


Clinical Impact of Staphylococcus aureus Skin and Soft Tissue Infections


Abstract
:1. Epidemiology
2. Morbidity and Mortality
3. Economic Burden
4. Clinical Presentations
4.1. Folliculitis
4.2. Boils/Furunculosis
4.3. Carbuncles
4.4. Impetigo
4.5. Erysipelas/Cellulitis
4.6. Mastitis
4.7. Folliculitis Decalvans
4.8. Staphylococcal Scalded Skin Syndrome
4.9. Necrotizing Fasciitis
5. Pathogenesis
6. Immune Response
7. Treatment
7.1. Selection of Antimicrobial Therapeutic Agents
7.2. Antimicrobial Therapeutic Agents Options
Overview
7.3. Commonly Utilized Therapeutic Agents
7.3.1. Sulfamethoxazole-Trimethoprim (TMP/SMX)
7.3.2. Clindamycin
7.3.3. Doxycycline/Minocycline
7.3.4. Linezolid/Tedizolid
7.3.5. Vancomycin
7.3.6. Daptomycin
7.3.7. First Generation Cephalosporins/Penicillinase-Resistant Penicillins
7.4. Infrequently Utilized Therapeutic Agents
7.4.1. Ceftaroline
7.4.2. Dalbavancin/Oritavancin
7.4.3. Tigecycline/Omadacycline
7.4.4. Delafloxacin
7.4.5. Rifampin
7.5. Route of Treatment Administration and Duration
7.6. Phage Therapy
7.7. Treatment Challenges and Considerations
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wertheim, H.F.; Melles, D.C.; Vos, M.C.; van Leeuwen, W.; van Belkum, A.; Verbrugh, H.A.; Nouwen, J.L. The role of nasal carriage in Staphylococcus aureus infections. Lancet Infect. Dis. 2005, 5, 751–762. [Google Scholar] [CrossRef]
- Grundmann, H.; Aires-De-Sousa, M.; Boyce, J.; Tiemersma, E. Emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet 2006, 368, 874–885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenover, F.C.; McDougal, L.K.; Goering, R.V.; Killgore, G.; Projan, S.J.; Patel, J.B.; Dunman, P.M. Characterization of a Strain of Community-Associated Methicillin-Resistant Staphylococcus aureus Widely Disseminated in the United States. J. Clin. Microbiol. 2006, 44, 108–118. [Google Scholar] [CrossRef] [Green Version]
- De Leo, F.R.; Kennedy, A.D.; Chen, L.; Wardenburg, J.B.; Kobayashi, S.D.; Mathema, B.; Braughton, K.R.; Whitney, A.R.; Villaruz, A.E.; Martens, C.A.; et al. Molecular differentiation of historic phage-type 80/81 and contemporary epidemic Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2011, 108, 18091–18096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G., Jr. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Ray, G.T.; Suaya, J.A.; Baxter, R. Incidence, microbiology, and patient characteristics of skin and soft-tissue infections in a U.S. population: A retrospective population-based study. BMC Infect. Dis. 2013, 13, 252. [Google Scholar] [CrossRef] [Green Version]
- Pallin, D.J.; Egan, D.J.; Pelletier, A.J.; Espinola, J.A.; Hooper, D.C.; Camargo, C.A., Jr. Increased US Emergency Department Visits for Skin and Soft Tissue Infections, and Changes in Antibiotic Choices, During the Emergence of Community-Associated Methicillin-Resistant Staphylococcus aureus. Ann. Emerg. Med. 2008, 51, 291–298. [Google Scholar] [CrossRef]
- Hersh, A.L.; Chambers, H.F.; Maselli, J.H.; Gonzales, R. National Trends in Ambulatory Visits and Antibiotic Prescribing for Skin and Soft-Tissue Infections. Arch. Intern. Med. 2008, 168, 1585–1591. [Google Scholar] [CrossRef]
- Edelsberg, J.; Taneja, C.; Zervos, M.; Haque, N.; Moore, C.; Reyes, K.; Al, J.E.E.; Jiang, J.; Oster, G. Trends in US Hospital Admissions for Skin and Soft Tissue Infections. Emerg. Infect. Dis. 2009, 15, 1516–1518. [Google Scholar] [CrossRef]
- Frei, C.R.; Makos, B.R.; Daniels, K.R.; Oramasionwu, C.U. Emergence of community-acquired methicillin-resistant Staphylococcus aureus skin and soft tissue infections as a common cause of hospitalization in United States children. J. Pediatr. Surg. 2010, 45, 1967–1974. [Google Scholar] [CrossRef]
- Miller, L.G.; Eisenberg, D.F.; Liu, H.; Chang, C.-L.; Wang, Y.; Luthra, R.; Wallace, A.; Fang, C.; Singer, J.; Suaya, J.A. Incidence of skin and soft tissue infections in ambulatory and inpatient settings, 2005–2010. BMC Infect. Dis. 2015, 15, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, H.F.; De Leo, F.R. Waves of resistance: Staphylococcus aureus in the antibiotic era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Klein, E.Y.; Mojica, N.; Jiang, W.; Cosgrove, S.E.; Septimus, E.; Morgan, D.J.; Laxminarayan, R. Trends in Methicillin-Resistant Staphylococcus aureus Hospitalizations in the United States, 2010–2014. Clin. Infect. Dis. 2017, 65, 1921–1923. [Google Scholar] [CrossRef] [Green Version]
- Klein, E.Y.; Zhu, X.; Petersen, M.; Patel, E.U.; Cosgrove, S.E.; Tobian, A.A.R. Methicillin-Resistant and Methicillin-Sensitive Staphylococcus aureus Hospitalizations: National Inpatient Sample, 2016–2019. Open Forum Infect. Dis. 2022, 9, ofab585. [Google Scholar] [CrossRef]
- King, M.D.; Humphrey, B.B.J.; Wang, Y.F.; Kourbatova, E.V.; Ray, S.M.; Blumberg, H.M. Emergence of Community-Acquired Methicillin-Resistant Staphylococcus aureus USA 300 Clone as the Predominant Cause of Skin and Soft-Tissue Infections. Ann. Intern. Med. 2006, 144, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, G.J.; Krishnadasan, A.; Gorwitz, R.J.; Fosheim, G.E.; McDougal, L.K.; Carey, R.B.; Talan, D.A. Methicillin-Resistant S. aureus Infections among Patients in the Emergency Department. N. Engl. J. Med. 2006, 355, 666–674. [Google Scholar] [CrossRef]
- Talan, D.A.; Krishnadasan, A.; Gorwitz, R.J.; Fosheim, G.E.; Limbago, B.; Albrecht, V.; Moran, G.J. Comparison of Staphylococcus aureus From Skin and Soft-Tissue Infections in US Emergency Department Patients, 2004 and 2008. Clin. Infect. Dis. 2011, 53, 144–149. [Google Scholar] [CrossRef]
- Nichol, K.A.; Adam, H.J.; Hussain, Z.; Mulvey, M.R.; McCracken, M.; Mataseje, L.F.; Thompson, K.; Kost, S.; Lagacé-Wiens, P.R.; Hoban, D.J.; et al. Comparison of community-associated and health care-associated methicillin-resistant Staphylococcus aureus in Canada: Results of the CANWARD 2007–2009 study. Diagn. Microbiol. Infect. Dis. 2011, 69, 320–325. [Google Scholar] [CrossRef]
- Nimmo, G.R. USA300 abroad: Global spread of a virulent strain of community-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 725–734. [Google Scholar] [CrossRef] [Green Version]
- Otter, J.A.; Havill, N.L.; Boyce, J.M.; French, G.L. Comparison of community-associated meticillin-resistant Staphylococcus aureus from teaching hospitals in London and the USA, 2004–2006: Where is USA300 in the UK? Eur. J. Clin. Microbiol. Infect. Dis. 2009, 28, 835–839. [Google Scholar] [CrossRef]
- Enström, J.; Fröding, I.; Giske, C.G.; Ininbergs, K.; Bai, X.; Sandh, G.; Tollström, U.-B.; Ullberg, M.; Fang, H. USA300 methicillin-resistant Staphylococcus aureus in Stockholm, Sweden, from 2008 to 2016. PLoS ONE 2018, 13, e0205761. [Google Scholar] [CrossRef] [PubMed]
- Takadama, S.; Nakaminami, H.; Sato, A.; Shoshi, M.; Fujii, T.; Noguchi, N. Dissemination of Panton-Valentine leukocidin–positive methicillin-resistant Staphylococcus aureus USA300 clone in multiple hospitals in Tokyo, Japan. Clin. Microbiol. Infect. 2018, 24, 1211.e1–1211.e7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-C.; Chen, C.-J.; Kuo, C.-C.; Lu, M.-C. Emergence, transmission and phylogeny of meticillin-resistant Staphylococcus aureus sequence type 8 (USA300) in Taiwan. J. Hosp. Infect. 2018, 100, 355–358. [Google Scholar] [CrossRef]
- Nimmo, G.R.; Coombs, G.W.; Pearson, J.C.; O’brien, F.G.; Christiansen, K.J.; Turnidge, J.D.; Gosbell, I.B.; Collignon, P.; McLaws, M.-L. Methicillin-resistant Staphylococcus aureus in the Australian community: An evolving epidemic. Med. J. Aust. 2006, 184, 384–388. [Google Scholar] [CrossRef] [Green Version]
- Vella, V.; Galgani, I.; Polito, L.; Arora, A.K.; Creech, C.B.; David, M.Z.; Lowy, F.D.; Macesic, N.; Ridgway, J.P.; Uhlemann, A.-C.; et al. Staphylococcus aureus Skin and Soft Tissue Infection Recurrence Rates in Outpatients: A Retrospective Database Study at 3 US Medical Centers. Clin. Infect. Dis. 2021, 73, e1045–e1053. [Google Scholar] [CrossRef]
- Russo, A.; Concia, E.; Cristini, F.; De Rosa, F.G.; Esposito, S.; Menichetti, F.; Petrosillo, N.; Tumbarello, M.; Venditti, M.; Viale, P.; et al. Current and future trends in antibiotic therapy of acute bacterial skin and skin-structure infections. Clin. Microbiol. Infect. 2016, 22 (Suppl. 2), S27–S36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenstrom, R.; Grafstein, E.; Romney, M.; Fahimi, J.; Harris, D.; Hunte, G.; Innes, G.; Christenson, J. Prevalence of and risk factors for methicillin-resistant Staphylococcus aureus skin and soft tissue infection in a Canadian emergency department. Can. J. Emerg. Med. 2009, 11, 430–438. [Google Scholar] [CrossRef] [Green Version]
- Zilberberg, M.D.; Chaudhari, P.; Nathanson, B.H.; Campbell, R.S.; Emons, M.F.; Fiske, S.; Hays, H.D.; Shorr, A.F. Development and validation of a bedside risk score for MRSA among patients hospitalized with complicated skin and skin structure infections. BMC Infect. Dis. 2012, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Olaniyi, R.; Pozzi, C.; Grimaldi, L.; Bagnoli, F. Staphylococcus aureus -Associated Skin and Soft Tissue Infections: Anatomical Localization, Epidemiology, Therapy and Potential Prophylaxis. Curr. Top. Microbiol. Immunol. 2017, 409, 199–227. [Google Scholar] [CrossRef]
- Kazakova, S.V.; Hageman, J.C.; Matava, M.; Srinivasan, A.; Phelan, L.; Garfinkel, B.; Boo, T.; McAllister, S.; Anderson, J.; Jensen, B.; et al. A Clone of Methicillin-Resistant Staphylococcus aureus among Professional Football Players. N. Engl. J. Med. 2005, 352, 468–475. [Google Scholar] [CrossRef]
- Haysom, L.; Cross, M.; Anastasas, R.; Moore, E.; Hampton, S. Prevalence and Risk Factors for Methicillin-Resistant Staphylococcus aureus (MRSA) Infections in Custodial Populations: A Systematic Review. J. Correct. Health Care 2018, 24, 197–213. [Google Scholar] [CrossRef] [Green Version]
- Fritz, S.A.; Epplin, E.K.; Garbutt, J.; Storch, G.A. Skin infection in children colonized with community-associated methicillin-resistant Staphylococcus aureus. J. Infect. 2009, 59, 394–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Creech, C.B.; Al-Zubeidi, D.N.; Fritz, S.A. Prevention of Recurrent Staphylococcal Skin Infections. Infect. Dis. Clin. N. Am. 2015, 29, 429–464. [Google Scholar] [CrossRef] [Green Version]
- Prevention, C.f.D.C.a. Healthcare-Associated Infections (HAIs): Staphylococcus aureus. Available online: https://www.cdc.gov/hai/organisms/staph.html (accessed on 5 November 2022).
- Ikuta, K.S.; Swetschinski, L.R.; Aguilar, G.R.; Sharara, F.; Mestrovic, T.; Gray, A.P.; Weaver, N.D.; Wool, E.; Han, C.; Hayoon, A.G.; et al. Global mortality associated with 33 bacterial pathogens in 2019: A systematic analysis for the Global Burden of Disease Study 2019. Lancet 2022, 400, 2221–2248. [Google Scholar] [CrossRef] [PubMed]
- Mestrovic, T.; Aguilar, G.R.; Swetschinski, L.R.; Ikuta, K.S.; Gray, A.P.; Weaver, N.D.; Han, C.; Wool, E.; Hayoon, A.G.; Hay, S.; et al. The burden of bacterial antimicrobial resistance in the WHO European region in 2019: A cross-country systematic analysis. Lancet Public Health 2022, 7, e897–e913. [Google Scholar] [CrossRef]
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global burden of bacterial antimicrobial resistance in 2019: A systematic analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- Tacconelli, E.; Carrara, E.; Savoldi, A.; Harbarth, S.; Mendelson, M.; Monnet, D.L.; Pulcini, C.; Kahlmeter, G.; Kluytmans, J.; Carmeli, Y.; et al. Discovery, research, and development of new antibiotics: The WHO priority list of antibiotic-resistant bacteria and tuberculosis. Lancet Infect. Dis. 2018, 18, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Cranendonk, D.R.; Van Vught, L.A.; Wiewel, M.A.; Cremer, O.L.; Horn, J.; Bonten, M.J.; Schultz, M.J.; Van Der Poll, T.; Wiersinga, W.J. Clinical Characteristics and Outcomes of Patients with Cellulitis Requiring Intensive Care. JAMA Dermatol. 2017, 153, 578–582. [Google Scholar] [CrossRef]
- Klevens, R.M.; Morrison, M.A.; Nadle, J.; Petit, S.; Gershman, K.; Ray, S.; Harrison, L.H.; Lynfield, R.; Dumyati, G.; Townes, J.M.; et al. Invasive Methicillin-Resistant Staphylococcus aureus Infections in the United States. JAMA 2007, 298, 1763–1771. [Google Scholar] [CrossRef] [Green Version]
- Furst, M.J.L.; de Vedia, L.; Fernández, S.; Gardella, N.; Ganaha, M.C.; Prieto, S.; Carbone, E.; Lista, N.; Rotryng, F.; Morera, G.I.; et al. Prospective Multicenter Study of Community-Associated Skin and Skin Structure Infections due to Methicillin-Resistant Staphylococcus aureus in Buenos Aires, Argentina. PLoS ONE 2013, 8, e78303. [Google Scholar] [CrossRef] [Green Version]
- Macmorran, E.; Harch, S.; Athan, E.; Lane, S.; Tong, S.; Crawford, L.; Krishnaswamy, S.; Hewagama, S. The rise of methicillin resistant Staphylococcus aureus: Now the dominant cause of skin and soft tissue infection in Central Australia. Epidemiol. Infect. 2017, 145, 2817–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.Y.; Singh, A.; David, M.Z.; Bartsch, S.M.; Slayton, R.B.; Huang, S.S.; Zimmer, S.M.; Potter, M.A.; Macal, C.M.; Lauderdale, D.S.; et al. The economic burden of community-associated methicillin-resistant Staphylococcus aureus (CA-MRSA). Clin. Microbiol. Infect. 2013, 19, 528–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun, K.; Shurko, J.F.; Ryan, L.; Lee, G.C. Age-based health and economic burden of skin and soft tissue infections in the United States, 2000 and 2012. PLoS ONE 2018, 13, e0206893. [Google Scholar] [CrossRef] [PubMed]
- Marton, J.P.; Jackel, J.L.; Carson, R.T.; Rothermel, C.D.; Friedman, M.; Menzin, J. Costs of skin and skin structure infections due to Staphylococcus aureus: An analysis of managed- care claims. Curr. Med. Res. Opin. 2008, 24, 2821–2828. [Google Scholar] [CrossRef]
- Suaya, J.A.; Mera, R.M.; Cassidy, A.; O’Hara, P.; Amrine-Madsen, H.; Burstin, S.; Miller, L.G. Incidence and cost of hospitalizations associated with Staphylococcus aureus skin and soft tissue infections in the United States from 2001 through 2009. BMC Infect. Dis. 2014, 14, 296. [Google Scholar] [CrossRef] [Green Version]
- Itani, K.M.F.; Merchant, S.; Lin, S.-J.; Akhras, K.; Alandete, J.C.; Hatoum, H.T. Outcomes and management costs in patients hospitalized for skin and skin-structure infections. Am. J. Infect. Control. 2011, 39, 42–49. [Google Scholar] [CrossRef]
- Frazee, B.W.; Lynn, J.; Charlebois, E.D.; Lambert, L.; Lowery, D.; Perdreau-Remington, F. High Prevalence of Methicillin-Resistant Staphylococcus aureus in Emergency Department Skin and Soft Tissue Infections. Ann. Emerg. Med. 2005, 45, 311–320. [Google Scholar] [CrossRef]
- LaBreche, M.J.; Lee, G.C.; Attridge, R.T.; Mortensen, E.M.; Koeller, J.; Du, L.C.; Nyren, N.R.; Treviño, L.B.; Treviño, S.B.; Peña, J.; et al. Treatment Failure and Costs in Patients with Methicillin-Resistant Staphylococcus aureus (MRSA) Skin and Soft Tissue Infections: A South Texas Ambulatory Research Network (STARNet) Study. J. Am. Board Fam. Med. 2013, 26, 508–517. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.E.; Samore, M.H.; Jones, M.; Greene, T.; Stevens, V.W.; Liu, C.-F.; Graves, N.; Evans, M.F.; Rubin, M.A. Reducing Time-dependent Bias in Estimates of the Attributable Cost of Health Care–associated Methicillin-resistant Staphylococcus aureus Infections: A Comparison of Three Estimation Strategies. Med. Care 2015, 53, 827–834. [Google Scholar] [CrossRef]
- Branch-Elliman, W.; Lee, G.M.; Golen, T.H.; Gold, H.S.; Baldini, L.M.; Wright, S.B. Health and Economic Burden of Post-Partum Staphylococcus aureus Breast Abscess. PLoS ONE 2013, 8, e73155. [Google Scholar] [CrossRef] [Green Version]
- Zhen, X.; Lundborg, C.S.; Sun, X.; Hu, X.; Dong, H. Economic burden of antibiotic resistance in ESKAPE organisms: A systematic review. Antimicrob. Resist. Infect. Control. 2019, 8, 137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laureano, A.C.; Schwartz, R.A.; Cohen, P.J. Facial bacterial infections: Folliculitis. Clin. Dermatol. 2014, 32, 711–714. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.; Halmi, B.H. Bockhart’s impetigo: Complication of waterbed use. Int. J. Dermatol. 1999, 38, 769–770. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, P. Skin Infections Caused by Staphylococcus aureus. Acta Derm. Venereol. 2020, 100, adv00110. [Google Scholar] [CrossRef]
- Breyre, A.; Frazee, B.W. Skin and Soft Tissue Infections in the Emergency Department. Emerg. Med. Clin. N. Am. 2018, 36, 723–750. [Google Scholar] [CrossRef] [PubMed]
- Marques, S.A.; Abbade, L.P.F. Severe bacterial skin infections. An. Bras. Dermatol. 2020, 95, 407–417. [Google Scholar] [CrossRef]
- Durupt, F.; Mayor, L.; Bes, M.; Reverdy, M.-E.; Vandenesch, F.; Thomas, L.; Etienne, J. Prevalence of Staphylococcus aureus toxins and nasal carriage in furuncles and impetigo. Br. J. Dermatol. 2007, 157, 1161–1167. [Google Scholar] [CrossRef]
- Yamasaki, O.; Kaneko, J.; Morizane, S.; Akiyama, H.; Arata, J.; Narita, S.; Chiba, J.; Kamio, Y.; Iwatsuki, K. The Association between Staphylococcus aureus Strains Carrying Panton-Valentine Leukocidin Genes and the Development of Deep-Seated Follicular Infection. Clin. Infect. Dis. 2005, 40, 381–385. [Google Scholar] [CrossRef] [Green Version]
- Masiuk, H.; Kopron, K.; Grumann, D.; Goerke, C.; Kolata, J.; Jursa-Kulesza, J.; Giedrys-Kalemba, S.; Bröker, B.M.; Holtfreter, S. Association of Recurrent Furunculosis with Panton-Valentine Leukocidin and the Genetic Background of Staphylococcus aureus. J. Clin. Microbiol. 2010, 48, 1527–1535. [Google Scholar] [CrossRef] [Green Version]
- Baggett, H.C.; Hennessy, T.W.; Rudolph, K.; Bruden, D.; Reasonover, A.; Parkinson, A.; Sparks, R.; Donlan, R.M.; Martinez, P.; Mongkolrattanothai, K.; et al. Community-Onset Methicillin-Resistant Staphylococcus aureus Associated with Antibiotic Use and the Cytotoxin Panton-Valentine Leukocidin during a Furunculosis Outbreak in Rural Alaska. J. Infect. Dis. 2004, 189, 1565–1573. [Google Scholar] [CrossRef] [Green Version]
- Eley, C.D.; Gan, V.N. Picture of the Month: Folliculitis, furunculosis, and carbuncles. Arch. Pediatr. Adolesc. Med. 1997, 151, 625–626. [Google Scholar] [CrossRef]
- Mine, Y.; Higuchi, W.; Taira, K.; Nakasone, I.; Tateyama, M.; Yamamoto, T.; Uezato, H.; Takahashi, K. Nosocomial outbreak of multidrug-resistant USA300 methicillin-resistant Staphylococcus aureus causing severe furuncles and carbuncles in Japan. J. Dermatol. 2011, 38, 1167–1171. [Google Scholar] [CrossRef] [PubMed]
- Torok, M.E.; Conlon, C.P. Skin and soft tissue infections. Medicine 2009, 37, 603–609. [Google Scholar] [CrossRef]
- Rodriguez-Iturbe, B.; Haas, M. Post-Streptococcus pyogenes Glomerulonephritis. In Streptococcus Pyogenes: Basic Biology to Clinical Manifestations; Ferretti, J.J., Stevens, D.L., Fischetti, V.A., Eds.; The University of Oklahoma Health Sciences Center: Oklahoma City, OK, USA, 2022. [Google Scholar]
- Gunderson, C.G.; Martinello, R.A. A systematic review of bacteremias in cellulitis and erysipelas. J. Infect. 2012, 64, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Bläckberg, A.; Trell, K.; Rasmussen, M. Erysipelas, a large retrospective study of aetiology and clinical presentation. BMC Infect. Dis. 2015, 15, 402. [Google Scholar] [CrossRef] [Green Version]
- Bruun, T.; Oppegaard, O.; Kittang, B.R.; Mylvaganam, H.; Langeland, N.; Skrede, S. Etiology of Cellulitis and Clinical Prediction of Streptococcal Disease: A Prospective Study. Open Forum Infect. Dis. 2016, 3, ofv181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.P. Management of mastitis in breastfeeding women. Am. Fam. Physician 2008, 78, 727–731. [Google Scholar]
- Delgado, S.; García, P.; Fernández, L.; Jiménez, E.; Rodríguez-Baños, M.; del Campo, R.; Rodríguez, J.M. Characterization of Staphylococcus aureus strains involved in human and bovine mastitis. FEMS Immunol. Med. Microbiol. 2011, 62, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Angelopoulou, A.; Field, D.; Ryan, C.A.; Stanton, C.; Hill, C.; Ross, R.P. The microbiology and treatment of human mastitis. Med. Microbiol. Immunol. 2018, 207, 83–94. [Google Scholar] [CrossRef]
- Rimoldi, S.G.; Pileri, P.; Mazzocco, M.I.; Romeri, F.; Bestetti, G.; Calvagna, N.; Tonielli, C.; Fiori, L.; Gigantiello, A.; Pagani, C.; et al. The Role of Staphylococcus aureus in Mastitis: A Multidisciplinary Working Group Experience. J. Hum. Lact. 2020, 36, 503–509. [Google Scholar] [CrossRef]
- Asfour, L.; Trautt, E.; Harries, M.J. Folliculitis decalvans in the era of antibiotic resistance: Microbiology and antibiotic sensitivities in a tertiary hair clinic. Int. J. Trichology 2020, 12, 193–194. [Google Scholar] [CrossRef]
- Matard, B.; Donay, J.L.; Resche-Rigon, M.; Tristan, A.; Farhi, D.; Rousseau, C.; Mercier-Delarue, S.; Cavelier–Balloy, B.; Assouly, P.; Petit, A.; et al. Folliculitis decalvans is characterized by a persistent, abnormal subepidermal microbiota. Exp. Dermatol. 2020, 29, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Ladhani, S.; Joannou, C.L.; Lochrie, D.P.; Evans, R.W.; Poston, S.M. Clinical, Microbial, and Biochemical Aspects of the Exfoliative Toxins Causing Staphylococcal Scalded-Skin Syndrome. Clin. Microbiol. Rev. 1999, 12, 224–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamand, V.; Dauwalder, O.; Tristan, A.; Casalegno, J.S.; Meugnier, H.; Bes, M.; Dumitrescu, O.; Croze, M.; Vandenesch, F.; Etienne, J.; et al. Epidemiological data of staphylococcal scalded skin syndrome in France from 1997 to 2007 and microbiological characteristics of Staphylococcus aureus associated strains. Clin. Microbiol. Infect. 2012, 18, E514–E521. [Google Scholar] [CrossRef] [Green Version]
- Yamasaki, O.; Yamaguchi, T.; Sugai, M.; Chapuis-Cellier, C.; Arnaud, F.; Vandenesch, F.; Etienne, J.; Lina, G. Clinical Manifestations of Staphylococcal Scalded-Skin Syndrome Depend on Serotypes of Exfoliative Toxins. J. Clin. Microbiol. 2005, 43, 1890–1893. [Google Scholar] [CrossRef] [Green Version]
- Haggerty, J.; Grimaldo, F. A Desquamating Skin Rash in a Pediatric Patient. Clin. Pract. Cases Emerg. Med. 2019, 3, 112–114. [Google Scholar] [CrossRef]
- Jordan, K.S. Staphylococcal Scalded Skin Syndrome. Adv. Emerg. Nurs. J. 2019, 41, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.G.; Perdreau-Remington, F.; Rieg, G.; Mehdi, S.; Perlroth, J.; Bayer, A.S.; Tang, A.W.; Phung, T.O.; Spellberg, B. Necrotizing Fasciitis Caused by Community-Associated Methicillin-Resistant Staphylococcus aureus in Los Angeles. N. Engl. J. Med. 2005, 352, 1445–1453. [Google Scholar] [CrossRef] [Green Version]
- Venbrocks, R.; Lange, M.; Roth, A.; Mollenhauer, J.; Straube, E. Overwhelming septic infection with a multi-resistant Staphylococcus aureus (MRSA) after total knee replacement. Arch. Orthop. Trauma Surg. 2003, 123, 429–432. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Park, S.Y.; Kwak, Y.G.; Jung, J.; Kim, M.-C.; Choi, S.-H.; Yu, S.N.; Hong, H.-L.; Kim, Y.K.; Park, S.Y.; et al. Etiology, characteristics, and outcomes of community-onset necrotizing fasciitis in Korea: A multicenter study. PLoS ONE 2019, 14, e0218668. [Google Scholar] [CrossRef] [Green Version]
- Puvanendran, R.; Huey, J.C.; Pasupathy, S. Necrotizing fasciitis. Can. Fam. Physician 2009, 55, 981–987. [Google Scholar] [PubMed]
- Thapaliya, D.; O’Brien, A.M.; Wardyn, S.E.; Smith, T.C. Epidemiology of necrotizing infection caused by Staphylococcus aureus and Streptococcus pyogenes at an Iowa hospital. J. Infect. Public Health 2015, 8, 634–641. [Google Scholar] [CrossRef] [Green Version]
- Cheng, N.-C.; Wang, J.-T.; Chang, S.-C.; Tai, H.-C.; Tang, Y.-B. Necrotizing Fasciitis Caused by Staphylococcus aureus: The emergence of methicillin-resistant strains. Ann. Plast. Surg. 2011, 67, 632–636. [Google Scholar] [CrossRef] [PubMed]
- Wilke, G.A.; Wardenburg, J.B. Role of a disintegrin and metalloprotease 10 in Staphylococcus aureus α-hemolysin-mediated cellular injury. Proc. Natl. Acad. Sci. USA 2010, 107, 13473–13478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maretzky, T.; Reiss, K.; Ludwig, A.; Buchholz, J.; Scholz, F.; Proksch, E.; de Strooper, B.; Hartmann, D.; Saftig, P. ADAM10 mediates E-cadherin shedding and regulates epithelial cell-cell adhesion, migration, and β-catenin translocation. Proc. Natl. Acad. Sci. USA 2005, 102, 9182–9187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bramley, A.J.; Patel, A.H.; O’Reilly, M.; Foster, R.; Foster, T.J. Roles of alpha-toxin and beta-toxin in virulence of Staphylococcus aureus for the mouse mammary gland. Infect. Immun. 1989, 57, 2489–2494. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, A.D.; Wardenburg, J.B.; Gardner, D.J.; Long, D.; Whitney, A.R.; Braughton, K.R.; Schneewind, O.; DeLeo, F.R. Targeting of Alpha-Hemolysin by Active or Passive Immunization Decreases Severity of USA300 Skin Infection in a Mouse Model. J. Infect. Dis. 2010, 202, 1050–1058. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; Malachowa, N.; DeLeo, F.R. Pathogenesis of Staphylococcus aureus Abscesses. Am. J. Pathol. 2015, 185, 1518–1527. [Google Scholar] [CrossRef] [Green Version]
- Soong, G.; Paulino, F.; Wachtel, S.; Parker, D.; Wickersham, M.; Zhang, D.; Brown, A.; Lauren, C.; Dowd, M.; West, E.; et al. Methicillin-Resistant Staphylococcus aureus Adaptation to Human Keratinocytes. mBio 2015, 6, e00289-15. [Google Scholar] [CrossRef] [Green Version]
- Cruz, A.R.; van Strijp, J.A.G.; Bagnoli, F.; Manetti, A.G.O. Virulence Gene Expression of Staphylococcus aureus in Human Skin. Front. Microbiol. 2021, 12. [Google Scholar] [CrossRef]
- Yanai, M.; Rocha, M.A.; Matolek, A.Z.; Chintalacharuvu, A.; Taira, Y.; Chintalacharuvu, K.; Beenhouwer, D.O. Separately or Combined, LukG/LukH Is Functionally Unique Compared to Other Staphylococcal Bicomponent Leukotoxins. PLoS ONE 2014, 9, e89308. [Google Scholar] [CrossRef] [PubMed]
- Baede, V.O.; Voet, M.M.; van der Reijden, T.J.K.; van Wengen, A.; Horst-Kreft, D.E.; Toom, N.A.L.-D.; Tavakol, M.; Vos, M.C.; Nibbering, P.H.; van Wamel, W.J.B. The survival of epidemic and sporadic MRSA on human skin mimics is determined by both host and bacterial factors. Epidemiol. Infect. 2022, 150, e203. [Google Scholar] [CrossRef] [PubMed]
- Spaan, A.N.; Vrieling, M.; Wallet, P.; Badiou, C.; Reyes-Robles, T.; Ohneck, E.A.; Benito, Y.; de Haas, C.J.C.; Day, C.J.; Jennings, M.P.; et al. The staphylococcal toxins γ-haemolysin AB and CB differentially target phagocytes by employing specific chemokine receptors. Nat. Commun. 2014, 5, 5438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tromp, A.T.; Van Gent, M.; Abrial, P.; Martin, A.; Jansen, J.P.; De Haas, C.J.C.; Van Kessel, K.P.M.; Bardoel, B.W.; Kruse, E.; Bourdonnay, E.; et al. Human CD45 is an F-component-specific receptor for the staphylococcal toxin Panton–Valentine leukocidin. Nat. Microbiol. 2018, 3, 708–717. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcus aureus toxins. Curr. Opin. Microbiol. 2014, 17, 32–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, Y.; Oscherwitz, J.; Cease, K.B.; Chan, S.M.; Muñoz-Planillo, R.; Hasegawa, M.; Villaruz, A.E.; Cheung, G.Y.C.; McGavin, M.J.; Travers, J.B.; et al. Staphylococcus δ-toxin induces allergic skin disease by activating mast cells. Nature 2013, 503, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prince, A.; Wang, H.; Kitur, K.; Parker, D. Humanized Mice Exhibit Increased Susceptibility to Staphylococcus aureus Pneumonia. J. Infect. Dis. 2017, 215, 1386–1395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shallcross, L.J.; Fragaszy, E.; Johnson, A.M.; Hayward, A.C. The role of the Panton-Valentine leucocidin toxin in staphylococcal disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, S.D.; Malachowa, N.; Whitney, A.R.; Braughton, K.R.; Gardner, D.J.; Long, D.; Wardenburg, J.B.; Schneewind, O.; Otto, M.; DeLeo, F.R. Comparative Analysis of USA300 Virulence Determinants in a Rabbit Model of Skin and Soft Tissue Infection. J. Infect. Dis. 2011, 204, 937–941. [Google Scholar] [CrossRef]
- Lipinska, U.; Hermans, K.; Meulemans, L.; Dumitrescu, O.; Badiou, C.; Duchateau, L.; Haesebrouck, F.; Etienne, J.; Lina, G. Panton-Valentine Leukocidin Does Play a Role in the Early Stage of Staphylococcus aureus Skin Infections: A Rabbit Model. PLoS ONE 2011, 6, e22864. [Google Scholar] [CrossRef]
- Malachowa, N.; Kobayashi, S.D.; Braughton, K.R.; Whitney, A.R.; Parnell, M.J.; Gardner, D.J.; DeLeo, F.R. Staphylococcus aureus Leukotoxin GH Promotes Inflammation. J. Infect. Dis. 2012, 206, 1185–1193. [Google Scholar] [CrossRef]
- Chi, C.-Y.; Lin, C.-C.; Liao, I.-C.; Yao, Y.-C.; Shen, F.-C.; Liu, C.-C.; Lin, C.-F. Panton-Valentine Leukocidin Facilitates the Escape of Staphylococcus aureus From Human Keratinocyte Endosomes and Induces Apoptosis. J. Infect. Dis. 2014, 209, 224–235. [Google Scholar] [CrossRef]
- Malachowa, N.; Kobayashi, S.D.; Sturdevant, D.E.; Scott, D.P.; DeLeo, F.R. Insights into the Staphylococcus aureus -Host Interface: Global Changes in Host and Pathogen Gene Expression in a Rabbit Skin Infection Model. PLoS ONE 2015, 10, e0117713. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.W.; Biancotti, J.C.; Berg, B.L.; Gate, D.; Kolar, S.L.; Müller, S.; Rodriguez, M.D.; Rezai-Zadeh, K.; Fan, X.; Beenhouwer, D.O.; et al. Increased Susceptibility of Humanized NSG Mice to Panton-Valentine Leukocidin and Staphylococcus aureus Skin Infection. PLoS Pathog. 2015, 11, e1005292. [Google Scholar] [CrossRef] [PubMed]
- McCormick, J.K.; Yarwood, J.M.; Schlievert, P.M. Toxic Shock Syndrome and Bacterial Superantigens: An Update. Annu. Rev. Microbiol. 2001, 55, 77–104. [Google Scholar] [CrossRef]
- Monday, S.R.; Vath, G.M.; Ferens, W.A.; Deobald, C.; Rago, J.V.; Gahr, P.J.; Monie, D.D.; Iandolo, J.J.; Chapes, S.K.; Davis, W.C.; et al. Unique Superantigen Activity of Staphylococcal Exfoliative Toxins. J. Immunol. 1999, 162, 4550–4559. [Google Scholar] [CrossRef]
- Hanakawa, Y.; Selwood, T.; Woo, D.; Lin, C.; Schechter, N.M.; Stanley, J.R. Calcium-Dependent Conformation of Desmoglein 1 Is Required for its Cleavage by Exfoliative Toxin. J. Investig. Dermatol. 2003, 121, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Amagai, M.; Matsuyoshi, N.; Wang, Z.H.; Andl, C.; Stanley, J.R. Toxin in bullous impetigo and staphylococcal scalded-skin syndrome targets desmoglein 1. Nat. Med. 2000, 6, 1275–1277. [Google Scholar] [CrossRef]
- Amagai, M.; Nishifuji, K.; Yamaguchi, T.; Hanakawa, Y.; Sugai, M.; Stanley, J.R. Staphylococcal Exfoliative Toxin B Specifically Cleaves Desmoglein 1. J. Investig. Dermatol. 2002, 118, 845–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phonimdaeng, P.; O’Reilly, M.; Nowlan, P.; Bramley, A.J.; Foster, T.J. The coagulase of Staphylococcus aureus 8325-4. Sequence analysis and virulence of site-specific coagulase-deficient mutants. Mol. Microbiol. 1990, 4, 393–404. [Google Scholar] [CrossRef]
- Malachowa, N.; Kobayashi, S.D.; Porter, A.R.; Braughton, K.R.; Scott, D.P.; Gardner, D.J.; Missiakas, D.M.; Schneewind, O.; DeLeo, F.R. Contribution of Staphylococcus aureus Coagulases and Clumping Factor A to Abscess Formation in a Rabbit Model of Skin and Soft Tissue Infection. PLoS ONE 2016, 11, e0158293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, T.J.; Geoghegan, J.A.; Ganesh, V.K.; Höök, M. Adhesion, invasion and evasion: The many functions of the surface proteins of Staphylococcus aureus. Nat. Rev. Microbiol. 2014, 12, 49–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwiecinski, J.; Jin, T.; Josefsson, E. Surface proteins of Staphylococcus aureus play an important role in experimental skin infection. Apmis 2014, 122, 1240–1250. [Google Scholar] [CrossRef] [PubMed]
- Cheng, A.G.; Kim, H.K.; Burts, M.L.; Krausz, T.; Schneewind, O.; Missiakas, D.M. Genetic requirements for Staphylococcus aureus abscess formation and persistence in host tissues. FASEB J. 2009, 23, 3393–3404. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; DeDent, A.; Cheng, A.G.; McAdow, M.; Bagnoli, F.; Missiakas, D.M.; Schneewind, O. IsdA and IsdB antibodies protect mice against Staphylococcus aureus abscess formation and lethal challenge. Vaccine 2010, 28, 6382–6392. [Google Scholar] [CrossRef] [Green Version]
- Kuklin, N.A.; Clark, D.J.; Secore, S.; Cook, J.; Cope, L.D.; McNeely, T.; Noble, L.; Brown, M.J.; Zorman, J.K.; Wang, X.M.; et al. A Novel Staphylococcus aureus Vaccine: Iron Surface Determinant B Induces Rapid Antibody Responses in Rhesus Macaques and Specific Increased Survival in a Murine S. aureus Sepsis Model. Infect. Immun. 2006, 74, 2215–2223. [Google Scholar] [CrossRef] [Green Version]
- Proctor, R.A. Challenges for a Universal Staphylococcus aureus Vaccine. Clin. Infect. Dis. 2012, 54, 1179–1186. [Google Scholar] [CrossRef] [Green Version]
- Da Costa, T.M.; Viljoen, A.; Towell, A.M.; Dufrêne, Y.F.; Geoghegan, J.A. Fibronectin binding protein B binds to loricrin and promotes corneocyte adhesion by Staphylococcus aureus. Nat. Commun. 2022, 13, 2517. [Google Scholar] [CrossRef]
- Towell, A.M.; Feuillie, C.; Vitry, P.; Da Costa, T.M.; Mathelié-Guinlet, M.; Kezic, S.; Fleury, O.M.; McAleer, M.A.; Dufrêne, Y.F.; Irvine, A.D.; et al. Staphylococcus aureus binds to the N-terminal region of corneodesmosin to adhere to the stratum corneum in atopic dermatitis. Proc. Natl. Acad. Sci. USA 2020, 118, e2014444118. [Google Scholar] [CrossRef]
- Burian, M.; Plange, J.; Schmitt, L.; Kaschke, A.; Marquardt, Y.; Huth, L.; Baron, J.M.; Hornef, M.W.; Wolz, C.; Yazdi, A.S. Adaptation of Staphylococcus aureus to the Human Skin Environment Identified Using an ex vivo Tissue Model. Front. Microbiol. 2020, 12, 728989. [Google Scholar] [CrossRef]
- Kintarak, S.; Whawell, S.A.; Speight, P.M.; Packer, S.; Nair, S.P. Internalization of Staphylococcus aureus by Human Keratinocytes. Infect. Immun. 2004, 72, 5668–5675. [Google Scholar] [CrossRef] [Green Version]
- Bur, S.; Preissner, K.T.; Herrmann, M.; Bischoff, M. The Staphylococcus aureus Extracellular Adherence Protein Promotes Bacterial Internalization by Keratinocytes Independent of Fibronectin-Binding Proteins. J. Investig. Dermatol. 2013, 133, 2004–2012. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldmann, O.; Tuchscherr, L.; Rohde, M.; Medina, E. α-Hemolysin enhances Staphylococcus aureus internalization and survival within mast cells by modulating the expression of β1 integrin. Cell. Microbiol. 2016, 18, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Hansen, U.; Hussain, M.; Villone, D.; Herrmann, M.; Robenek, H.; Peters, G.; Sinha, B.; Bruckner, P. The anchorless adhesin Eap (extracellular adherence protein) from Staphylococcus aureus selectively recognizes extracellular matrix aggregates but binds promiscuously to monomeric matrix macromolecules. Matrix Biol. 2006, 25, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.; Cockayne, A.; Morrissey, J.A. Iron-Regulated Biofilm Formation in Staphylococcus aureus Newman Requires ica and the Secreted Protein Emp. Infect. Immun. 2008, 76, 1756–1765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geraci, J.; Neubauer, S.; Pöllath, C.; Hansen, U.; Rizzo, F.; Krafft, C.; Westermann, M.; Hussain, M.; Peters, G.; Pletz, M.W.; et al. The Staphylococcus aureus extracellular matrix protein (Emp) has a fibrous structure and binds to different extracellular matrices. Sci. Rep. 2017, 7, 13665. [Google Scholar] [CrossRef] [Green Version]
- Araujo-Alves, A.V.; Kraychete, G.B.; Gilmore, M.S.; Barros, E.M.; Giambiagi-Demarval, M. shsA: A novel orthologous of sasX/sesI virulence genes is detected in Staphylococcus haemolyticus Brazilian strains. Infect. Genet. Evol. 2022, 97, 105189. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Du, X.; Villaruz, A.E.; Diep, B.A.; Wang, D.; Song, Y.; Tian, Y.; Hu, J.; Yu, F.; Lu, Y.; et al. MRSA epidemic linked to a quickly spreading colonization and virulence determinant. Nat. Med. 2012, 18, 816–819. [Google Scholar] [CrossRef]
- Liu, Q.; Du, X.; Hong, X.; Li, T.; Zheng, B.; He, L.; Wang, Y.; Otto, M.; Li, M. Targeting Surface Protein SasX by Active and Passive Vaccination to Reduce Staphylococcus aureus Colonization and Infection. Infect. Immun. 2015, 83, 2168–2174. [Google Scholar] [CrossRef] [Green Version]
- Foster, T.J. Immune evasion by staphylococci. Nat. Rev. Genet. 2005, 3, 948–958. [Google Scholar] [CrossRef]
- Goodyear, C.S.; Silverman, G.J. Death by a B Cell Superantigen: In vivo VH-targeted apoptotic supraclonal B cell deletion by a Staphylococcal Toxin. J. Exp. Med. 2003, 197, 1125–1139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claßen, A.; Kalali, B.N.; Schnopp, C.; Andres, C.; Aguilar-Pimentel, J.A.; Ring, J.; Ollert, M.; Mempel, M. TNF receptor I on human keratinocytes is a binding partner for staphylococcal protein A resulting in the activation of NF kappa B, AP-1, and downstream gene transcription. Exp. Dermatol. 2011, 20, 48–52. [Google Scholar] [CrossRef]
- Kozman, A.; Yao, Y.; Bina, P.; Saha, C.; Yao, W.; Kaplan, M.H.; Travers, J.B. Encoding a superantigen by Staphylococcus aureus does not affect clinical characteristics of infected atopic dermatitis lesions. Br. J. Dermatol. 2010, 163, 1308–1311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jun, S.H.; Lee, J.H.; Kim, S.I.; Choi, C.W.; Park, T.I.; Jung, H.R.; Cho, J.W.; Kim, S.H.; Lee, J.C. Staphylococcus aureus -derived membrane vesicles exacerbate skin inflammation in atopic dermatitis. Clin. Exp. Allergy 2017, 47, 85–96. [Google Scholar] [CrossRef] [PubMed]
- Ledo, C.; Gonzalez, C.D.; Garofalo, A.; Sabbione, F.; Keitelman, I.A.; Giai, C.; Stella, I.; Trevani, A.S.; Gómez, M.I. Protein A Modulates Neutrophil and Keratinocyte Signaling and Survival in Response to Staphylococcus aureus. Front. Immunol. 2020, 11, 524180. [Google Scholar] [CrossRef]
- Soong, G.; Chun, J.; Parker, D.; Prince, A. Staphylococcus aureus Activation of Caspase 1/Calpain Signaling Mediates Invasion Through Human Keratinocytes. J. Infect. Dis. 2012, 205, 1571–1579. [Google Scholar] [CrossRef] [Green Version]
- Cruz, A.R.; Bentlage, A.E.H.; Blonk, R.; de Haas, C.J.C.; Aerts, P.C.; Scheepmaker, L.M.; Bouwmeester, I.G.; Lux, A.; van Strijp, J.A.G.; Nimmerjahn, F.; et al. Toward Understanding How Staphylococcal Protein a Inhibits IgG-Mediated Phagocytosis. J. Immunol. 2022, 209, 1146–1155. [Google Scholar] [CrossRef]
- Cruz, A.R.; den Boer, M.A.; Strasser, J.; Zwarthoff, S.A.; Beurskens, F.J.; de Haas, C.J.C.; Aerts, P.C.; Wang, G.; de Jong, R.N.; Bagnoli, F.; et al. Staphylococcal protein A inhibits complement activation by interfering with IgG hexamer formation. Proc. Natl. Acad. Sci. USA 2021, 118, e2016772118. [Google Scholar] [CrossRef]
- Corrigan, R.M.; Miajlovic, H.; Foster, T.J. Surface proteins that promote adherence of Staphylococcus aureus to human desquamated nasal epithelial cells. BMC Microbiol. 2009, 9, 22. [Google Scholar] [CrossRef] [Green Version]
- Gotz, F. Staphylococcus and biofilms. Mol. Microbiol. 2002, 43, 1367–1378. [Google Scholar] [CrossRef]
- Cramton, S.E.; Ulrich, M.; Götz, F.; Döring, G. Anaerobic Conditions Induce Expression of Polysaccharide Intercellular Adhesin in Staphylococcus aureus and Staphylococcus epidermidis. Infect. Immun. 2001, 69, 4079–4085. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, E.; Pozzi, C.; Houston, P.; Smyth, D.; Humphreys, H.; Robinson, D.A.; O’Gara, J.P. Association between Methicillin Susceptibility and Biofilm Regulation in Staphylococcus aureus Isolates from Device-Related Infections. J. Clin. Microbiol. 2007, 45, 1379–1388. [Google Scholar] [CrossRef] [Green Version]
- Merino, N.; Toledo-Arana, A.; Vergara-Irigaray, M.; Valle, J.; Solano, C.; Calvo, E.; Lopez, J.A.; Foster, T.J.; Penadés, J.R.; Lasa, I. Protein A-Mediated Multicellular Behavior in Staphylococcus aureus. J. Bacteriol. 2009, 191, 832–843. [Google Scholar] [CrossRef] [Green Version]
- Houston, P.; Rowe, S.E.; Pozzi, C.; Waters, E.M.; O’Gara, J.P. Essential Role for the Major Autolysin in the Fibronectin-Binding Protein-Mediated Staphylococcus aureus Biofilm Phenotype. Infect. Immun. 2011, 79, 1153–1165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, K.C.; Mann, E.E.; Endres, J.L.; Weiss, E.C.; Cassat, J.E.; Smeltzer, M.S.; Bayles, K.W. The cidA murein hydrolase regulator contributes to DNA release and biofilm development in Staphylococcus aureus. Proc. Natl. Acad. Sci. USA 2007, 104, 8113–8118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cucarella, C.; Solano, C.; Valle, J.; Amorena, B.; Lasa, Í.; Penadés, J.R. Bap, a Staphylococcus aureus Surface Protein Involved in Biofilm Formation. J. Bacteriol. 2001, 183, 2888–2896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hennig, S.; Wai, S.N.; Ziebuhr, W. Spontaneous switch to PIA-independent biofilm formation in an ica-positive Staphylococcus epidermidis isolate. Int. J. Med. Microbiol. 2007, 297, 117–122. [Google Scholar] [CrossRef]
- Rohde, H.; Burandt, E.C.; Siemssen, N.; Frommelt, L.; Burdelski, C.; Wurster, S.; Scherpe, S.; Davies, A.P.; Harris, L.G.; Horstkotte, M.A.; et al. Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prosthetic hip and knee joint infections. Biomaterials 2007, 28, 1711–1720. [Google Scholar] [CrossRef]
- Chokr, A.; Watier, D.; Eleaume, H.; Pangon, B.; Ghnassia, J.-C.; Mack, D.; Jabbouri, S. Correlation between biofilm formation and production of polysaccharide intercellular adhesin in clinical isolates of coagulase-negative staphylococci. Int. J. Med. Microbiol. 2006, 296, 381–388. [Google Scholar] [CrossRef]
- Doern, C.D.; Roberts, A.L.; Hong, W.; Nelson, J.; Lukomski, S.; Swords, W.E.; Reid, S.D. Biofilm formation by group A Streptococcus: A role for the streptococcal regulator of virulence (Srv) and streptococcal cysteine protease (SpeB). Microbiology 2009, 155, 46–52. [Google Scholar] [CrossRef] [Green Version]
- Manetti, A.G.; Zingaretti, C.; Falugi, F.; Capo, S.; Bombaci, M.; Bagnoli, F.; Gambellini, G.; Bensi, G.; Mora, M.; Edwards, A.M.; et al. Streptococcus pyogenes pili promote pharyngeal cell adhesion and biofilm formation. Mol. Microbiol. 2007, 64, 968–983. [Google Scholar] [CrossRef] [PubMed]
- Courtney, H.S.; Ofek, I.; Penfound, T.; Nizet, V.; Pence, M.A.; Kreikemeyer, B.; Podbielbski, A.; Hasty, D.L.; Dale, J.B. Relationship between Expression of the Family of M Proteins and Lipoteichoic Acid to Hydrophobicity and Biofilm Formation in Streptococcus pyogenes. PLoS ONE 2009, 4, e4166. [Google Scholar] [CrossRef]
- Ridder, M.J.; McReynolds, A.K.G.; Dai, H.; Pritchard, M.T.; Markiewicz, M.A.; Bose, J.L. Kinetic Characterization of the Immune Response to Methicillin-Resistant Staphylococcus aureus Subcutaneous Skin Infection. Infect. Immun. 2022, 90, e0006522. [Google Scholar] [CrossRef]
- Wang, X.; Li, X.; Chen, L.; Yuan, B.; Liu, T.; Dong, Q.; Liu, Y.; Yin, H. Interleukin-33 facilitates cutaneous defense against Staphylococcus aureus by promoting the development of neutrophil extracellular trap. Int. Immunopharmacol. 2020, 81, 106256. [Google Scholar] [CrossRef]
- Leyva-Castillo, J.-M.; Das, M.; Kane, J.; Strakosha, M.; Singh, S.; Wong, D.S.H.; Horswill, A.R.; Karasuyama, H.; Brombacher, F.; Miller, L.S.; et al. Basophil-derived IL-4 promotes cutaneous Staphylococcus aureus infection. J. Clin. Investig. 2021, 6, e149953. [Google Scholar] [CrossRef]
- Brady, R.A.; Mocca, C.P.; Plaut, R.D.; Takeda, K.; Burns, D.L. Comparison of the immune response during acute and chronic Staphylococcus aureus infection. PLoS ONE 2018, 13, e0195342. [Google Scholar] [CrossRef] [Green Version]
- Kisich, K.O.; Howell, M.D.; Boguniewicz, M.; Heizer, H.R.; Watson, N.U.; Leung, D.Y. The Constitutive Capacity of Human Keratinocytes to Kill Staphylococcus aureus Is Dependent on β-Defensin 3. J. Investig. Dermatol. 2007, 127, 2368–2380. [Google Scholar] [CrossRef] [Green Version]
- Ngo, Q.V.; Faass, L.; Sähr, A.; Hildebrand, D.; Eigenbrod, T.; Heeg, K.; Nurjadi, D. Inflammatory Response Against Staphylococcus aureus via Intracellular Sensing of Nucleic Acids in Keratinocytes. Front. Immunol. 2022, 13, 828626. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Limjunyawong, N.; Sypek, E.I.; Wang, G.; Ortines, R.V.; Youn, C.; Alphonse, M.P.; Dikeman, D.; Wang, Y.; Lay, M.; et al. Keratinocyte-derived defensins activate neutrophil-specific receptors Mrgpra2a/b to prevent skin dysbiosis and bacterial infection. Immunity 2022, 55, 1645–1662.e1647. [Google Scholar] [CrossRef] [PubMed]
- Hanzelmann, D.; Joo, H.S.; Franz-Wachtel, M.; Hertlein, T.; Stevanovic, S.; Macek, B.; Wolz, C.; Götz, F.; Otto, M.; Kretschmer, D.; et al. Toll-like receptor 2 activation depends on lipopeptide shedding by bacterial surfactants. Nat. Commun. 2016, 7, 12304. [Google Scholar] [CrossRef]
- Hruz, P.; Zinkernagel, A.S.; Jenikova, G.; Botwin, G.J.; Hugot, J.-P.; Karin, M.; Nizet, V.; Eckmann, L. NOD2 contributes to cutaneous defense against Staphylococcus aureus through α-toxin-dependent innate immune activation. Proc. Natl. Acad. Sci. USA 2009, 106, 12873–12878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dalen, R.; De la Cruz Diaz, J.S.; Rumpret, M.; Fuchsberger, F.F.; van Teijlingen, N.H.; Hanske, J.; Rademacher, C.; Geijtenbeek, T.B.H.; van Strijp, J.A.G.; Weidenmaier, C.; et al. Langerhans Cells Sense Staphylococcus aureus Wall Teichoic Acid through Langerin to Induce Inflammatory Responses. mBio 2019, 10, e00330-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pires, S.; Parker, D. IL-1β activation in response to Staphylococcus aureus lung infection requires inflammasome-dependent and independent mechanisms. Eur. J. Immunol. 2018, 48, 1707–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.S.; Guo, Y.; Ramos, R.I.; Hebroni, F.; Plaisier, S.B.; Xuan, C.; Granick, J.L.; Matsushima, H.; Takashima, A.; Iwakura, Y.; et al. Neutrophil-derived IL-1β Is Sufficient for Abscess Formation in Immunity against Staphylococcus aureus in Mice. PLoS Pathog. 2012, 8, e1003047. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.S.; O’Connell, R.M.; Gutierrez, M.A.; Pietras, E.M.; Shahangian, A.; Gross, C.E.; Thirumala, A.; Cheung, A.L.; Cheng, G.; Modlin, R.L. MyD88 Mediates Neutrophil Recruitment Initiated by IL-1R but Not TLR2 Activation in Immunity against Staphylococcus aureus. Immunity 2006, 24, 79–91. [Google Scholar] [CrossRef] [Green Version]
- Feuerstein, R.; Seidl, M.; Prinz, M.; Henneke, P. MyD88 in Macrophages Is Critical for Abscess Resolution in Staphylococcal Skin Infection. J. Immunol. 2015, 194, 2735–2745. [Google Scholar] [CrossRef] [Green Version]
- Castleman, M.J.; Febbraio, M.; Hall, P.R. CD36 Is Essential for Regulation of the Host Innate Response to Staphylococcus aureus α-Toxin–Mediated Dermonecrosis. J. Immunol. 2015, 195, 2294–2302. [Google Scholar] [CrossRef] [Green Version]
- Özcan, A.; Collado-Diaz, V.; Egholm, C.; Tomura, M.; Gunzer, M.; Halin, C.; Kolios, A.G.A.; Boyman, O. CCR7-guided neutrophil redirection to skin-draining lymph nodes regulates cutaneous inflammation and infection. Sci. Immunol. 2022, 7, eabi9126. [Google Scholar] [CrossRef]
- Montgomery, C.P.; Daniels, M.; Zhao, F.; Alegre, M.-L.; Chong, A.S.; Daum, R.S. Protective Immunity against Recurrent Staphylococcus aureus Skin Infection Requires Antibody and Interleukin-17A. Infect. Immun. 2014, 82, 2125–2134. [Google Scholar] [CrossRef] [Green Version]
- Beesetty, P.; Si, Y.; Li, Z.; Yang, C.; Zhao, F.; Chong, A.S.; Montgomery, C.P. Tissue specificity drives protective immunity against Staphylococcus aureus infection. Front. Immunol. 2022, 13, 795792. [Google Scholar] [CrossRef]
- Starkl, P.; Watzenboeck, M.L.; Popov, L.M.; Zahalka, S.; Hladik, A.; Lakovits, K.; Radhouani, M.; Haschemi, A.; Marichal, T.; Reber, L.L.; et al. IgE Effector Mechanisms, in Concert with Mast Cells, Contribute to Acquired Host Defense against Staphylococcus aureus. Immunity 2020, 53, 793–804. [Google Scholar] [CrossRef]
- Marchitto, M.C.; Dillen, C.A.; Liu, H.; Miller, R.J.; Archer, N.K.; Ortines, R.V.; Alphonse, M.P.; Marusina, A.I.; Merleev, A.A.; Wang, Y.; et al. Clonal Vγ6 + Vδ4 + T cells promote IL-17–mediated immunity against Staphylococcus aureus skin infection. Proc. Natl. Acad. Sci. USA 2019, 116, 10917–10926. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.S.; Pietras, E.M.; Garcia, N.C.; Ramos, R.I.; Farzam, D.M.; Monroe, H.R.; Magorien, J.E.; Blauvelt, A.; Kolls, J.K.; Cheung, A.L.; et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Investig. 2010, 120, 1762–1773. [Google Scholar] [CrossRef] [Green Version]
- Dillen, C.A.; Pinsker, B.L.; Marusina, A.I.; Merleev, A.A.; Farber, O.N.; Liu, H.; Archer, N.; Lee, D.B.; Wang, Y.; Ortines, R.V.; et al. Clonally expanded γδ T cells protect against Staphylococcus aureus skin reinfection. J. Clin. Investig. 2018, 128, 1026–1042. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, A.S.; Hemmers, S.; Garijo, O.; Chabod, M.; Mowen, K.; Witherden, D.A.; Havran, W.L. Dendritic epidermal T cells regulate skin antimicrobial barrier function. J. Clin. Investig. 2013, 123, 4364–4374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, C.; Wuethrich, B.; Matter, L.A.; Wilhelm, J.; Schopfer, K. Immune Response to Staphylococcus aureus in Atopic Dermatitis. Dermatology 1985, 170, 114–120. [Google Scholar] [CrossRef]
- Farag, A.K.; Roesner, L.M.; Wieschowski, S.; Heratizadeh, A.; Eiz-Vesper, B.; Kwok, W.W.; Valenta, R.; Werfel, T. Specific T cells targeting Staphylococcus aureus fibronectin-binding protein 1 induce a type 2/type 1 inflammatory response in sensitized atopic dermatitis patients. Allergy 2022, 77, 1245–1253. [Google Scholar] [CrossRef]
- Brown, N.M.; Goodman, A.L.; Horner, C.; Jenkins, A.; Brown, E.M. Treatment of methicillin-resistant Staphylococcus aureus (MRSA): Updated guidelines from the UK. JAC-Antimicrob. Resist. 2021, 3, dlaa114. [Google Scholar] [CrossRef] [PubMed]
- Stevens, D.L.; Bisno, A.L.; Chambers, H.F.; Dellinger, E.P.; Goldstein, E.J.C.; Gorbach, S.L.; Hirschmann, J.; Kaplan, S.L.; Montoya, J.G.; Wade, J.C.; et al. Practice guidelines for the diagnosis and management of skin and soft tissue infections: 2014 update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2014, 59, e10–e52. [Google Scholar] [CrossRef] [Green Version]
- Esposito, S.; Bassetti, M.; Concia, E.; De Simone, G.; De Rosa, F.G.; Grossi, P.; Novelli, A.; Menichetti, F.; Petrosillo, N.; Tinelli, M.; et al. Diagnosis and management of skin and soft-tissue infections (SSTI). A literature review and consensus statement: An update. J. Chemother. 2017, 29, 197–214. [Google Scholar] [CrossRef]
- Sartelli, M.; Guirao, X.; Hardcastle, T.C.; Kluger, Y.; Boermeester, M.A.; Raşa, K.; Ansaloni, L.; Coccolini, F.; Montravers, P.; Abu-Zidan, F.M.; et al. 2018 WSES/SIS-E consensus conference: Recommendations for the management of skin and soft-tissue infections. World J. Emerg. Surg. 2018, 13, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duane, T.M.; Huston, J.M.; Collom, M.; Beyer, A.; Parli, S.; Buckman, S.; Shapiro, M.; McDonald, A.; Diaz, J.; Tessier, J.M.; et al. Surgical Infection Society 2020 Updated Guidelines on the Management of Complicated Skin and Soft Tissue Infections. Surg. Infect. 2021, 22, 383–399. [Google Scholar] [CrossRef] [PubMed]
- US Food and Drug Administration. Acute Bacterial Skin and Skin Structure Infections: Developing Drugs for Treatment; United States Food and Drug Administration: Silver Spring, MD, USA, 2013. [Google Scholar]
- Hindy, J.-R.; Haddad, S.F.; Kanj, S.S. New drugs for methicillin-resistant Staphylococcus aureus skin and soft tissue infections. Curr. Opin. Infect. Dis. 2022, 35, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Hatlen, T.J.; Miller, L.G. Staphylococcal Skin and Soft Tissue Infections. Infect. Dis. Clin. N. Am. 2020, 35, 81–105. [Google Scholar] [CrossRef]
- Bouza, E.; Burillo, A. Current international and national guidelines for managing skin and soft tissue infections. Curr. Opin. Infect. Dis. 2022, 35, 61–71. [Google Scholar] [CrossRef]
- Gilbert, D.N.; Chambers, H.F.; Saag, M.S.; Pavia, A.T.; Boucher, H.W. The Sanford Guide to Antimicrobial Therapy; BI Publications Pvt Ltd: Sperryville, VA, USA, 2022. [Google Scholar]
- González, C.; Rubio, M.; Romero-Vivas, J.; González, M.; Picazo, J.J. Bacteremic Pneumonia Due to Staphylococcus aureus: A Comparison of Disease Caused by Methicillin-Resistant and Methicillin-Susceptible Organisms. Clin. Infect. Dis. 1999, 29, 1171–1177. [Google Scholar] [CrossRef] [Green Version]
- Chang, F.-Y.; Peacock, J.E.; Musher, D.M.; Triplett, P.; MacDonald, B.B.; Mylotte, J.M.; O’Donnell, A.; Wagener, M.M.; Yu, V.L. Staphylococcus aureus Bacteremia: Recurrence and the impact of antibiotic treatment in a prospective multicenter study. Medicine 2003, 82, 333–339. [Google Scholar] [CrossRef]
- Bowen, A.C.; Carapetis, J.R.; Currie, B.J.; Fowler, V., Jr.; Chambers, H.F.; Tong, S.Y.C. Sulfamethoxazole-Trimethoprim (Cotrimoxazole) for Skin and Soft Tissue Infections Including Impetigo, Cellulitis, and Abscess. Open Forum Infect. Dis. 2017, 4, ofx232. [Google Scholar] [CrossRef] [Green Version]
- Pallin, D.J.; Binder, W.D.; Allen, M.B.; Lederman, M.; Parmar, S.; Filbin, M.R.; Hooper, D.C.; Camargo, C.A., Jr. Clinical Trial: Comparative Effectiveness of Cephalexin Plus Trimethoprim-Sulfamethoxazole Versus Cephalexin Alone for Treatment of Uncomplicated Cellulitis: A Randomized Controlled Trial. Clin. Infect. Dis. 2013, 56, 1754–1762. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.G.; Daum, R.S.; Creech, C.B.; Young, D.; Downing, M.D.; Eells, S.J.; Pettibone, S.; Hoagland, R.J.; Chambers, H.F. Clindamycin versus Trimethoprim–Sulfamethoxazole for Uncomplicated Skin Infections. N. Engl. J. Med. 2015, 372, 1093–1103. [Google Scholar] [CrossRef] [Green Version]
- Moran, G.J.; Krishnadasan, A.; Mower, W.R.; Abrahamian, F.M.; LoVecchio, F.; Steele, M.T.; Rothman, R.E.; Karras, D.J.; Hoagland, R.; Pettibone, S.; et al. Effect of Cephalexin Plus Trimethoprim-Sulfamethoxazole vs Cephalexin Alone on Clinical Cure of Uncomplicated Cellulitis: A Randomized Clinical Trial. JAMA 2017, 317, 2088–2096. [Google Scholar] [CrossRef] [PubMed]
- Diekema, D.J.; Pfaller, M.A.; Shortridge, D.; Zervos, M.; Jones, R.N. Twenty-Year Trends in Antimicrobial Susceptibilities Among Staphylococcus aureus from the SENTRY Antimicrobial Surveillance Program. Open Forum Infect. Dis. 2019, 6, S47–S53. [Google Scholar] [CrossRef] [PubMed]
- Lexicomp Online. 2023. Available online: https://online.lexi.com (accessed on 1 March 2023).
- Levin, T.P.; Suh, B.; Axelrod, P.; Truant, A.L.; Fekete, T. Potential Clindamycin Resistance in Clindamycin-Susceptible, Erythromycin-Resistant Staphylococcus aureus: Report of a Clinical Failure. Antimicrob. Agents Chemother. 2005, 49, 1222–1224. [Google Scholar] [CrossRef] [Green Version]
- Steward, C.D.; Raney, P.M.; Morrell, A.K.; Williams, P.P.; McDougal, L.K.; Jevitt, L.; McGowan, J.E., Jr.; Tenover, F.C. Testing for Induction of Clindamycin Resistance in Erythromycin-Resistant Isolates of Staphylococcus aureus. J. Clin. Microbiol. 2005, 43, 1716–1721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaPlante, K.L.; Leonard, S.N.; Andes, D.R.; Craig, W.A.; Rybak, M.J. Activities of Clindamycin, Daptomycin, Doxycycline, Linezolid, Trimethoprim-Sulfamethoxazole, and Vancomycin against Community-Associated Methicillin-Resistant Staphylococcus aureus with Inducible Clindamycin Resistance in Murine Thigh Infection and In Vitro Pharmacodynamic Models. Antimicrob. Agents Chemother. 2008, 52, 2156–2162. [Google Scholar] [CrossRef] [Green Version]
- Siberry, G.K.; Tekle, T.; Carroll, K.; Dick, J. Failure of Clindamycin Treatment of Methicillin-Resistant Staphylococcus aureus Expressing Inducible Clindamycin Resistance In Vitro. Clin. Infect. Dis. 2003, 37, 1257–1260. [Google Scholar] [CrossRef] [PubMed]
- Cenizal, M.J.; Skiest, D.; Luber, S.; Bedimo, R.; Davis, P.; Fox, P.; Delaney, K.; Hardy, R.D. Prospective Randomized Trial of Empiric Therapy with Trimethoprim-Sulfamethoxazole or Doxycycline for Outpatient Skin and Soft Tissue Infections in an Area of High Prevalence of Methicillin-Resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 2628–2630. [Google Scholar] [CrossRef] [Green Version]
- Itani, K.M.; Dryden, M.S.; Bhattacharyya, H.; Kunkel, M.J.; Baruch, A.M.; Weigelt, J.A. Efficacy and safety of linezolid versus vancomycin for the treatment of complicated skin and soft-tissue infections proven to be caused by methicillin-resistant Staphylococcus aureus. Am. J. Surg. 2010, 199, 804–816. [Google Scholar] [CrossRef]
- Weigelt, J.; Itani, K.; Stevens, D.; Lau, W.; Dryden, M.; Knirsch, C.; Linezolid, C.S.G. Linezolid versus Vancomycin in Treatment of Complicated Skin and Soft Tissue Infections. Antimicrob. Agents Chemother. 2005, 49, 2260–2266. [Google Scholar] [CrossRef] [Green Version]
- Prokocimer, P.; De Anda, C.; Fang, E.; Mehra, P.; Das, A. Tedizolid Phosphate vs Linezolid for Treatment of Acute Bacterial Skin and Skin Structure Infections: The ESTABLISH-1 randomized trial. JAMA 2013, 309, 559–569. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.; Dong, B.R.; Yang, M.; Chen, X.; Wu, T.; Liu, G.J. Linezolid versus vancomycin for skin and soft tissue infections. Evid. -Based Child Health A Cochrane Rev. J. 2013, 9, 103–166. [Google Scholar] [CrossRef] [PubMed]
- McCool, R.; Gould, I.M.; Eales, J.; Barata, T.; Arber, M.; Fleetwood, K.; Glanville, J.; Kauf, T.L. Systematic review and network meta-analysis of tedizolid for the treatment of acute bacterial skin and skin structure infections caused by MRSA. BMC Infect. Dis. 2017, 17, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomczak, H.; Szałek, E.; Błażejewska, W.; Myczko, K.; Horla, A.; Grześkowiak, E. The need to assay the real MIC when making the decision to eradicate Staphylococcus aureus with vancomycin. Postep. Hig. Med. Dosw. (Online) 2013, 67, 921–925. [Google Scholar] [CrossRef] [PubMed]
- Rossatto, F.C.P.; Proença, L.A.; Becker, A.P.; Silveira, A.C.D.O.; Caierão, J.; D’Azevedo, P.A. Evaluation of Methods in Detecting Vancomycin Mic among Mrsa Isolates and the Changes in Accuracy Related to Different Mic Values. Rev. Inst. Med. Trop. Sao Paulo 2014, 56, 469–472. [Google Scholar] [CrossRef]
- Kalil, A.C.; Van Schooneveld, T.C.; Fey, P.D.; Rupp, M.E. Association Between Vancomycin Minimum Inhibitory Concentration and Mortality Among Patients with Staphylococcus aureus Bloodstream Infections: A systematic review and meta-analysis. JAMA 2014, 312, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Dhand, A.; Sakoulas, G. Reduced vancomycin susceptibility among clinical Staphylococcus aureus isolates (‘the MIC Creep’): Implications for therapy. F1000 Med. Rep. 2012, 4, 4. [Google Scholar] [CrossRef]
- Diaz, R.; Afreixo, V.; Ramalheira, E.; Rodrigues, C.; Gago, B. Evaluation of vancomycin MIC creep in methicillin-resistant Staphylococcus aureus infections—A systematic review and meta-analysis. Clin. Microbiol. Infect. 2018, 24, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Rybak, M.J.; Le, J.; Lodise, T.P.; Levine, D.P.; Bradley, J.S.; Liu, C.; Mueller, B.A.; Pai, M.P.; Wong-Beringer, A.; Rotschafer, J.C.; et al. Executive Summary: Therapeutic Monitoring of Vancomycin for Serious Methicillin-Resistant Staphylococcus aureus Infections: A Revised Consensus Guideline and Review of the American Society of Health-System Pharmacists, the Infectious Diseases Society of America, the Pediatric Infectious Diseases Society, and the Society of Infectious Diseases Pharmacists. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2020, 40, 363–367. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Y.; Hu, J.T.; Zhang, C.; Zhou, W.; Chen, X.F.; Jiang, L.Y.; Tang, Z.H. The safety and efficacy of daptomycin versus other antibiotics for skin and soft-tissue infections: A meta-analysis of randomised controlled trials. BMJ Open 2014, 4, e004744. [Google Scholar] [CrossRef] [Green Version]
- Zhou, F.; Liu, C.; Mao, Z.; Yang, M.; Kang, H.; Liu, H.; Pan, L.; Hu, J.; Luo, J. Efficacy and safety of daptomycin for skin and soft tissue infections: A systematic review with trial sequential analysis. Ther. Clin. Risk Manag. 2016, 12, 1455–1466. [Google Scholar] [CrossRef] [Green Version]
- Youngster, I.; Shenoy, E.S.; Hooper, D.C.; Nelson, S.B. Comparative Evaluation of the Tolerability of Cefazolin and Nafcillin for Treatment of Methicillin-Susceptible Staphylococcus aureus Infections in the Outpatient Setting. Clin. Infect. Dis. 2014, 59, 369–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campioli, C.C.; Go, J.R.; Abu Saleh, O.; Challener, D.; Yetmar, Z.; Osmon, D.R. Antistaphylococcal Penicillin vs Cefazolin for the Treatment of Methicillin-Susceptible Staphylococcus aureus Spinal Epidural Abscesses. Open Forum Infect. Dis. 2021, 8, ofab071. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R.; Seas, C.; Carvajal, L.P.; Diaz, L.; Echeverri, A.M.; Ferro, C.; Rios, R.; Porras, P.; Luna, C.; Gotuzzo, E.; et al. The Cefazolin Inoculum Effect Is Associated with Increased Mortality in Methicillin-Susceptible Staphylococcus aureus Bacteremia. Open Forum Infect. Dis. 2018, 5, ofy123. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. Assessment Report: Zinforo; European Medicines Agency: London, UK, 2012. [Google Scholar]
- Chen, C.-Y.; Chen, W.-C.; Lai, C.-C.; Shih, T.-P.; Tang, H.-J. Anti-MRSA Cephalosporin versus Vancomycin-Based Treatment for Acute Bacterial Skin and Skin Structure Infection: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Antibiotics 2021, 10, 1020. [Google Scholar] [CrossRef] [PubMed]
- Holland, T.L.; Bayer, A.S.; Fowler, V.G. Persistent methicillin-Resistant Staphylococcus aureus Bacteremia: Resetting the Clock for Optimal Management. Clin. Infect. Dis. 2022, 75, 1668–1674. [Google Scholar] [CrossRef] [PubMed]
- Zasowski, E.J.; Trinh, T.D.; Claeys, K.C.; Casapao, A.M.; Sabagha, N.; Lagnf, A.M.; Klinker, K.P.; Davis, S.L.; Rybak, M.J. Multicenter Observational Study of Ceftaroline Fosamil for Methicillin-Resistant Staphylococcus aureus Bloodstream Infections. Antimicrob. Agents Chemother. 2017, 61, e02015-1. [Google Scholar] [CrossRef] [Green Version]
- Molina, K.C.; Morrisette, T.; Miller, M.A.; Huang, V.; Fish, D.N. The Emerging Role of β-Lactams in the Treatment of Methicillin-Resistant Staphylococcus aureus Bloodstream Infections. Antimicrob. Agents Chemother. 2020, 64, e00468-20. [Google Scholar] [CrossRef]
- Geriak, M.; Haddad, F.; Rizvi, K.; Rose, W.; Kullar, R.; LaPlante, K.; Yu, M.; Vasina, L.; Ouellette, K.; Zervos, M.; et al. Clinical Data on Daptomycin plus Ceftaroline versus Standard of Care Monotherapy in the Treatment of Methicillin-Resistant Staphylococcus aureus Bacteremia. Antimicrob. Agents Chemother. 2019, 63, e02483-18. [Google Scholar] [CrossRef] [Green Version]
- Tong, S.Y.C.; Mora, J.; Bowen, A.C.; Cheng, M.P.; Daneman, N.; Goodman, A.L.; Heriot, G.S.; Lee, T.C.; Lewis, R.J.; Lye, D.C.; et al. The Staphylococcus aureus Network Adaptive Platform Trial Protocol: New Tools for an Old Foe. Clin. Infect. Dis. 2022, 75, 2027–2034. [Google Scholar] [CrossRef]
- Rubino, C.M.; Van Wart, S.A.; Bhavnani, S.M.; Ambrose, P.G.; McCollam, J.S.; Forrest, A. Oritavancin Population Pharmacokinetics in Healthy Subjects and Patients with Complicated Skin and Skin Structure Infections or Bacteremia. Antimicrob. Agents Chemother. 2009, 53, 4422–4428. [Google Scholar] [CrossRef] [Green Version]
- Bowker, K.E.; Noel, A.R.; MacGowan, A.P. Pharmacodynamics of dalbavancin studied in an in vitro pharmacokinetic system. J. Antimicrob. Chemother. 2006, 58, 802–805. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.-K.; Chen, C.-Y.; Chen, W.-C.; Chao, C.-M.; Lai, C.-C. Clinical efficacy and safety of novel lipoglycopeptides in the treatment of acute bacterial skin and skin structure infections: A systematic review and meta-analysis of randomized controlled trials. Expert Rev. Anti-Infect. Ther. 2022, 20, 435–444. [Google Scholar] [CrossRef] [PubMed]
- De Vito, A.; Fiore, V.; Colpani, A.; Zauli, B.; Fanelli, C.; Tiseo, G.; Occhineri, S.; Babudieri, S.; Falcone, M.; Madeddu, G. The current and future off-label uses of dalbavancin: A narrative review. Eur. Rev. Med. Pharm. Sci. 2023, 27, 1222–1238. [Google Scholar] [CrossRef]
- Turner, N.A.; Zaharoff, S.; King, H.; Evans, S.; Hamasaki, T.; Lodise, T.; Ghazaryan, V.; Beresnev, T.; Riccobene, T.; Patel, R.; et al. Dalbavancin as an option for treatment of S. aureus bacteremia (DOTS): Study protocol for a phase 2b, multicenter, randomized, open-label clinical trial. Trials 2022, 23, 407. [Google Scholar] [CrossRef]
- Bidell, M.R.; Lodise, T.P. Use of oral tetracyclines in the treatment of adult outpatients with skin and skin structure infections: Focus on doxycycline, minocycline, and omadacycline. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2021, 41, 915–931. [Google Scholar] [CrossRef]
- O’Riordan, W.; McManus, A.J.; Teras, J.; Poromanski, I.; Cruz-Saldariagga, M.; Quintas, M.; Lawrence, L.; Liang, S.; Cammarata, S.; PROCEED Study Group; et al. A Comparison of the Efficacy and Safety of Intravenous Followed by Oral Delafloxacin With Vancomycin Plus Aztreonam for the Treatment of Acute Bacterial Skin and Skin Structure Infections: A Phase 3, Multinational, Double-Blind, Randomized Study. Clin. Infect. Dis. 2018, 67, 657–666. [Google Scholar] [CrossRef] [PubMed]
- Pullman, J.; Gardovskis, J.; Farley, B.; Sun, E.; Quintas, M.; Lawrence, L.; Ling, R.; Cammarata, S. PROCEED Study Group Efficacy and safety of delafloxacin compared with vancomycin plus aztreonam for acute bacterial skin and skin structure infections: A Phase 3, double-blind, randomized study. J. Antimicrob. Chemother. 2017, 72, 3471–3480. [Google Scholar] [CrossRef] [Green Version]
- McCurdy, S.; Lawrence, L.; Quintas, M.; Woosley, L.; Flamm, R.; Tseng, C.; Cammarata, S. In Vitro Activity of Delafloxacin and Microbiological Response against Fluoroquinolone-Susceptible and Nonsusceptible Staphylococcus aureus Isolates from Two Phase 3 Studies of Acute Bacterial Skin and Skin Structure Infections. Antimicrob. Agents Chemother. 2017, 61, e00772-17. [Google Scholar] [CrossRef] [Green Version]
- Baggio, D.; Ananda-Rajah, M.R. Fluoroquinolone antibiotics and adverse events. Aust. Prescr. 2021, 44, 161–164. [Google Scholar] [CrossRef]
- Wald-Dickler, N.; Holtom, P.D.; Phillips, M.C.; Centor, R.M.; Lee, R.A.; Baden, R.; Spellberg, B. Oral Is the New IV. Challenging Decades of Blood and Bone Infection Dogma: A Systematic Review. Am. J. Med. 2021, 135, 369–379. [Google Scholar] [CrossRef]
- Lee, R.A.; Stripling, J.T.; Spellberg, B.; Centor, R.M. Short-course antibiotics for common infections: What do we know and where do we go from here? Clin. Microbiol. Infect. 2023, 29, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Moran, G.J.; Fang, E.; Corey, G.R.; Das, A.F.; De Anda, C.; Prokocimer, P. Tedizolid for 6 days versus linezolid for 10 days for acute bacterial skin and skin-structure infections (ESTABLISH-2): A randomised, double-blind, phase 3, non-inferiority trial. Lancet Infect. Dis. 2014, 14, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Hepburn, M.J.; Dooley, D.P.; Skidmore, P.J.; Ellis, M.W.; Starnes, W.F.; Hasewinkle, W.C. Comparison of Short-Course (5 Days) and Standard (10 Days) Treatment for Uncomplicated Cellulitis. Arch. Intern. Med. 2004, 164, 1669–1674. [Google Scholar] [CrossRef] [PubMed]
- Suh, G.A.; Lodise, T.P.; Tamma, P.D.; Knisely, J.M.; Alexander, J.; Aslam, S.; Barton, K.D.; Bizzell, E.; Totten, K.M.C.; Campbell, J.L.; et al. Considerations for the Use of Phage Therapy in Clinical Practice. Antimicrob. Agents Chemother. 2022, 66, e0207121. [Google Scholar] [CrossRef]
- Duplessis, C.A.; Biswas, B. A Review of topical phage therapy for chronically infected wounds and preparations for a randomized adaptive clinical trial evaluating topical phage therapy in chronically infected diabetic foot ulcers. Antibiotics 2020, 9, 377. [Google Scholar] [CrossRef]
- Wang, Z.; Kong, L.; Liu, Y.; Fu, Q.; Cui, Z.; Wang, J.; Ma, J.; Wang, H.; Yan, Y.; Sun, J. A Phage Lysin Fused to a Cell-Penetrating Peptide Kills Intracellular Methicillin-Resistant Staphylococcus aureus in Keratinocytes and Has Potential as a Treatment for Skin Infections in Mice. Appl. Environ. Microbiol. 2018, 84, e00380-18. [Google Scholar] [CrossRef] [Green Version]
- Chhibber, S.; Shukla, A.; Kaur, S. Transfersomal Phage Cocktail Is an Effective Treatment against Methicillin-Resistant Staphylococcus aureus -Mediated Skin and Soft Tissue Infections. Antimicrob. Agents Chemother. 2017, 61, e02146-16. [Google Scholar] [CrossRef] [Green Version]
- Jikia, D.; Chkhaidze, N.; Imedashvili, E.; Mgaloblishvili, I.; Tsitlanadze, G.; Katsarava, R.; Glenn Morris, J., Jr.; Sulakvelidze, A. The use of a novel biodegradable preparation capable of the sustained release of bacteriophages and ciprofloxacin, in the complex treatment of multidrug-resistant Staphylococcus aureus -infected local radiation injuries caused by exposure to Sr90. Clin. Exp. Dermatol. 2005, 30, 23–26. [Google Scholar] [CrossRef]
- Plumet, L.; Ahmad-Mansour, N.; Dunyach-Remy, C.; Kissa, K.; Sotto, A.; Lavigne, J.-P.; Costechareyre, D.; Molle, V. Bacteriophage Therapy for Staphylococcus aureus Infections: A Review of Animal Models, Treatments, and Clinical Trials. Front. Cell. Infect. Microbiol. 2022, 12, 907314. [Google Scholar] [CrossRef]
- Medina, E.; Pieper, D.H. Tackling Threats and Future Problems of Multidrug-Resistant Bacteria. Curr. Top. Microbiol. Immunol. 2016, 398, 3–33. [Google Scholar] [CrossRef]
- Duong, M.; Markwell, S.; Peter, J.; Barenkamp, S. Randomized, Controlled Trial of Antibiotics in the Management of Community-Acquired Skin Abscesses in the Pediatric Patient. Ann. Emerg. Med. 2010, 55, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Talan, D.A.; Mower, W.R.; Krishnadasan, A.; Abrahamian, F.M.; Lovecchio, F.; Karras, D.J.; Steele, M.T.; Rothman, R.E.; Hoagland, R.; Moran, G.J. Trimethoprim–Sulfamethoxazole versus Placebo for Uncomplicated Skin Abscess. N. Engl. J. Med. 2016, 374, 823–832. [Google Scholar] [CrossRef] [PubMed] [Green Version]


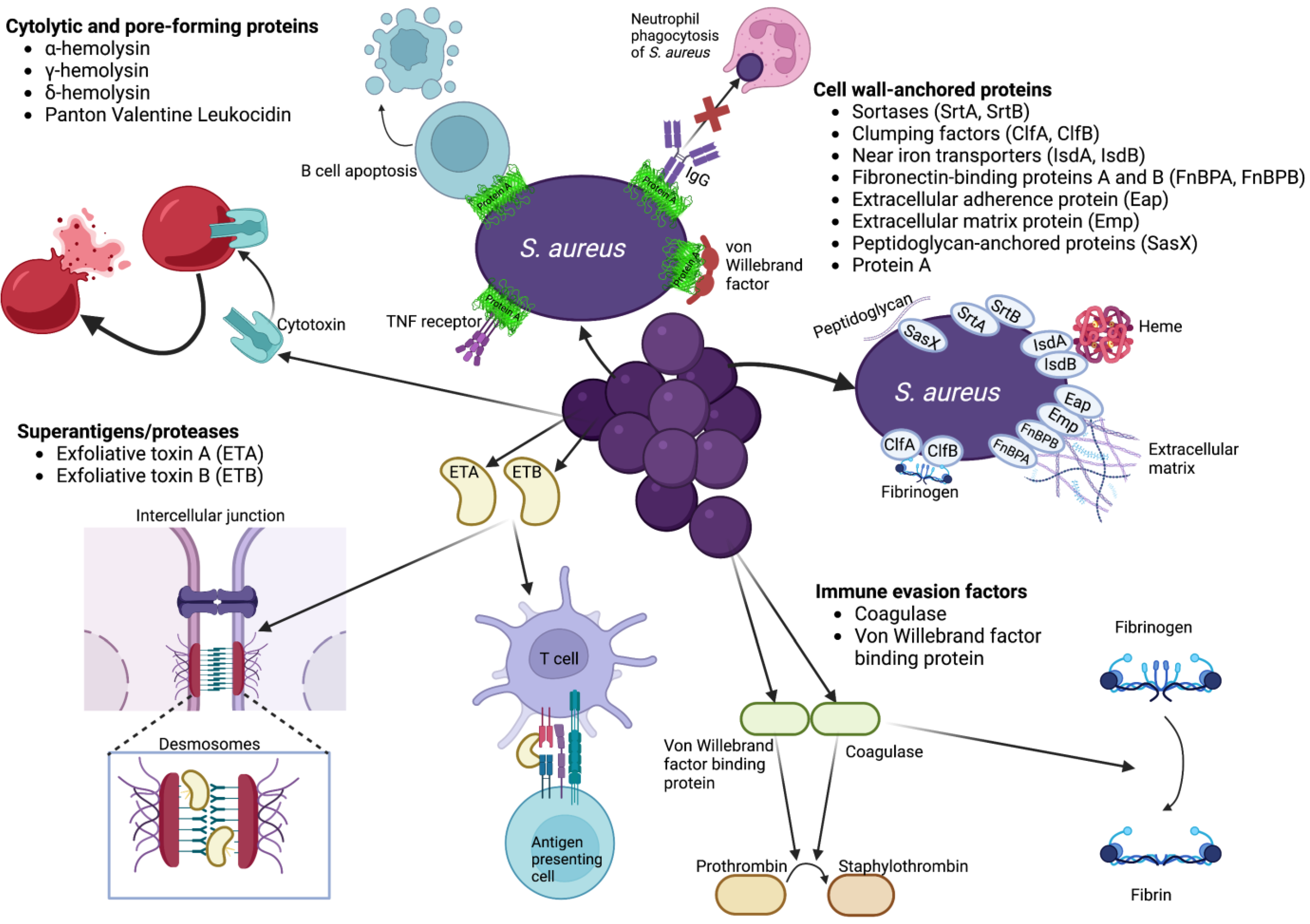{kind=link}
| Virulence Factor | Function | Models | References |
|---|---|---|---|
| Cytolytic and pore-forming proteins | |||
| ɑ-hemolysin (Hla, ɑ-toxin) | General cell lysis | Mouse skin, mastitis | [86,87,88,89,90,91,92] |
| Human HaCaT cells, skin explants | |||
| γ-hemolysin (HlgAB, HlgCB) | Lysis of neutrophils, other leukocytes | Mouse skin | [93,94,95,96] |
| Human skin mimic models | |||
| δ-hemolysin (δ-toxin) | General cell lysis | Mouse skin | [97,98] |
| Mast cell degranulation | |||
| Panton Valentine Leukocidin (PVL) | Lysis of neutrophils, other leukocytes | Mouse skin | [99,100,101,102,103,104,105,106] |
| Rabbit skin | |||
| Monkey skin | |||
| Humanized mice | |||
| Human keratinocytes | |||
| Virulence Factor | Function | Models | References |
|---|---|---|---|
| Superantigens/proteases | |||
| Exfoliative toxins (ETA, ETB) | Crosslink T cell receptor with MHC class II molecules Proteolysis of epidermal desmoglein 1 | Mouse lymphocytes, skin | [107,108,109,110,111] |
| Rabbit intravenous | |||
| Human lymphocytes, skin cryosections | |||
| Immune evasion factors | |||
| Coagulase (Coa) | Activation of prothrombin | Mouse skin, mastitis Rabbit skin | [90,112,113] |
| Proteolytic conversion of fibrinogen to fibrin | |||
| Abscess formation | |||
| Von Willebrand factor binding protein | Activation of prothrombin | Rabbit skin | [90,113] |
| Abscess formation | |||
| Virulence Factor | Function | Models | References |
|---|---|---|---|
| Cell wall-anchored proteins | |||
| Sortases (SrtA, SrtB) | Covalent attachment of surface proteins to cell wall | Mouse skin | [114,115] |
| Abscess formation | |||
| Clumping factors (ClfA, ClfB, SdrC, SdrD, SdrE) | Binding fibrinogen (ClfA, ClfB, SdrE), desquamated epithelial cells (SdrC, SdrD) | Mouse skin, intravenous Rabbit skin | [90,105,114,115,116] |
| Early dissemination (ClfA, ClfB) | |||
| Abscess formation (SdrD) | |||
| Near iron transporters (IsdA, IsdB) | Binding and transporting heme Abscess formation | Mouse skin | [90,105,116,117,118,119] |
| Rabbit skin | |||
| Monkey skin | |||
| Fibronectin-binding proteins A and B (FnBPA, FnBPB) | Adhesion to extracellular matrix | Human corneocytes, human immortal keratinocytes, skin explants, mouse mast cells | [90,114,116,120,121,122,123,124,125] |
| Internalization of bacteria | |||
| Abscess formation | |||
| Extracellular adherence protein (Eap) | Adherence to extracellular matrix | Human HaCaT keratinocytes | [90,116,124,126,127] |
| Internalization by eukaryotic cells | |||
| Biofilm formation | |||
| Abscess formation | |||
| Extracellular matrix protein (Emp) | Binding to extracellular matrix | Human skin | [90,116,127,128] |
| Biofilm formation | |||
| Abscess formation | |||
| Peptidoglycan-anchored proteins (SasX) | Covalent attachment to peptidoglycan | Mouse skin | [129,130,131] |
| Colonization | |||
| Protein A | Binding IgG, TNF receptor 1, von Willebrand factor | Mouse skin Human HaCaT keratinocytes | [90,92,114,132,133,134,135,136,137,138,139,140] |
| Evasion of phagocytosis | |||
| Superantigenic factor for B cell apoptosis | |||
| Pro-inflammatory cytokine expression | |||
| Atopic dermatitis | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Linz, M.S.; Mattappallil, A.; Finkel, D.; Parker, D. Clinical Impact of Staphylococcus aureus Skin and Soft Tissue Infections. Antibiotics 2023, 12, 557. https://doi.org/10.3390/antibiotics12030557
Linz MS, Mattappallil A, Finkel D, Parker D. Clinical Impact of Staphylococcus aureus Skin and Soft Tissue Infections. Antibiotics. 2023; 12(3):557. https://doi.org/10.3390/antibiotics12030557
Chicago/Turabian StyleLinz, Matthew S., Arun Mattappallil, Diana Finkel, and Dane Parker. 2023. "Clinical Impact of Staphylococcus aureus Skin and Soft Tissue Infections" Antibiotics 12, no. 3: 557. https://doi.org/10.3390/antibiotics12030557
APA StyleLinz, M. S., Mattappallil, A., Finkel, D., & Parker, D. (2023). Clinical Impact of Staphylococcus aureus Skin and Soft Tissue Infections. Antibiotics, 12(3), 557. https://doi.org/10.3390/antibiotics12030557