

Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
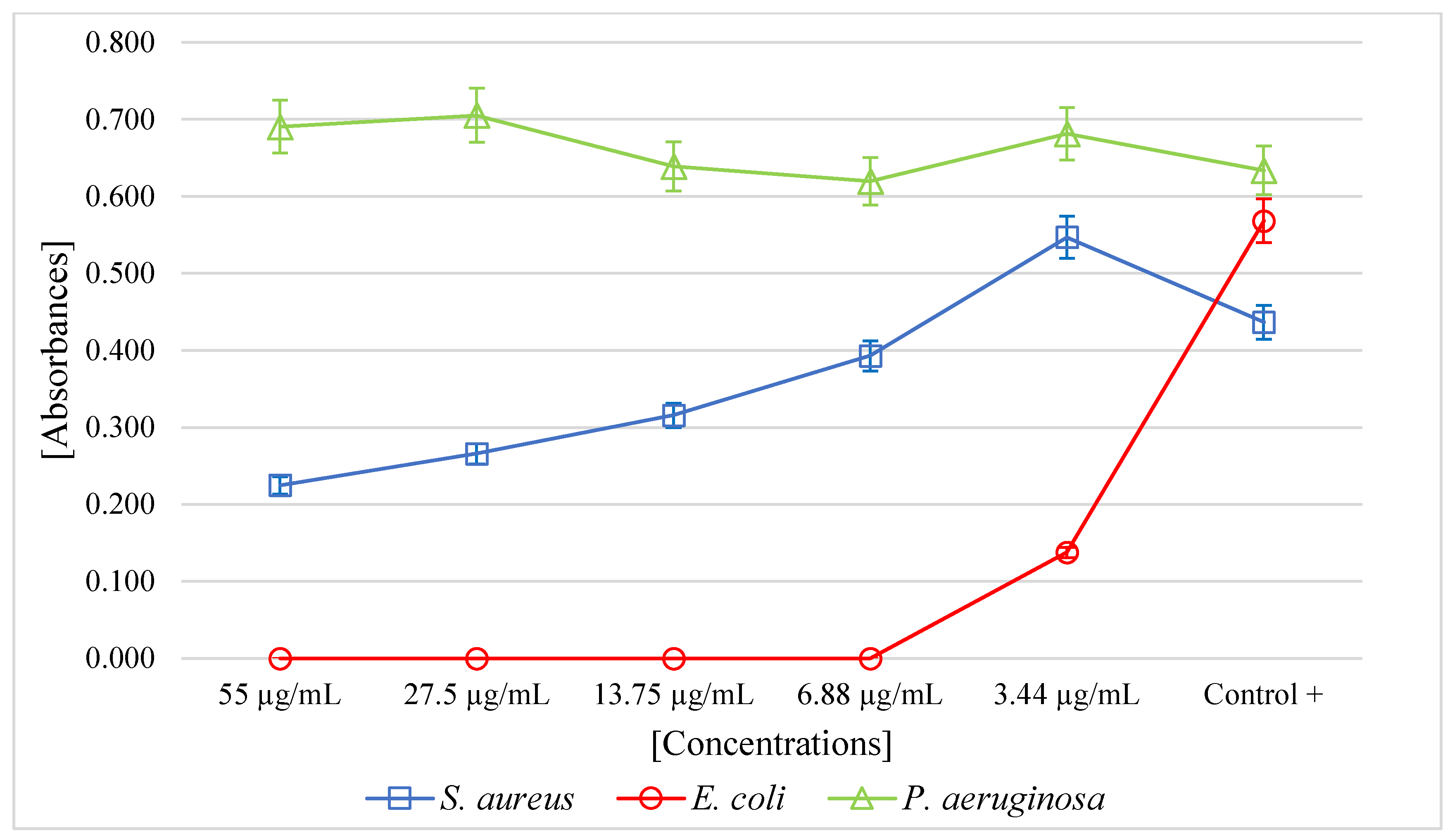
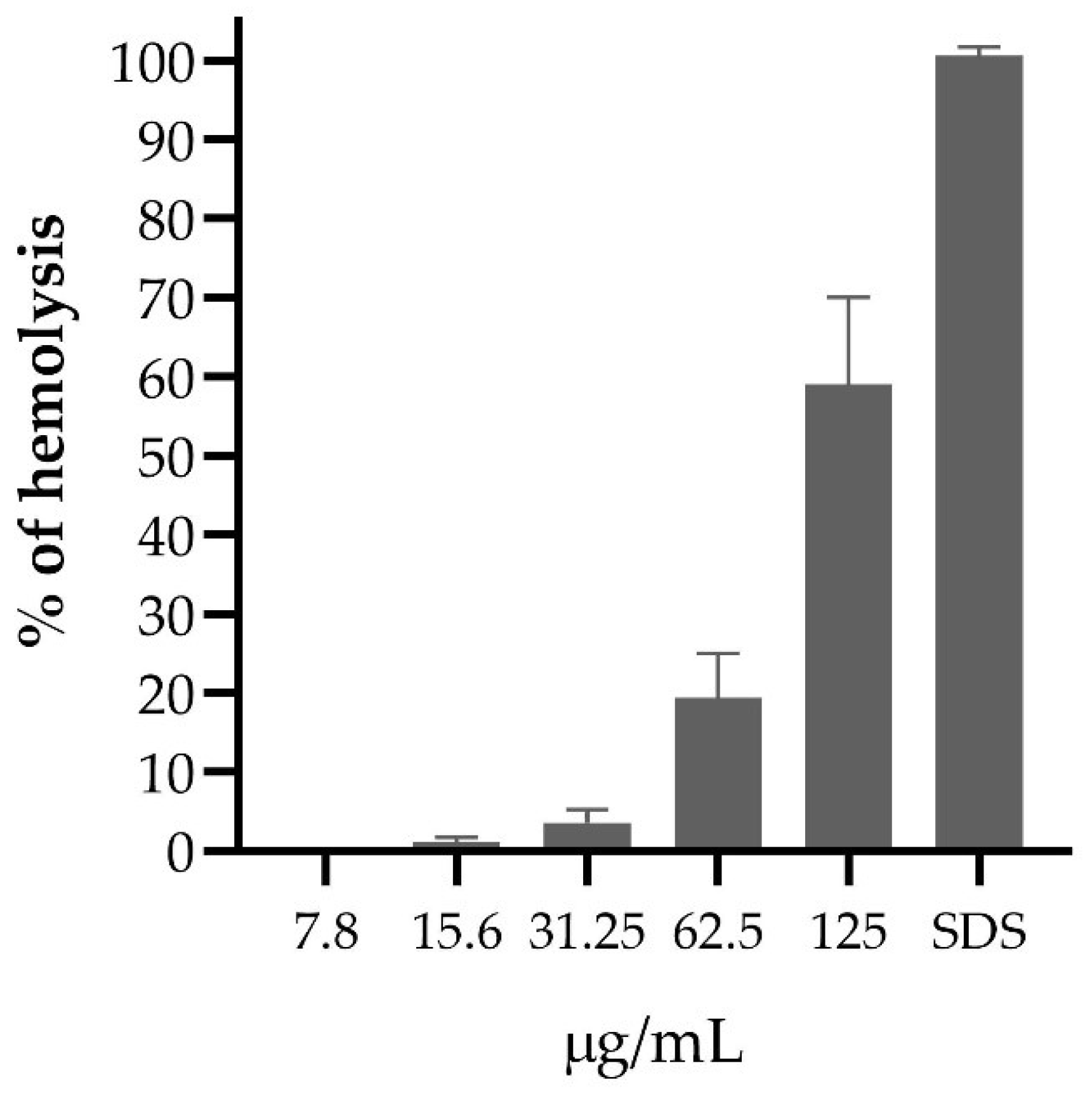
2. Results
3. Discussion
4. Materials and Methods
4.1. Bee Venom Samples
4.2. Fraction Concentration and Electrophoresis
4.3. Antimicrobial Activity Test
4.4. Evaluation of Hemolytic Activity
4.5. Evaluation of Antioxidant Activity
4.6. Statistic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Antimicrobial Resistance: Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; ISBN 978-92-4-156474-8. [Google Scholar]
- Pollitt, E.J.G.; Szkuta, P.T.; Burns, N.; Foster, S.J. Staphylococcus Aureus Infection Dynamics. PLoS Pathog. 2018, 14, e1007112. [Google Scholar] [CrossRef]
- Tong, S.Y.C.; Davis, J.S.; Eichenberger, E.; Holland, T.L.; Fowler, V.G. Staphylococcus Aureus Infections: Epidemiology, Pathophysiology, Clinical Manifestations, and Management. Clin. Microbiol. Rev. 2015, 28, 603–661. [Google Scholar] [CrossRef]
- Safdari, H.; Aryan, E.; Sadeghian, H.; Shams, S.F.; Aganj, M. Frequency of Methicillin-Resistant Staphylococcus Aureus (MRSA) in Nose and Cellular Phone of Medical and Non-Medical Personnel of Emergency Departments of Ghaem Hospital in Mashhad City. Clin. Epidemiol. Glob. Health 2020, 8, 1043–1046. [Google Scholar] [CrossRef]
- Belayhun, C.; Tilahun, M.; Seid, A.; Shibabaw, A.; Sharew, B.; Belete, M.A.; Demsiss, W. Asymptomatic Nasopharyngeal Bacterial Carriage, Multi-Drug Resistance Pattern and Associated Factors among Primary School Children at Debre Berhan Town, North Shewa, Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2023, 22, 9. [Google Scholar] [CrossRef]
- Akanbi, O.E.; Njom, H.A.; Fri, J.; Otigbu, A.C.; Clarke, A.M. Antimicrobial Susceptibility of Staphylococcus Aureus Isolated from Recreational Waters and Beach Sand in Eastern Cape Province of South Africa. Int. J. Environ. Res. Public Health 2017, 14, 1001. [Google Scholar] [CrossRef]
- Qodrati, M.; SeyedAlinaghi, S.; Dehghan Manshadi, S.A.; Abdollahi, A.; Dadras, O. Antimicrobial Susceptibility Testing of Staphylococcus Aureus Isolates from Patients at a Tertiary Hospital in Tehran, Iran, 2018–2019. Eur. J. Med. Res. 2022, 27, 152. [Google Scholar] [CrossRef]
- Nikbakht, M.; Ahangarzadeh Rezaee, M.; Hasani, A.; Nahaei, M.R.; Sadeghi, J.; Jedari Seifi, S. Molecular Characterization and Antimicrobial Susceptibility Patterns of Methicillin-Resistant Staphylococcus Aureus Isolates in Tabriz, Northwest of Iran. Arch. Pediatr. Infect. Dis. 2017, in press. [CrossRef]
- Guo, Y.; Song, G.; Sun, M.; Wang, J.; Wang, Y. Prevalence and Therapies of Antibiotic-Resistance in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2020, 10, 107. [Google Scholar] [CrossRef]
- Hu, P.; Chen, J.; Chen, Y.; Zhou, T.; Xu, X.; Pei, X. Molecular Epidemiology, Resistance, and Virulence Properties of Pseudomonas aeruginosa Cross-Colonization Clonal Isolates in the Non-Outbreak Setting. Infect. Genet. Evol. 2017, 55, 288–296. [Google Scholar] [CrossRef]
- Nasser, M.; Gayen, S.; Kharat, A.S. Prevalence of β-Lactamase and Antibiotic-Resistant Pseudomonas aeruginosa in the Arab Region. J. Glob. Antimicrob. Resist. 2020, 22, 152–160. [Google Scholar] [CrossRef]
- Hoque, M.N.; Jahan, M.I.; Hossain, M.A.; Sultana, M. Genomic Diversity and Molecular Epidemiology of a Multidrug-Resistant Pseudomonas aeruginosa DMC30b Isolated from a Hospitalized Burn Patient in Bangladesh. J. Glob. Antimicrob. Resist. 2022, 31, 110–118. [Google Scholar] [CrossRef]
- Coșeriu, R.L.; Vintilă, C.; Mare, A.D.; Ciurea, C.N.; Togănel, R.O.; Cighir, A.; Simion, A.; Man, A. Epidemiology, Evolution of Antimicrobial Profile and Genomic Fingerprints of Pseudomonas aeruginosa before and during COVID-19: Transition from Resistance to Susceptibility. Life 2022, 12, 2049. [Google Scholar] [CrossRef]
- Asadpour, L. Antimicrobial Resistance, Biofilm-Forming Ability and Virulence Potential of Pseudomonas aeruginosa Isolated from Burn Patients in Northern Iran. J. Glob. Antimicrob. Resist. 2018, 13, 214–220. [Google Scholar] [CrossRef]
- Schiavano, G.F.; Carloni, E.; Andreoni, F.; Magi, S.; Chironna, M.; Brandi, G.; Amagliani, G. Prevalence and Antibiotic Resistance of Pseudomonas aeruginosa in Water Samples in Central Italy and Molecular Characterization of OprD in Imipenem Resistant Isolates. PLoS ONE 2017, 12, e0189172. [Google Scholar] [CrossRef]
- Alotaibi, B.S.; Tantry, B.A.; Farhana, A.; Alammar, M.A.; Shah, N.N.; Mohammed, A.H.; Wani, F.; Bandy, A. Resistance Pattern in Mostly Gram-Negative Bacteria Causing Urinary Tract Infections. Infect. Disord. Drug Targets 2023, 23, 56–64. [Google Scholar] [CrossRef]
- Umpiérrez, A.; Ernst, D.; Fernández, M.; Oliver, M.; Casaux, M.L.; Caffarena, R.D.; Schild, C.; Giannitti, F.; Fraga, M.; Zunino, P. Virulence Genes of Escherichia coli in Diarrheic and Healthy Calves. Rev. Argent. Microbiol. 2021, 53, 34–38. [Google Scholar] [CrossRef]
- Sarjana Safain, K.; Bhuyan, G.S.; Hassan Hasib, S.; Islam, M.S.; Mahmud-Un-Nabi, M.A.; Sultana, R.; Tasnim, S.; Noor, F.A.; Sarker, S.K.; Islam, M.T.; et al. Genotypic and Phenotypic Profiles of Antibiotic-resistant Bacteria Isolated from Hospitalised Patients in Bangladesh. Trop. Med. Int. Health 2021, 26, 720–729. [Google Scholar] [CrossRef]
- Jang, J.; Hur, H.-G.; Sadowsky, M.J.; Byappanahalli, M.N.; Yan, T.; Ishii, S. Environmental Escherichia coli.: Ecology and Public Health Implications—A Review. J. Appl. Microbiol. 2017, 123, 570–581. [Google Scholar] [CrossRef]
- Edmond, T.; Yehouenou, L.C.; Malick, Z.F.; Arsene, K.A.; Rene, K.K.; Diouara, A.A.M.; Tonde, I.; Bankole, H.S.; Wilfried, B.K.; Marius, E.A.; et al. Antimicrobial Susceptibility of Community Acquired Escherichia coli in Urinary Tract Infections (UTI) in Benin for Eleven Years (2005–2015). Am. J. Infect. Dis. 2017, 13, 21–27. [Google Scholar] [CrossRef]
- Grados, M.C.; Thuissard, I.J.; Alós, J.-I. Stratification by Demographic and Clinical Data of the Antibiotic Susceptibility of Escherichia Coli from Urinary Tract Infections of the Community. Atención Primaria 2019, 51, 494–498. [Google Scholar] [CrossRef]
- Amanati, A.; Sajedianfard, S.; Khajeh, S.; Ghasempour, S.; Mehrangiz, S.; Nematolahi, S.; Shahhosein, Z. Bloodstream Infections in Adult Patients with Malignancy, Epidemiology, Microbiology, and Risk Factors Associated with Mortality and Multi-Drug Resistance. BMC Infect. Dis. 2021, 21, 636. [Google Scholar] [CrossRef]
- Mahmoudi, S.; Mahzari, M.; Banar, M.; Pourakbari, B.; Haghi Ashtiani, M.T.; Mohammadi, M.; Keshavarz Valian, S.; Mamishi, S. Antimicrobial Resistance Patterns of Gram-Negative Bacteria Isolated from Bloodstream Infections in an Iranian Referral Paediatric Hospital: A 5.5-Year Study. J. Glob. Antimicrob. Resist. 2017, 11, 17–22. [Google Scholar] [CrossRef]
- Shi, N.; Kang, J.; Wang, S.; Song, Y.; Yin, D.; Li, X.; Guo, Q.; Duan, J.; Zhang, S. Bacteriological Profile and Antimicrobial Susceptibility Patterns of Gram-Negative Bloodstream Infection and Risk Factors Associated with Mortality and Drug Resistance: A Retrospective Study from Shanxi, China. Infect. Drug Resist. 2022, 15, 3561–3578. [Google Scholar] [CrossRef]
- Esposito, S.; Maglietta, G.; Di Costanzo, M.; Ceccoli, M.; Vergine, G.; La Scola, C.; Malaventura, C.; Falcioni, A.; Iacono, A.; Crisafi, A.; et al. Retrospective 8-Year Study on the Antibiotic Resistance of Uropathogens in Children Hospitalised for Urinary Tract Infection in the Emilia-Romagna Region, Italy. Antibiotics 2021, 10, 1207. [Google Scholar] [CrossRef]
- Boschetti, G.; Sgarabotto, D.; Meloni, M.; Bruseghin, M.; Whisstock, C.; Marin, M.; Ninkovic, S.; Pinfi, M.; Brocco, E. Antimicrobial Resistance Patterns in Diabetic Foot Infections, an Epidemiological Study in Northeastern Italy. Antibiotics 2021, 10, 1241. [Google Scholar] [CrossRef]
- Obakiro, S.B.; Kiyimba, K.; Paasi, G.; Napyo, A.; Anthierens, S.; Waako, P.; Royen, P.V.; Iramiot, J.S.; Goossens, H.; Kostyanev, T. Prevalence of Antibiotic-Resistant Bacteria among Patients in Two Tertiary Hospitals in Eastern Uganda. J. Glob. Antimicrob. Resist. 2021, 25, 82–86. [Google Scholar] [CrossRef]
- Mahlapuu, M.; Håkansson, J.; Ringstad, L.; Björn, C. Antimicrobial Peptides: An Emerging Category of Therapeutic Agents. Front. Cell. Infect. Microbiol. 2016, 6, 194. [Google Scholar] [CrossRef]
- Tawfik, M.M.; Bertelsen, M.; Abdel-Rahman, M.A.; Strong, P.N.; Miller, K. Scorpion Venom Antimicrobial Peptides Induce Siderophore Biosynthesis and Oxidative Stress Responses in Escherichia coli. mSphere 2021, 6, e00267-21. [Google Scholar] [CrossRef]
- Pérez-Delgado, O.; Rincon-Cortés, C.A.; Vega-Castro, N.A.; Reyes-Montaño, E.A.; Gómez-Garzón, M. Purificación Parcial de Péptidos Del Veneno de Escorpión Hadruroides Charcasus (Karsch, 1879) Con Actividad Antimicrobiana. Bionat. Lat. Am. J. Biotechnol. Life Sci. 2021, 6, 1917–1923. [Google Scholar] [CrossRef]
- Zhao, Z.; Ma, Y.; Dai, C.; Zhao, R.; Li, S.; Wu, Y.; Cao, Z.; Li, W. Imcroporin, a New Cationic Antimicrobial Peptide from the Venom of the Scorpion Isometrus maculates. Antimicrob. Agents Chemother. 2009, 53, 3472–3477. [Google Scholar] [CrossRef]
- de Barros, E.; Gonçalves, R.M.; Cardoso, M.H.; Santos, N.C.; Franco, O.L.; Cândido, E.S. Snake Venom Cathelicidins as Natural Antimicrobial Peptides. Front. Pharmacol. 2019, 10, 1415. [Google Scholar] [CrossRef]
- Lee, B.; Shin, M.K.; Yoo, J.S.; Jang, W.; Sung, J.-S. Identifying Novel Antimicrobial Peptides from Venom Gland of Spider Pardosa Astrigera by Deep Multi-Task Learning. Front. Microbiol. 2022, 13, 971503. [Google Scholar] [CrossRef]
- Ko, S.J.; Park, E.; Asandei, A.; Choi, J.-Y.; Lee, S.-C.; Seo, C.H.; Luchian, T.; Park, Y. Bee Venom-Derived Antimicrobial Peptide Melectin Has Broad-Spectrum Potency, Cell Selectivity, and Salt-Resistant Properties. Sci. Rep. 2020, 10, 10145. [Google Scholar] [CrossRef]
- Abd El-Wahed, A.A.; Khalifa, S.A.M.; Sheikh, B.Y.; Farag, M.A.; Saeed, A.; Larik, F.A.; Koca-Caliskan, U.; AlAjmi, M.F.; Hassan, M.; Wahabi, H.A.; et al. Bee Venom Composition: From Chemistry to Biological Activity. In Studies in Natural Products Chemistry; Elsevier: Amsterdam, The Netherlands, 2019; Volume 60, pp. 459–484. ISBN 978-0-444-64181-6. [Google Scholar]
- Amin, M.A.; Abdel-Raheem, I.T. Accelerated Wound Healing and Anti-Inflammatory Effects of Physically Cross Linked Polyvinyl Alcohol–Chitosan Hydrogel Containing Honey Bee Venom in Diabetic Rats. Arch. Pharm. Res. 2014, 37, 1016–1031. [Google Scholar] [CrossRef]
- Ward, R.; Coffey, M.; Kavanagh, K. Proteomic Analysis of Summer and Winter Apis mellifera Workers Shows Reduced Protein Abundance in Winter Samples. J. Insect Physiol. 2022, 139, 104397. [Google Scholar] [CrossRef]
- Kwon, N.-Y.; Sung, S.-H.; Sung, H.-K.; Park, J.-K. Anticancer Activity of Bee Venom Components against Breast Cancer. Toxins 2022, 14, 460. [Google Scholar] [CrossRef]
- Kim, H.; Park, S.-Y.; Lee, G. Potential Therapeutic Applications of Bee Venom on Skin Disease and Its Mechanisms: A Literature Review. Toxins 2019, 11, 374. [Google Scholar] [CrossRef]
- Hegazi, A.; Abdou, A.M.; El-Moez, S.I.A.; Allah, F.A. Evaluation of the Antibacterial Activity of Bee Venom from Different Sources. World Appl. Sci. J. 2014, 30, 266–270. [Google Scholar]
- Nguyen, C.D.; Yoo, J.; Hwang, S.-Y.; Cho, S.-Y.; Kim, M.; Jang, H.; No, K.O.; Shin, J.C.; Kim, J.-H.; Lee, G. Bee Venom Activates the Nrf2/HO-1 and TrkB/CREB/BDNF Pathways in Neuronal Cell Responses against Oxidative Stress Induced by Aβ1–42. Int. J. Mol. Sci. 2022, 23, 1193. [Google Scholar] [CrossRef]
- Tanner, C.M.; Kamel, F.; Ross, G.W.; Hoppin, J.A.; Goldman, S.M.; Korell, M.; Marras, C.; Bhudhikanok, G.S.; Kasten, M.; Chade, A.R.; et al. Rotenone, Paraquat, and Parkinson’s Disease. Environ. Health Perspect. 2011, 119, 866–872. [Google Scholar] [CrossRef]
- Khalil, W.K.B.; Assaf, N.; ElShebiney, S.A.; Salem, N.A. Neuroprotective Effects of Bee Venom Acupuncture Therapy against Rotenone-Induced Oxidative Stress and Apoptosis. Neurochem. Int. 2015, 80, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Socarras, K.; Theophilus, P.; Torres, J.; Gupta, K.; Sapi, E. Antimicrobial Activity of Bee Venom and Melittin against Borrelia Burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef]
- Zolfagharian, H.; Mohajeri, M.; Babaie, M. Bee Venom (Apis mellifera) an Effective Potential Alternative to Gentamicin for Specific Bacteria Strains: Bee Venom an Effective Potential for Bacteria. J. Pharmacopunct. 2016, 19, 225–230. [Google Scholar] [CrossRef]
- Han, S.; Kim, J.; Hong, I.; Woo, S.; Kim, S.; Jang, H.; Pak, S. Antibacterial Activity and Antibiotic-Enhancing Effects of Honeybee Venom against Methicillin-Resistant Staphylococcus Aureus. Molecules 2016, 21, 79. [Google Scholar] [CrossRef]
- Leandro, L.F.; Mendes, C.A.; Casemiro, L.A.; Vinholis, A.H.C.; Cunha, W.R.; de Almeida, R.; Martins, C.H.G. Antimicrobial Activity of Apitoxin, Melittin and Phospholipase A2 of Honey Bee (Apis mellifera) Venom against Oral Pathogens. Ann. Acad. Bras. Ciênc. 2015, 87, 147–155. [Google Scholar] [CrossRef] [PubMed]
- El-Didamony, S.E.; Kalaba, M.H.; El-Fakharany, E.M.; Sultan, M.H.; Sharaf, M.H. Antifungal and Antibiofilm Activities of Bee Venom Loaded on Chitosan Nanoparticles: A Novel Approach for Combating Fungal Human Pathogens. World J. Microbiol. Biotechnol. 2022, 38, 244. [Google Scholar] [CrossRef]
- Uddin, M.B.; Lee, B.-H.; Nikapitiya, C.; Kim, J.-H.; Kim, T.-H.; Lee, H.-C.; Kim, C.G.; Lee, J.-S.; Kim, C.-J. Inhibitory Effects of Bee Venom and Its Components against Viruses In Vitro and In Vivo. J. Microbiol. 2016, 54, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Fenard, D.; Lambeau, G.; Valentin, E.; Lefebvre, J.C.; Lazdunski, M.; Doglio, A. Secreted Phospholipases A(2), a New Class of HIV Inhibitors That Block Virus Entry into Host Cells. J. Clin. Investig. 1999, 104, 611–618. [Google Scholar] [CrossRef]
- de Brito, J.C.M.; Bastos, E.M.A.F.; Heneine, L.G.D.; de Souza Figueiredo, K.C. Fractionation of Apis mellifera Venom by Means of Ultrafiltration: Removal of Phospholipase A2. Braz. J. Chem. Eng. 2018, 35, 229–236. [Google Scholar] [CrossRef]
- El-Seedi, H.; Abd El-Wahed, A.; Yosri, N.; Musharraf, S.G.; Chen, L.; Moustafa, M.; Zou, X.; Al-Mousawi, S.; Guo, Z.; Khatib, A.; et al. Antimicrobial Properties of Apis mellifera’s Bee Venom. Toxins 2020, 12, 451. [Google Scholar] [CrossRef] [PubMed]
- Memariani, H.; Memariani, M.; Moravvej, H.; Shahidi-Dadras, M. Melittin: A Venom-Derived Peptide with Promising Anti-Viral Properties. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Kurek-Górecka, A.; Komosinska-Vassev, K.; Rzepecka-Stojko, A.; Olczyk, P. Bee Venom in Wound Healing. Molecules 2020, 26, 148. [Google Scholar] [CrossRef] [PubMed]
- Huh, J.-E.; Seo, B.-K.; Lee, J.-W.; Park, Y.-C.; Baek, Y.-H. Analgesic Effects of Diluted Bee Venom Acupuncture Mediated by δ-Opioid and A2-Adrenergic Receptors in Osteoarthritic Rats. Altern. Ther. Health Med. 2018, 24, 28–35. [Google Scholar] [PubMed]
- Maitip, J.; Mookhploy, W.; Khorndork, S.; Chantawannakul, P. Comparative Study of Antimicrobial Properties of Bee Venom Extracts and Melittins of Honey Bees. Antibiotics 2021, 10, 1503. [Google Scholar] [CrossRef]
- Haktanir, I.; Masoura, M.; Mantzouridou, F.T.; Gkatzionis, K. Mechanism of Antimicrobial Activity of Honeybee (Apis mellifera) Venom on Gram-Negative Bacteria: Escherichia coli and Pseudomonas spp. AMB Expr. 2021, 11, 54. [Google Scholar] [CrossRef]
- Bakhiet, E.K.; Hussien, H.A.M.; Elshehaby, M. Apis mellifera Venom Inhibits Bacterial and Fungal Pathogens In Vitro. Pak. J. Biol. Sci. 2022, 25, 875–884. [Google Scholar] [CrossRef] [PubMed]
- Babaie, M.; Ghaem panah, A.; Mehrabi, Z.; Mollaei, A.; Sima Khalilifard, B. Partial Purification and Characterization of Antimicrobial Effects from Snake (Echis carinatus), Scorpion (Mesosobuthus epues) and Bee (Apis mellifera) Venoms. Iran. J. Med. Microbiol. 2020, 14, 460–477. [Google Scholar] [CrossRef]
- Pucca, M.B.; Cerni, F.A.; Oliveira, I.S.; Jenkins, T.P.; Argemí, L.; Sørensen, C.V.; Ahmadi, S.; Barbosa, J.E.; Laustsen, A.H. Bee Updated: Current Knowledge on Bee Venom and Bee Envenoming Therapy. Front. Immunol. 2019, 10, 2090. [Google Scholar] [CrossRef]
- van den Bogaart, G.; Guzmán, J.V.; Mika, J.T.; Poolman, B. On the Mechanism of Pore Formation by Melittin. J. Biol. Chem. 2008, 283, 33854–33857. [Google Scholar] [CrossRef]
- Zarrinnahad, H.; Mahmoodzadeh, A.; Hamidi, M.P.; Mahdavi, M.; Moradi, A.; Bagheri, K.P.; Shahbazzadeh, D. Apoptotic Effect of Melittin Purified from Iranian Honey Bee Venom on Human Cervical Cancer HeLa Cell Line. Int. J. Pept. Res. Ther. 2018, 24, 563–570. [Google Scholar] [CrossRef]
- de Roodt, A.R.; Lanari, L.C.; Lago, N.R.; Bustillo, S.; Litwin, S.; Morón-Goñi, F.; Gould, E.G.; van Grootheest, J.H.; Dokmetjian, J.C.; Dolab, J.A.; et al. Toxicological Study of Bee Venom (Apis mellifera Mellifera) from Different Regions of the Province of Buenos Aires, Argentina. Toxicon 2020, 188, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Viana, G.A.; Freitas, C.I.A.; Almeida, J.G.L.d.; Medeiros, G.V.D.d.; Teófilo, T.d.S.; Rodrigues, V.H.V.; Coelho, W.A.C.; Batista, J.S. Antioxidant, Genotoxic, Antigenotoxic, and Antineoplastic Activities of Apitoxin Produced by Apis mellifera in Northeast, Brazil. Cienc. Rural 2021, 51, e20200545. [Google Scholar] [CrossRef]
- Frangieh, J.; Salma, Y.; Haddad, K.; Mattei, C.; Legros, C.; Fajloun, Z.; El Obeid, D. First Characterization of the Venom from Apis mellifera Syriaca, a Honeybee from the Middle East Region. Toxins 2019, 11, 191. [Google Scholar] [CrossRef] [PubMed]
- Somwongin, S.; Chantawannakul, P.; Chaiyana, W. Antioxidant Activity and Irritation Property of Venoms from Apis Species. Toxicon 2018, 145, 32–39. [Google Scholar] [CrossRef]
- Tanuwidjaja, I.; Svečnjak, L.; Gugić, D.; Levanić, M.; Jurić, S.; Vinceković, M.; Mrkonjić Fuka, M. Chemical Profiling and Antimicrobial Properties of Honey Bee (Apis mellifera L.) Venom. Molecules 2021, 26, 3049. [Google Scholar] [CrossRef]
- Yaacoub, C.; Rifi, M.; El-Obeid, D.; Mawlawi, H.; Sabatier, J.-M.; Coutard, B.; Fajloun, Z. The Cytotoxic Effect of Apis mellifera Venom with a Synergistic Potential of Its Two Main Components—Melittin and PLA2—On Colon Cancer HCT116 Cell Lines. Molecules 2021, 26, 2264. [Google Scholar] [CrossRef]
- Kamel, A.; Suleiman, W.; Elfeky, A.; El-Sherbiny, G.; Elhaw, M. Characterization of Bee Venom and Its Synergistic Effect Combating Antibiotic Resistance of Pseudomonas aeruginosa. Egypt. J. Chem. 2021, 65, 1–2. [Google Scholar] [CrossRef]
- Nehme, H.; Ayde, H.; El Obeid, D.; Sabatier, J.M.; Fajloun, Z. Potential Inhibitory Effect of Apis mellifera’s Venom and of Its Two Main Components—Melittin and PLA2—On Escherichia coli F1F0-ATPase. Antibiotics 2020, 9, 824. [Google Scholar] [CrossRef]
- Sobral, F.; Sampaio, A.; Falcão, S.; Queiroz, M.J.R.P.; Calhelha, R.C.; Vilas-Boas, M.; Ferreira, I.C.F.R. Chemical Characterization, Antioxidant, Anti-Inflammatory and Cytotoxic Properties of Bee Venom Collected in Northeast Portugal. Food Chem. Toxicol. 2016, 94, 172–177. [Google Scholar] [CrossRef]
- Noble, J.E. Quantification of Protein Concentration Using UV Absorbance and Coomassie Dyes. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 536, pp. 17–26. ISBN 978-0-12-420070-8. [Google Scholar]
- Pérez-Delgado, O.; Espinoza-Vergara, M.A.; Castro-Vega, N.A.; Reyes-Montaño, E.A. Evaluación Preliminar de Actividad Antibacteriana in Vitro Del Veneno de Escorpión Hadruroides Charcasus (Karsch, 1879) Contra Pseudomonas aeruginosa y Staphylococcus aureus. Rev. Cuerpo Med. HNAAA 2019, 12, 6–12. [Google Scholar] [CrossRef]
- Schägger, H.; von Jagow, G. Tricine-Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis for the Separation of Proteins in the Range from 1 to 100 KDa. Anal. Biochem. 1987, 166, 368–379. [Google Scholar] [CrossRef] [PubMed]
- Brunelle, J.L.; Green, R. Coomassie Blue Staining. In Methods in Enzymology; Elsevier: Amsterdam, The Netherlands, 2014; Volume 541, pp. 161–167. ISBN 978-0-12-420119-4. [Google Scholar]
- Approved Standard M07; Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, 9th ed. Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012; Volume 32, ISBN 1-56238-784-7.
- Oddo, A.; Hansen, P.R. Hemolytic Activity of Antimicrobial Peptides. In Antimicrobial Peptides; Hansen, P.R., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2017; Volume 1548, pp. 427–435. ISBN 978-1-4939-6735-3. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Delgado, O.; Espinoza-Culupú, A.O.; López-López, E. Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru. Antibiotics 2023, 12, 779. https://doi.org/10.3390/antibiotics12040779
Pérez-Delgado O, Espinoza-Culupú AO, López-López E. Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru. Antibiotics. 2023; 12(4):779. https://doi.org/10.3390/antibiotics12040779
Chicago/Turabian StylePérez-Delgado, Orlando, Abraham Omar Espinoza-Culupú, and Elmer López-López. 2023. "Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru" Antibiotics 12, no. 4: 779. https://doi.org/10.3390/antibiotics12040779
APA StylePérez-Delgado, O., Espinoza-Culupú, A. O., & López-López, E. (2023). Antimicrobial Activity of Apis mellifera Bee Venom Collected in Northern Peru. Antibiotics, 12(4), 779. https://doi.org/10.3390/antibiotics12040779