12, 944. [Google Scholar] [CrossRef]
- Jurač, K.; Nabergoj, D.; Podgornik, A. Bacteriophage Production Processes. Appl. Microbiol. Biotechnol. 2019, 103, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Ames, G.F.; Prody, C.; Kustu, S. Simple, Rapid, and Quantitative Release of Periplasmic Proteins by Chloroform. J. Bacteriol. 1984, 160, 1181–1183. [Google Scholar] [CrossRef] [PubMed]
- Schofield, D.; Sharp, N.J.; Westwater, C. Phage-Based Platforms for the Clinical Detection of Human Bacterial Pathogens. Bacteriophage 2012, 2, 105–121. [Google Scholar] [CrossRef]
- Triffault-Fillit, C.; Ferry, T.; Laurent, F.; Pradat, P.; Dupieux, C.; Conrad, A.; Becker, A.; Lustig, S.; Fessy, M.H.; Chidiac, C.; et al. Microbiologic Epidemiology Depending on Time to Occurrence of Prosthetic Joint Infection: A Prospective Cohort Study. Clin. Microbiol. Infect. 2019, 25, 353–358. [Google Scholar] [CrossRef]
- Sebastian, S.; Malhotra, R.; Sreenivas, V.; Kapil, A.; Chaudhry, R.; Dhawan, B. Sonication of Orthopaedic Implants: A Valuable Technique for Diagnosis of Prosthetic Joint Infections. J. Microbiol. Methods 2018, 146, 51–54. [Google Scholar] [CrossRef]
- Erivan, R.; Villatte, G.; Eymond, G.; Mulliez, A.; Descamps, S.; Boisgard, S. Usefulness of Sonication for Diagnosing Infection in Explanted Orthopaedic Implants. Orthop. Traumatol. Surg. Res. 2018, 104, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Ueda, N.; Oe, K.; Nakamura, T.; Tsuta, K.; Iida, H.; Saito, T. Sonication of Extracted Implants Improves Microbial Detection in Patients with Orthopedic Implant-Associated Infections. J. Arthroplasty 2019, 34, 1189–1196. [Google Scholar] [CrossRef]
- Inacio, R.C.; Klautau, G.B.; Murça, M.A.S.; da Silva, C.B.; Nigro, S.; Rivetti, L.A.; Pereira, W.L.; Salles, M.J.C. Microbial Diagnosis of Infection and Colonization of Cardiac Implantable Electronic Devices by Use of Sonication. Int. J. Infect. Dis. 2015, 38, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Jost, G.F.; Wasner, M.; Taub, E.; Walti, L.; Mariani, L.; Trampuz, A. Sonication of Catheter Tips for Improved Detection of Microorganisms on External Ventricular Drains and Ventriculo-Peritoneal Shunts. J. Clin. Neurosci. 2014, 21, 578–582. [Google Scholar] [CrossRef] [PubMed]
- Fleischmann, S.; Robben, C.; Alter, T.; Rossmanith, P.; Mester, P. How to Evaluate Non-Growing Cells—Current Strategies for Determining Antimicrobial Resistance of VBNC Bacteria. Antibiotics 2021, 10, 115. [Google Scholar] [CrossRef] [PubMed]
- Cerca, F.; Trigo, G.; Correia, A.; Cerca, N.; Azeredo, J.; Vilanova, M. SYBR Green as a Fluorescent Probe to Evaluate the Biofilm Physiological State of Staphylococcus epidermidis, Using Flow Cytometry. Can. J. Microbiol. 2011, 57, 850–856. [Google Scholar] [CrossRef] [PubMed]
- Ou, A.; Wang, K.; Mao, Y.; Yuan, L.; Ye, Y.; Chen, L.; Zou, Y.; Huang, T. First Report on the Rapid Detection and Identification of Methicillin-Resistant Staphylococcus Aureus (MRSA) in Viable but Non-Culturable (VBNC) Under Food Storage Conditions. Front. Microbiol. 2021, 11, 615875. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, T.-Y.; Mao, Y.; Chen, Y.; Shi, F.; Peng, R.; Chen, J.; Yuan, L.; Bai, C.; Chen, L.; et al. Study on the Viable but Non-Culturable (VBNC) State Formation of Staphylococcus Aureus and Its Control in Food System. Front. Microbiol. 2020, 11, 599739. [Google Scholar] [CrossRef] [PubMed]
- Awais, R.; Fukudomi, H.; Miyanaga, K.; Unno, H.; Tanji, Y. A Recombinant Bacteriophage-Based Assay for the Discriminative Detection of Culturable and Viable but Nonculturable Escherichia Coli O157:H7. Biotechnol. Prog. 2006, 22, 853–859. [Google Scholar] [CrossRef]
- Fernandes, E.; Martins, V.C.; Nóbrega, C.; Carvalho, C.M.; Cardoso, F.A.; Cardoso, S.; Dias, J.; Deng, D.; Kluskens, L.D.; Freitas, P.P.; et al. A Bacteriophage Detection Tool for Viability Assessment of Salmonella Cells. Biosens. Bioelectron. 2014, 52, 239–246. [Google Scholar] [CrossRef]
- Zimmerli, W.; Trampuz, A.; Ochsner, P.E. Prosthetic-Joint Infections. N. Engl. J. Med. 2004, 351, 1645–1654. [Google Scholar] [CrossRef]
- Peel, T.N.; Buising, K.L.; Choong, P.F.M. Prosthetic Joint Infection: Challenges of Diagnosis and Treatment: Prosthetic Joint Infection. ANZ J. Surg. 2011, 81, 32–39. [Google Scholar] [CrossRef]
- Yan, H.; Li, M.; Meng, L.; Zhao, F. Formation of Viable but Nonculturable State of Staphylococcus Aureus under Frozen Condition and Its Characteristics. Int. J. Food Microbiol. 2021, 357, 109381. [Google Scholar] [CrossRef]
- Robben, C.; Fister, S.; Witte, A.K.; Schoder, D.; Rossmanith, P.; Mester, P. Induction of the Viable but Non-Culturable State in Bacterial Pathogens by Household Cleaners and Inorganic Salts. Sci. Rep. 2018, 8, 15132. [Google Scholar] [CrossRef] [PubMed]
- Cerca, F.; Andrade, F.; Franca, A.; Andrade, E.B.; Ribeiro, A.; Almeida, A.A.; Cerca, N.; Pier, G.; Azeredo, J.; Vilanova, M. Staphylococcus Epidermidis Biofilms with Higher Proportions of Dormant Bacteria Induce a Lower Activation of Murine Macrophages. J. Med. Microbiol. 2011, 60, 1717–1724. [Google Scholar] [CrossRef] [Green Version]
- Zandri, G.; Pasquaroli, S.; Vignaroli, C.; Talevi, S.; Manso, E.; Donelli, G.; Biavasco, F. Detection of Viable but Non-Culturable Staphylococci in Biofilms from Central Venous Catheters Negative on Standard Microbiological Assays. Clin. Microbiol. Infect. 2012, 18, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Wilks, S.A.; Koerfer, V.V.; Prieto, J.A.; Fader, M.; Keevil, C.W. Biofilm Development on Urinary Catheters Promotes the Appearance of Viable but Nonculturable Bacteria. Mbio 2021, 12, e03584-20. [Google Scholar] [CrossRef] [PubMed]
- França, A.; Gaio, V.; Lopes, N.; Melo, L.D.R. Virulence Factors in Coagulase-Negative Staphylococci. Pathogens 2021, 10, 170. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and Broth Dilution Methods to Determine the Minimal Inhibitory Concentration (MIC) of Antimicrobial Substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Merritt, J.H.; Kadouri, D.E.; O’Toole, G.A. Growing and Analyzing Static Biofilms. In Current Protocols in Microbiology; Coico, R., Kowalik, T., Quarles, J., Stevenson, B., Taylor, R., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2005; ISBN 978-0-471-72925-9. [Google Scholar]
- De Kievit, T. 1.39—Biofilms. In Comprehensive Biotechnology, 3rd ed.; Moo-Young, M., Ed.; Pergamon: Oxford, UK, 2011; pp. 529–540. ISBN 978-0-444-64047-5. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Bacterial Strain | Conditions for VBNC Induction | Days under Stress Conditions Until Loss of Culturability (Mean ± SE) 1 | % of Viable Bacterial Cells after Culturability Loss (Mean ± SE) 1 |
|---|---|---|---|
| S. aureus ATCC 25923 | S | >30 | - |
| S + G 4 × MIC | 11.3 ± 1.3 | 11.9 ± 1.5 | |
| S + G 8 × MIC | 8.0 ± 1.0 | 13.5 ± 0.4 | |
| S + G 16 × MIC | 9.0 ± 1.0 | 14.0 ± 0.7 | |
| S. aureus OBV 17/166 | S | >30 | - |
| S + G 4 × MIC | 19.3 ± 1.4 | 9.7 ± 0.1 | |
| S + G 8 × MIC | 19.3 ± 1.4 | 14.2 ± 0.0 | |
| S + G 16 × MIC | 13.7 ± 3.3 | 10.3 ± 0.9 | |
| S. epidermidis DSM 3269 | S | >30 | - |
| S + G 4 × MIC | 16.0 ± 0.0 | 6.1 ± 0.4 | |
| S + G 8 × MIC | 12.7 ± 1.3 | 10.7 ± 1.6 | |
| S + G 16 × MIC | 8.0 ± 1.0 | 12.0 ± 0.7 | |
| S. lugdunensis OBV 20/143 | S | >30 | - |
| S + G 4 × MIC | 15.3 ± 0.7 | 15.2 ± 3.5 | |
| S + G 8 × MIC | 14.0 ± 0.0 | 23.7 ± 0.5 | |
| S + G 16 × MIC | 10.0 ± 0.0 | 23.9 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Šuster, K.; Cör, A. Induction of Viable but Non-Culturable State in Clinically Relevant Staphylococci and Their Detection with Bacteriophage K. Antibiotics 2023, 12, 311. https://doi.org/10.3390/antibiotics12020311
Šuster K, Cör A. Induction of Viable but Non-Culturable State in Clinically Relevant Staphylococci and Their Detection with Bacteriophage K. Antibiotics. 2023; 12(2):311. https://doi.org/10.3390/antibiotics12020311
Chicago/Turabian StyleŠuster, Katja, and Andrej Cör. 2023. "Induction of Viable but Non-Culturable State in Clinically Relevant Staphylococci and Their Detection with Bacteriophage K" Antibiotics 12, no. 2: 311. https://doi.org/10.3390/antibiotics12020311
APA StyleŠuster, K., & Cör, A. (2023). Induction of Viable but Non-Culturable State in Clinically Relevant Staphylococci and Their Detection with Bacteriophage K. Antibiotics, 12(2), 311. https://doi.org/10.3390/antibiotics12020311