
Phenotypic and Genotypic Investigation of Carbapenem-Resistant Acinetobacter baumannii in Maharaj Nakhon Si Thammarat Hospital, Thailand

 , , and
, , and
Abstract
:1. Introduction
2. Results
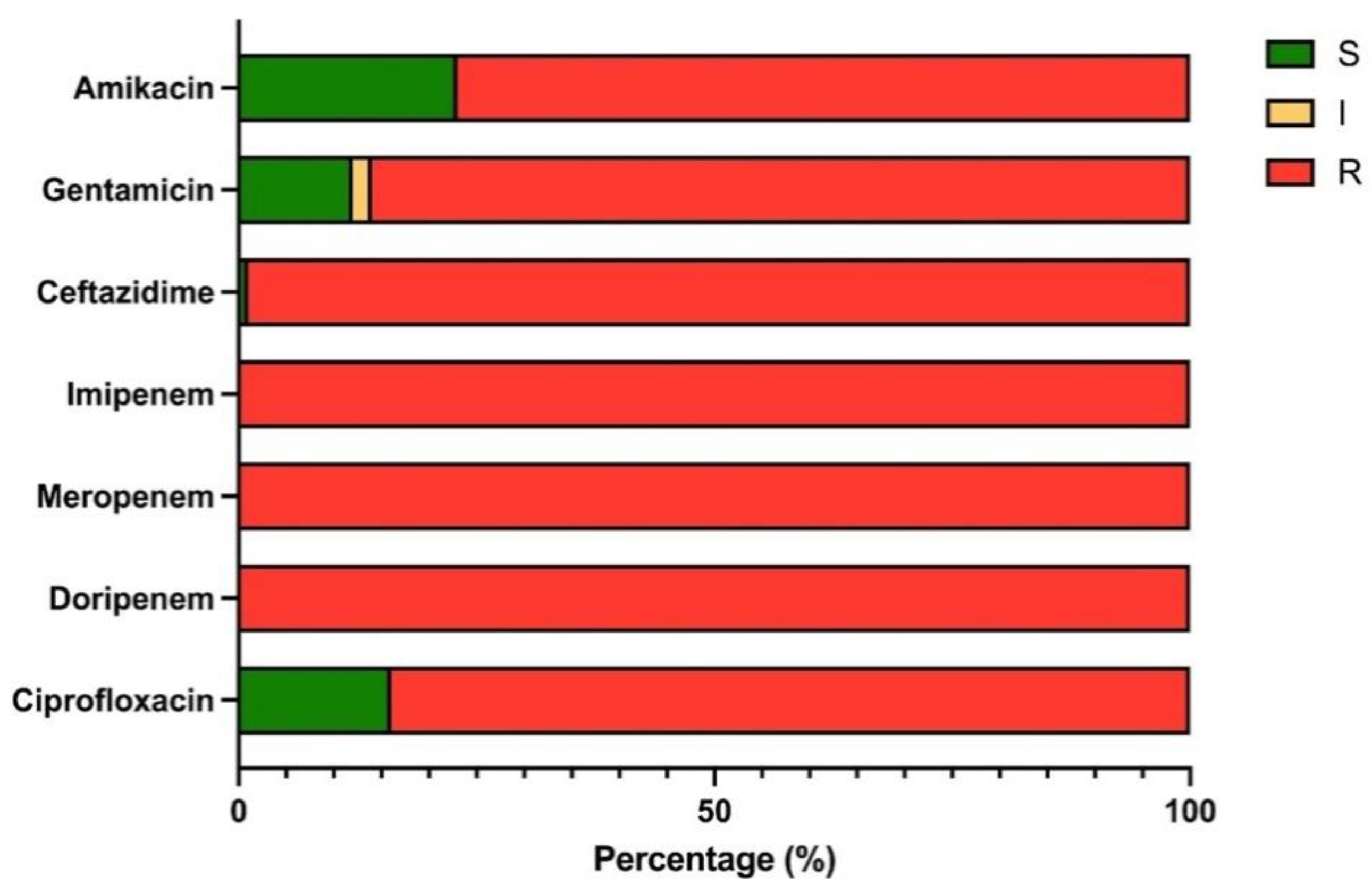
2.1. Distribution of CRAB Isolates and Antimicrobial Profiles
2.2. Hemolysis and Protease Activities
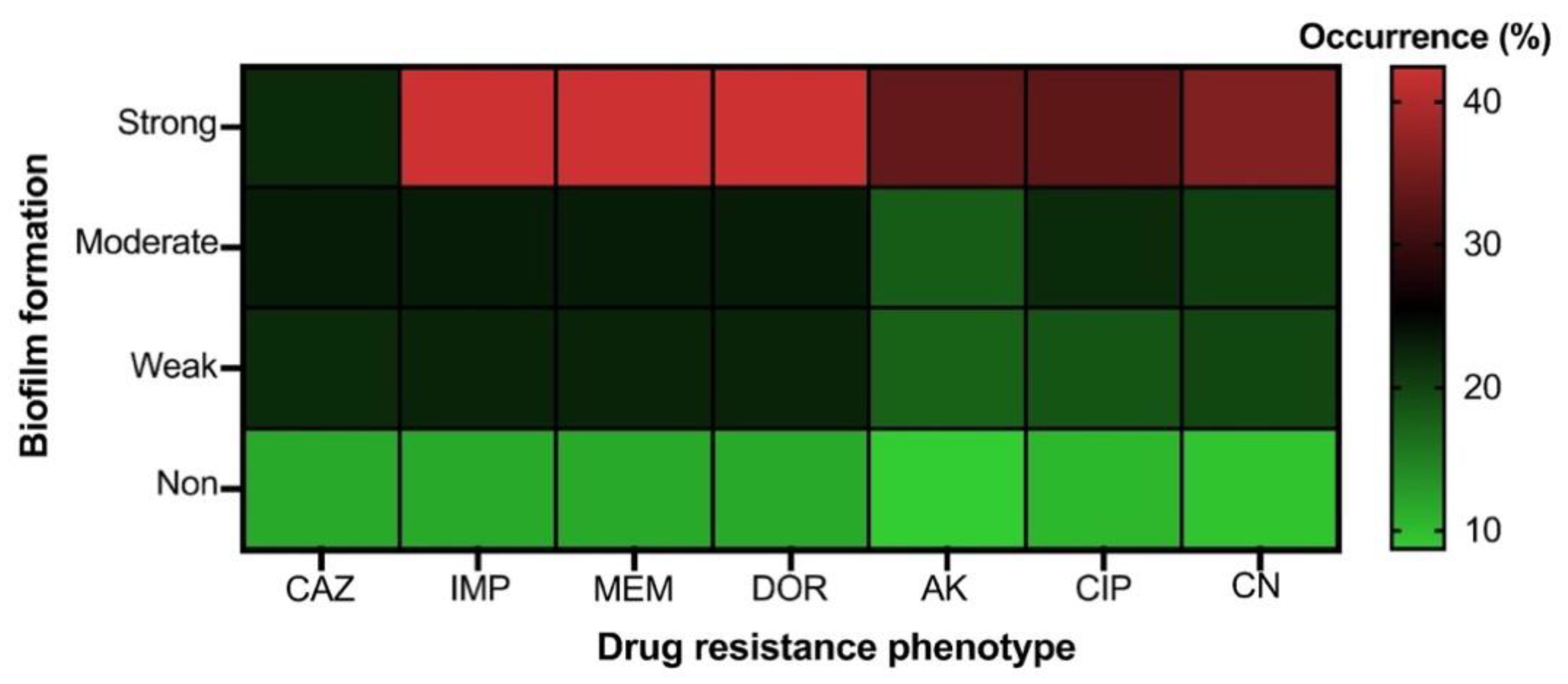
2.3. Biofilm Formation Phenotypes of CRAB
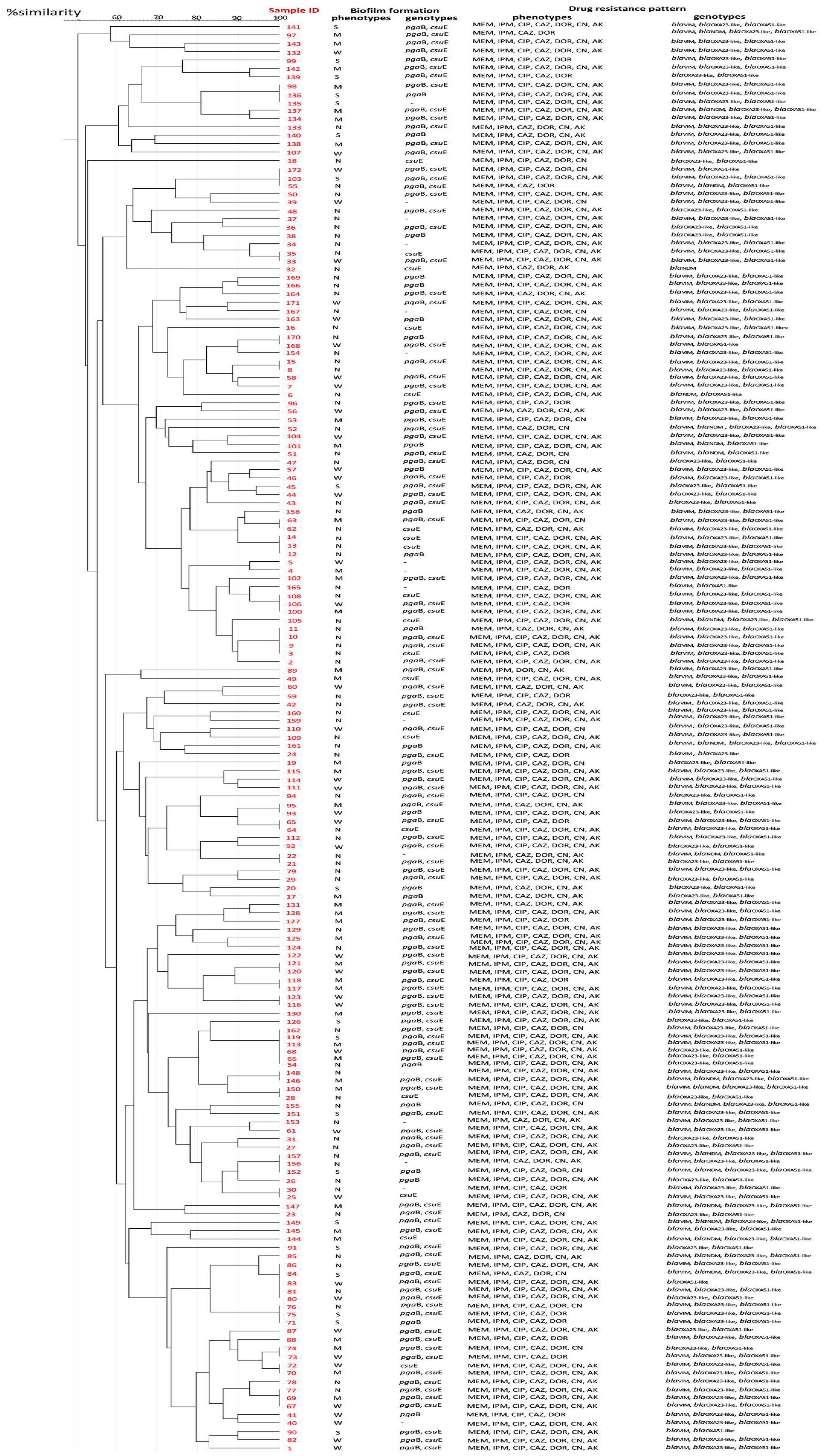
2.4. Prevalence of Drug Resistance, Biofilm-Related Genotypes and Genetic Diversity of CRAB
3. Discussion
4. Materials and Methods
4.1. Study Settings and Ethical Approval
4.2. Bacterial Isolation and Identification
4.3. Antimicrobial Susceptibility Testing
4.4. Hemolysis Assay and Protease Activity
4.5. Biofilm Formation Using Microtiter Plate Assay
4.6. Genotypic Characterisation of Antimicrobial Resistance and Biofilm-Associated Genes by Polymerase Chain Reaction (PCR)
4.7. Molecular Typing and Clonal Relationship between CRAB Strains by Repetitive Element Sequence-Based PCR (REP-PCR)
4.8. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wong, D.; Nielsen, T.B.; Bonomo, R.A.; Pantapalangkoor, P.; Luna, B.; Spellberg, B. Clinical and pathophysiological overview of Acinetobacter infections: A century of challenges. Clin. Microbiol. Rev. 2017, 30, 409–447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahador, A.; Farshadzadeh, Z.; Raoofian, R.; Mokhtaran, M.; Pourakbari, B.; Pourhajibagher, M.; Hashemi, F.B. Association of virulence gene expression with colistin-resistance in Acinetobacter baumannii: Analysis of genotype, antimicrobial susceptibility, and biofilm formation. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 24. [Google Scholar] [CrossRef] [PubMed]
- Zeighami, H.; Valadkhani, F.; Shapouri, R.; Samadi, E.; Haghi, F. Virulence characteristics of multidrug resistant biofilm forming Acinetobacter baumannii isolated from intensive care unit patients. BMC Infect. Dis. 2019, 19, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunenshine, R.H.; Wright, M.O.; Maragakis, L.L.; Harris, A.D.; Song, X.; Hebden, J.; Cosgrove, S.E.; Anderson, A.; Carnell, J.; Jernigan, D.B.; et al. Multidrug-resistant Acinetobacter infection mortality rate and length of hospitalization. Emerg. Infect. Dis. 2007, 13, 97–103. [Google Scholar] [CrossRef]
- Weber, D.J.; Rutala, W.A.; Miller, M.B.; Huslage, K.; Sickbert-Bennett, E. Role of hospital surfaces in the transmission of emerging health care-associated pathogens: Norovirus, Clostridium difficile, and Acinetobacter species. Am. J. Infect. Control 2010, 38, S25–S33. [Google Scholar] [CrossRef]
- World Health Organization. Media Centre. WHO Publishes List of Bacteria for Which New Antibiotics Are Urgently Needed. 2017. Available online: http://www.who.int/mediacentre/news/releases/2017/bacteria-antibiotics-needed/en/ (accessed on 2 December 2021).
- Thummeepak, R.; Kongthai, P.; Leungtongkam, U.; Sitthisak, S. Distribution of virulence genes involved in biofilm formation in multi-drug resistant Acinetobacter baumannii clinical isolates. Int. Microbiol. 2016, 19, 121–129. [Google Scholar]
- Lee, C.R.; Lee, J.H.; Park, M.; Park, K.S.; Bae, I.K.; Kim, Y.B.; Cha, C.J.; Jeong, B.C.; Lee, S.H. Biology of Acinetobacter baumannii: Pathogenesis, antibiotic resistance mechanisms, and prospective treatment options. Front. Cell. Infect. Microbiol. 2017, 7, 55. [Google Scholar] [CrossRef] [Green Version]
- Antunes, N.T.; Lamoureaux, T.L.; Toth, M.; Stewart, N.K.; Frase, H.; Vakulenko, S.B. Class D β-Lactamases: Are They All Carbapenemases? Antimicrob. Agents Chemother. 2014, 58, 2119–2125. [Google Scholar] [CrossRef] [Green Version]
- Kateete, D.P.; Nakanjako, R.; Namugenyi, J.; Erume, J.; Joloba, M.L.; Najjuka, C.F. Carbapenem resistant Pseudomonas aeruginosa and Acinetobacter baumannii at Mulago hospital in Kampala, Uganda (2007–2009). Springerplus 2016, 5, 1308. [Google Scholar] [CrossRef] [Green Version]
- Leski, T.A.; Bangura, U.; Jimmy, D.H.; Ansumana, R.; Lizewski, S.E.; Li, R.W.; Stenger, D.A.; Taitt, C.R.; Vora, G.J. Identification of blaOXA-51-like, blaOXA-58, blaDIM-1, and blaVIM carbapenemase genes in hospital Enterobacteriaceae isolates from Sierra Leone. J. Clin. Microbiol. 2013, 51, 2435–2438. [Google Scholar] [CrossRef] [Green Version]
- Amudhan, M.S.; Sekar, U.; Kamalanathan, A.; Balaraman, S. blaIMP and blaVIM mediated carbapenem resistance in Pseudomonas and Acinetobacter species in India. J. Infect. Dev. Ctries. 2012, 6, 757–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghasemi, E.; Ghalavand, Z.; Goudarzi, H.; Yeganeh, F.; Hashemi, A.; Dabiri, H.; Mirsamadi, E.S.; Foroumand, M. Phenotypic and genotypic investigation of biofilm formation in clinical and environmental isolates of Acinetobacter baumannii. Arch. Clin. Infect. Dis. 2018, 13, e12914. [Google Scholar] [CrossRef] [Green Version]
- Sung, J.Y. Molecular characterization and antimicrobial susceptibility of biofilm-forming Acinetobacter baumannii clinical isolates from Daejeon, Korea. Korean J. Clin. Lab. Sci. 2018, 50, 100–109. [Google Scholar] [CrossRef] [Green Version]
- Li, F.J.; Starrs, L.; Burgio, G. Tug of war between Acinetobacter baumannii and host immune responses. Pathog. Dis. 2018, 76, ftz004. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Chang, Y.; Xu, Y.; Luo, Y.; Wu, L.; Mei, Z.; Li, S.; Wang, R.; Jia, X. Distribution of virulence-associated genes and antimicrobial susceptibility in clinical Acinetobacter baumannii isolates. Oncotarget 2018, 9, 21663–21673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Shamiri, M.M.; Zhang, S.; Mi, P.; Liu, Y.; Xun, M.; Yang, E.; Ai, L.; Han, L.; Chen, Y. Phenotypic and genotypic characteristics of Acinetobacter baumannii enrolled in the relationship among antibiotic resistance, biofilm formation and motility. Microb. Pathog. 2021, 155, 104922. [Google Scholar] [CrossRef] [PubMed]
- Tyumentseva, M.; Mikhaylova, Y.; Prelovskaya, A.; Tyumentsev, A.; Petrova, L.; Fomina, V.; Zamyatin, M.; Shelenkov, A.; Akimkin, V. Genomic and phenotypic analysis of multidrug-resistant Acinetobacter baumannii clinical isolates carrying different types of CRISPR/Cas systems. Pathogens 2021, 10, 205. [Google Scholar] [CrossRef]
- Silva, A.; Costa Junior, S.D.; Lima, J.L.; Farias Filho, J.L.B.; Cavalcanti, I.M.; Maciel, M.A.V. Investigation of the association of virulence genes and biofilm production with infection and bacterial colonization processes in multidrug-resistant Acinetobacter spp. An. Acad. Bras. Cienc. 2021, 93, 1–13. [Google Scholar] [CrossRef]
- Xiong, L.; Huang, X.; Chen, H.; Liu, Z.; Wang, D.; Zeng, G.; Mo, S.; Wei, C. Correlation analysis between drug resistance and virulence genes of carbapenem-resistant Acinetobacter baumannii in Western Guilin. bioRxiv 2022. [Google Scholar] [CrossRef]
- Yang, C.H.; Su, P.W.; Moi, S.H.; Chuang, L.Y. Biofilm formation in Acinetobacter baumannii: Genotype-phenotype correlation. Molecules 2019, 24, 1849. [Google Scholar] [CrossRef] [Green Version]
- Tognim, M.C.; Andrade, S.S.; Silbert, S.; Gales, A.C.; Jones, R.N.; Sader, H.S. Resistance trends of Acinetobacter spp. in Latin America and characterization of international dissemination of multi-drug resistant strains: Five-year report of the SENTRY Antimicrobial Surveillance Program. Int. J. Infect. Dis. 2004, 8, 284–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peleg, A.Y.; Seifert, H.; Paterson, D.L. Acinetobacter baumannii: Emergence of a successful pathogen. Clin. Microbiol. Rev. 2008, 21, 538–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jain, M.; Sharma, A.; Sen, M.K.; Rani, V.; Gaind, R.; Suri, J.C. Phenotypic and molecular characterization of Acinetobacter baumannii isolates causing lower respiratory infections among ICU patients. Microb. Pathog. 2019, 128, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Zarrilli, R.; Crispino, M.; Bagattini, M.; Barretta, E.; Popolo, A.D.; Triassi, M. Molecular epidemiology of sequential outbreaks of Acinetobacter baumannii in an intensive care unit shows the emergence of carbapenem resistance. J. Clin. Microbiol. 2004, 42, 946–953. [Google Scholar] [CrossRef] [Green Version]
- Farivar, A.S.; Nowroozi, J.; Emami, M. The prevalence of Acinetobacter in sergical ICU in Rasoul Akram hospital in 2004–2005. J. Rafsanjan Univ. Med. Sci. Health Serv. 2005, 4, 342–347. [Google Scholar]
- Turton, J.F.; Woodford, N.; Glover, J.; Yarde, S.; Kaufmann, M.E.; Pitt, T.L. Identification of Acinetobacter baumannii by detection of the blaOXA-51-like carbapenemase gene intrinsic to this species. J. Clin. Microbiol. 2006, 44, 2974–2976. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, G.J.; Domingues, S. Insights on the horizontal gene transfer of carbapenemase determinants in the opportunistic pathogen Acinetobacter baumannii. Microorganisms 2016, 4, 29. [Google Scholar] [CrossRef] [Green Version]
- Akbari, M.; Niakan, M.; Taherikalani, M.; Feizabadi, M.M.; Azadi, N.A.; Soroush, S.; Emaneini, M.; Abdolkarimi, A.; Maleki, A.; Hematian, A. Rapid identification of Iranian Acinetobacter baumannii strains by single PCR assay using BLAoxa-51-like carbapenemase and evaluation of the antimicrobial resistance profiles of the isolates. Acta Microbiol. Immunol. Hung. 2010, 57, 87–94. [Google Scholar]
- Nigro, S.J.; Hall, R.M. Does the intrinsic oxaAb (blaOXA-51-like) gene of Acinetobacter baumannii confer resistance to carbapenems when activated by ISAba1? J. Antimicrob. Chemother. 2018, 73, 3518–3520. [Google Scholar] [CrossRef]
- Al-Hassan, L.; El Mehallawy, H.; Amyes, S.G. Diversity in Acinetobacter baumannii isolates from paediatric cancer patients in Egypt. Clin. Microbiol. Infect. 2013, 19, 1082–1088. [Google Scholar] [CrossRef] [Green Version]
- Fouad, M.; Attia, A.S.; Tawakkol, W.M.; Hashem, A.M. Emergence of carbapenem-resistant Acinetobacter baumannii harboring the OXA-23 carbapenemase in intensive care units of Egyptian hospitals. Int. J. Infect. Dis. 2013, 17, e1252–e1254. [Google Scholar] [CrossRef] [Green Version]
- Al-Agamy, M.H.; Khalaf, N.G.; Tawfick, M.M.; Shibl, A.M.; El Kholy, A. Molecular characterization of carbapenem-insensitive Acinetobacter baumannii in Egypt. Int. J. Infect. Dis. 2014, 22, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Todorova, B.; Velinov, T.; Ivanov, I.; Dobreva, E.; Kantardjiev, T. First detection of OXA-24 carbapenemase-producing Acinetobacter baumannii isolates in Bulgaria. World J. Microbiol. Biotechnol. 2014, 30, 1427–1430. [Google Scholar] [CrossRef] [PubMed]
- Leungtongkam, U.; Thummeepak, R.; Wongprachan, S.; Thongsuk, P.; Kitti, T.; Ketwong, K.; Runcharoen, C.; Chantratita, N.; Sitthisak, S. Dissemination of blaOXA-23, blaOXA-24, blaOXA-58, and blaNDM-1 genes of Acinetobacter baumannii isolates from four tertiary hospitals in Thailand. Microb. Drug Resist. 2018, 24, 55–62. [Google Scholar] [CrossRef]
- Le Hello, S.; Falcot, V.; Lacassin, F.; Baumann, F.; Nordmann, P.; Naas, T. Molecular epidemiology of carbapenem-resistant Acinetobacter baumannii in New Caledonia. Clin. Microbiol. Infect. 2008, 14, 977–981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, F.; Quinteira, S.; Poirel, L.; Novais, Â.; Peixe, L. Role of common blaOXA-24/OXA-40-carrying platforms and plasmids in the spread of OXA-24/OXA-40 among Acinetobacter species clinical isolates. Antimicrob. Agents Chemother. 2012, 56, 3969–3972. [Google Scholar] [CrossRef] [Green Version]
- Mussi, M.A.; Relling, V.M.; Limansky, A.S.; Viale, A.M. CarO, an Acinetobacter baumannii outer membrane protein involved in carbapenem resistance, is essential for L-ornithine uptake. FEBS Lett. 2007, 581, 5573–5578. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.Y.; Lee, H.J.; Suh, J.T.; Lee, K.M. Outbreaks of imipenem resistant Acinetobacter baumannii producing OXA-23 β-lactamase in a tertiary care hospital in Korea. Yonsei Med. J. 2009, 50, 764–770. [Google Scholar] [CrossRef] [Green Version]
- Almasaudi, S.B. Acinetobacter spp. as nosocomial pathogens: Epidemiology and resistance features. Saudi J. Biol. Sci. 2018, 25, 586–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Connell, M.J.; Actis, L.; Pachón, J. Acinetobacter baumannii: Human infections, factors contributing to pathogenesis and animal models. FEMS Microbiol. Rev. 2013, 37, 130–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerdes, K.; Christensen, S.K.; Løbner-Olesen, A. Prokaryotic toxin–antitoxin stress response loci. Nat. Rev. Microbiol. 2005, 3, 371–382. [Google Scholar] [CrossRef]
- Manchanda, V.; Sanchaita, S.; Singh, N. Multidrug resistant acinetobacter. J. Glob. Infect. Dis. 2010, 2, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Misba, L.; Khan, A.U. Antibiotics versus biofilm: An emerging battleground in microbial communities. Antimicrob. Resist. Infect. Control 2019, 8, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espinal, P.; Marti, S.; Vila, J. Effect of biofilm formation on the survival of Acinetobacter baumannii on dry surfaces. J. Hosp. Infect. 2012, 80, 56–60. [Google Scholar] [CrossRef]
- Roca, I.; Espinal, P.; Vila-Farres, X.; Vila, J. The Acinetobacter baumannii Oxymoron: Commensal hospital dweller turned pan-drug-resistant menace. Front. Microbiol. 2012, 3, 148. [Google Scholar] [CrossRef] [Green Version]
- Thom, K.A.; Johnson, J.K.; Lee, M.S.; Harris, A.D. Environmental contamination because of multidrug-resistant Acinetobacter baumannii surrounding colonized or infected patients. Am. J. Infect. Control 2011, 39, 711–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossard, K.A.; Campagnari, A.A. The Acinetobacter baumannii biofilm-associated protein plays a role in adherence to human epithelial cells. Infect. Immun. 2012, 80, 228–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longo, F.; Vuotto, C.; Donelli, G. Biofilm formation in Acinetobacter baumannii. New Microbiol. 2014, 37, 119–127. [Google Scholar] [PubMed]
- Choi, A.H.; Slamti, L.; Avci, F.Y.; Pier, G.B.; Maira-Litrán, T. The pgaABCD locus of Acinetobacter baumannii encodes the production of poly-β-1-6-N-acetylglucosamine, which is critical for biofilm formation. J. Bacteriol. 2009, 191, 5953–5963. [Google Scholar] [CrossRef] [Green Version]
- Tomaras, A.P.; Flagler, M.J.; Dorsey, C.W.; Gaddy, J.A.; Actis, L.A. Characterization of a two-component regulatory system from Acinetobacter baumannii that controls biofilm formation and cellular morphology. Microbiology 2008, 154, 3398–3409. [Google Scholar] [CrossRef] [Green Version]
- Goh, H.S.; Beatson, S.A.; Totsika, M.; Moriel, D.G.; Phan, M.D.; Szubert, J.; Runnegar, N.; Sidjabat, H.E.; Paterson, D.L.; Nimmo, G.R.; et al. Molecular analysis of the Acinetobacter baumannii biofilm-associated protein. Appl. Environ. Microbiol. 2013, 79, 6535–6543. [Google Scholar] [CrossRef] [Green Version]
- Dahdouh, E.; Hajjar, M.; Suarez, M.; Daoud, Z. Acinetobacter baumannii isolated from Lebanese patients: Phenotypes and genotypes of resistance, clonality, and determinants of pathogenicity. Front. Cell. Infect. Microbiol. 2016, 6, 163. [Google Scholar] [CrossRef] [Green Version]
- Sabat, A.J.; Budimir, A.; Nashev, D.; Sa-Leao, R.; van Dijl, J.; Laurent, F.; Grundmann, H.; Friedrich, A.W. ESCMID study group of epidemiological markers (ESGEM). Overview of molecular typing methods for outbreak detection and epidemiological surveillance. Eurosurveillance 2013, 18, 20380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grisold, A.J.; Zarfel, G.; Strenger, V.; Feierl, G.; Leitner, E.; Masoud, L.; Hoenigl, M.; Raggam, R.B.; Dosch, V.; Marth, E. Use of automated repetitive-sequence-based PCR for rapid laboratory confirmation of nosocomial outbreaks. J. Infect. 2010, 60, 44–51. [Google Scholar] [CrossRef] [PubMed]
- Pasanen, T.; Koskela, S.; Mero, S.; Tarkka, E.; Tissari, P.; Vaara, M.; Kirveskari, J. Rapid molecular characterization of Acinetobacter baumannii clones with rep-PCR and evaluation of carbapenemase genes by new multiplex PCR in Hospital District of Helsinki and Uusimaa. PLoS ONE 2014, 9, e85854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bou, G.; Cervero, G.; Dominguez, M.A.; Quereda, C.; Martinez-Beltran, J. PCR-based DNA fingerprinting (REP-PCR, AP-PCR) and pulsed-field gel electrophoresis characterization of a nosocomial outbreak caused by imipenem- and meropenem-resistant Acinetobacter baumannii. Clin. Microbiol. Infect. 2000, 6, 635–643. [Google Scholar] [CrossRef] [Green Version]
- Ghajavand, H.; Esfahani, B.N.; Havaei, S.A.; Moghim, S.; Fazeli, H. Molecular identification of Acinetobacter baumannii isolated from intensive care units and their antimicrobial resistance patterns. Adv. Biomed. Res. 2015, 4, 110. [Google Scholar] [PubMed]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Document M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020; pp. 46–49. [Google Scholar]
- Dent, L.L.; Marshall, D.R.; Pratap, S.; Hulette, R.B. Multidrug resistant Acinetobacter baumannii: A descriptive study in a city hospital. BMC Infect. Dis. 2010, 10, 196. [Google Scholar] [CrossRef] [Green Version]
- Castellanos, N.; Nakanouchi, J.; Yüzen, D.I.; Fung, S.; Fernandez, J.S.; Barberis, C.; Tuchscherr, L.; Ramirez, M.S. A study on Acinetobacter baumannii and Staphylococcus aureus strains recovered from the same infection site of a diabetic patient. Curr. Microbiol. 2019, 76, 842–847. [Google Scholar] [CrossRef]
- Vijayakumar, S.; Rajenderan, S.; Laishram, S.; Anandan, S.; Balaji, V.; Biswas, I. Biofilm formation and motility depend on the nature of the Acinetobacter baumannii clinical isolates. Front. Public. Health. 2016, 4, 105. [Google Scholar] [CrossRef] [PubMed]
- Shali, A.A.; Jalal, P.J.; Arif, S.K. Dissemination and Genetic Relatedness of Multidrug-Resistant and Extensively Drug-Resistant Acinetobacter baumannii Isolates from a Burn Hospital in Iraq. Can. J. Infect. Dis. Med. Microbiol. 2022, 2022, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Selim, S.; Faried, O.A.; Almuhayawi, M.S.; Mohammed, O.A.; Saleh, F.M.; Warrad, M. Dynamic gene clusters mediating carbapenem-resistant Acinetobacter baumannii clinical isolates. Antibiotics 2022, 11, 168. [Google Scholar] [CrossRef] [PubMed]
- Maleki, A.; Kaviar, V.H.; Koupaei, M.; Haddadi, M.H.; Kalani, B.S.; Valadbeigi, H.; Karamolahi, S.; Omidi, N.; Hashemian, M.; Sadeghifard, N.; et al. Molecular typing and antibiotic resistance patterns among clinical isolates of Acinetobacter baumannii recovered from burn patients in Tehran, Iran. Front. Microbiol. 2022, 13, 994303. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Antimicrobial Agents | Antimicrobial Susceptibility Profile of CRAB Isolates (n = 172) | ||
|---|---|---|---|
| Susceptible n (%) | Intermediate n (%) | Resistance n (%) | |
| Group Cephalosporins | |||
| ceftazidime (CAZ) | 1 (0.58) | 0 (0) | 171 (99.42) |
| Group Aminoglycosides | |||
| gentamicin (CN) | 21 (12.21) | 4 (2.33) | 147 (85.47) |
| amikacin (AK) | 40 (23.26) | 0 (0) | 132 (76.74) |
| Group Carbapenem | |||
| meropenem (MEM) | 0 (0) | 0 (0) | 172 (100) |
| imipenem (IPM) | 0 (0) | 0 (0) | 172 (100) |
| doripenem (DOR) | 0 (0) | 0 (0) | 172 (100) |
| Group Fluoroquinolones | |||
| ciprofloxacin (CIP) | 27 (15.70) | 0 (0) | 145 (84.30) |
| Antibiotics | Antibiotic Resistance | Antibiotic Susceptible | p-Value | ||
|---|---|---|---|---|---|
| Biofilm Producers Isolates (%) | Non-Biofilm Producers Isolates (%) | Biofilm Producers Isolates (%) | Non-Biofilm Producers Isolates (%) | ||
| Amikacin | 73 (55.30%) | 59 (44.70%) | 26 (65%) | 14 (35%) | 0.277 |
| Gentamicin | 85 (57.82%) | 62 (42.18%) | 14 (66.67%) | 7 (33.33%) | 0.441 |
| Ceftazidime | 73 (42.69%) | 98 (57.31%) | 1 (100%) | 0 (0%) | 0.248 |
| Imipenem/Meropenem/Doripenem | 99 (57.56%) | 73 (42.44%) | 0 (0%) | 0 (0%) | NT |
| Ciprofloxacin | 88 (60.69%) | 57 (39.31%) | 12(44.44%) | 15(55.56%) | 0.166 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santajit, S.; Bhoopong, P.; Kong-Ngoen, T.; Tunyong, W.; Horpet, D.; Paehoh-ele, W.; Zahedeng, T.; Pumirat, P.; Sookrung, N.; Hinthong, W.; et al. Phenotypic and Genotypic Investigation of Carbapenem-Resistant Acinetobacter baumannii in Maharaj Nakhon Si Thammarat Hospital, Thailand. Antibiotics 2023, 12, 580. https://doi.org/10.3390/antibiotics12030580
Santajit S, Bhoopong P, Kong-Ngoen T, Tunyong W, Horpet D, Paehoh-ele W, Zahedeng T, Pumirat P, Sookrung N, Hinthong W, et al. Phenotypic and Genotypic Investigation of Carbapenem-Resistant Acinetobacter baumannii in Maharaj Nakhon Si Thammarat Hospital, Thailand. Antibiotics. 2023; 12(3):580. https://doi.org/10.3390/antibiotics12030580
Chicago/Turabian StyleSantajit, Sirijan, Phuangthip Bhoopong, Thida Kong-Ngoen, Witawat Tunyong, Dararat Horpet, Wanfudhla Paehoh-ele, Tasneem Zahedeng, Pornpan Pumirat, Nitat Sookrung, Woranich Hinthong, and et al. 2023. "Phenotypic and Genotypic Investigation of Carbapenem-Resistant Acinetobacter baumannii in Maharaj Nakhon Si Thammarat Hospital, Thailand" Antibiotics 12, no. 3: 580. https://doi.org/10.3390/antibiotics12030580
APA StyleSantajit, S., Bhoopong, P., Kong-Ngoen, T., Tunyong, W., Horpet, D., Paehoh-ele, W., Zahedeng, T., Pumirat, P., Sookrung, N., Hinthong, W., & Indrawattana, N. (2023). Phenotypic and Genotypic Investigation of Carbapenem-Resistant Acinetobacter baumannii in Maharaj Nakhon Si Thammarat Hospital, Thailand. Antibiotics, 12(3), 580. https://doi.org/10.3390/antibiotics12030580