


Persistence of Campylobacter spp. in Poultry Flocks after Disinfection, Virulence, and Antimicrobial Resistance Traits of Recovered Isolates

Abstract
:1. Introduction
2. Results
2.1. Biosecurity Measures
2.1.1. Environment of the Farm
2.1.2. Characteristics of the Farm
2.1.3. Sanitary Measures
2.1.4. Personnel Hygiene
2.2. Contamination Rates of Campylobacter spp. in Breeding Hens and Environment before and after Cleaning and Disinfection
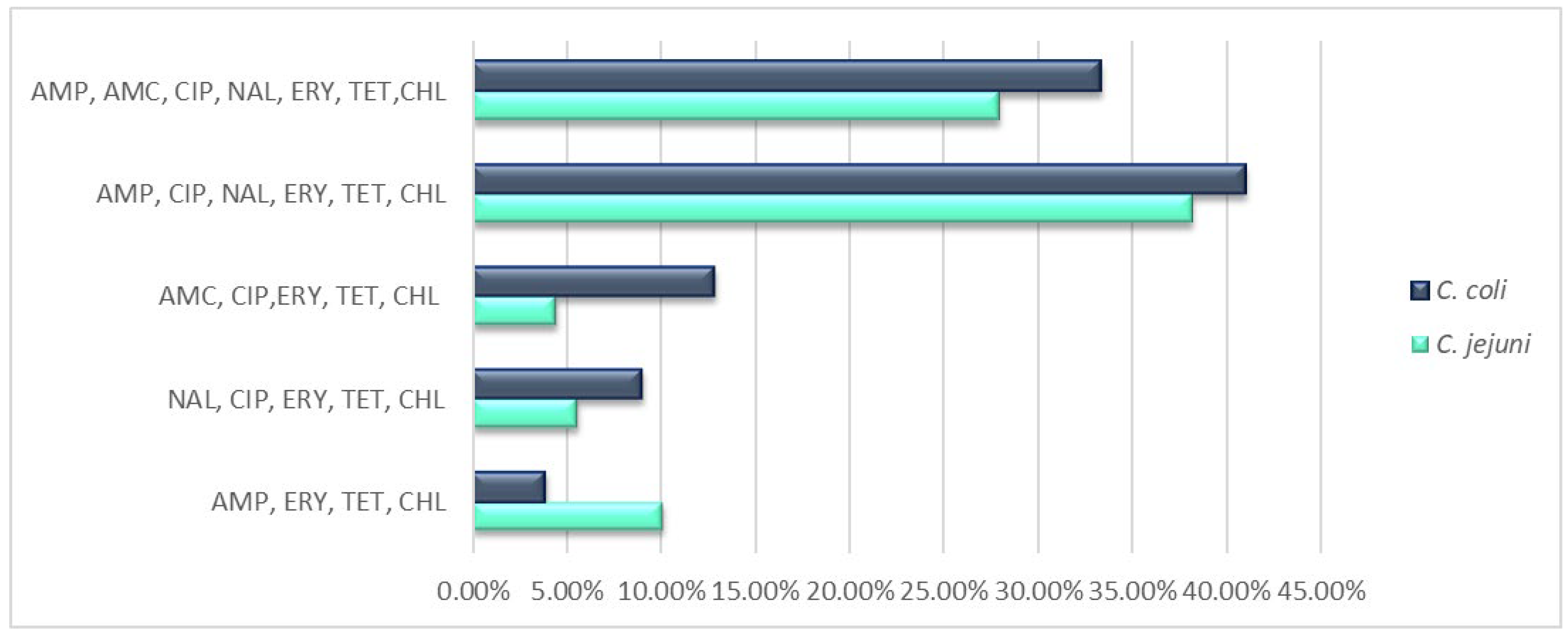
2.3. Antimicrobial Resistance Patterns
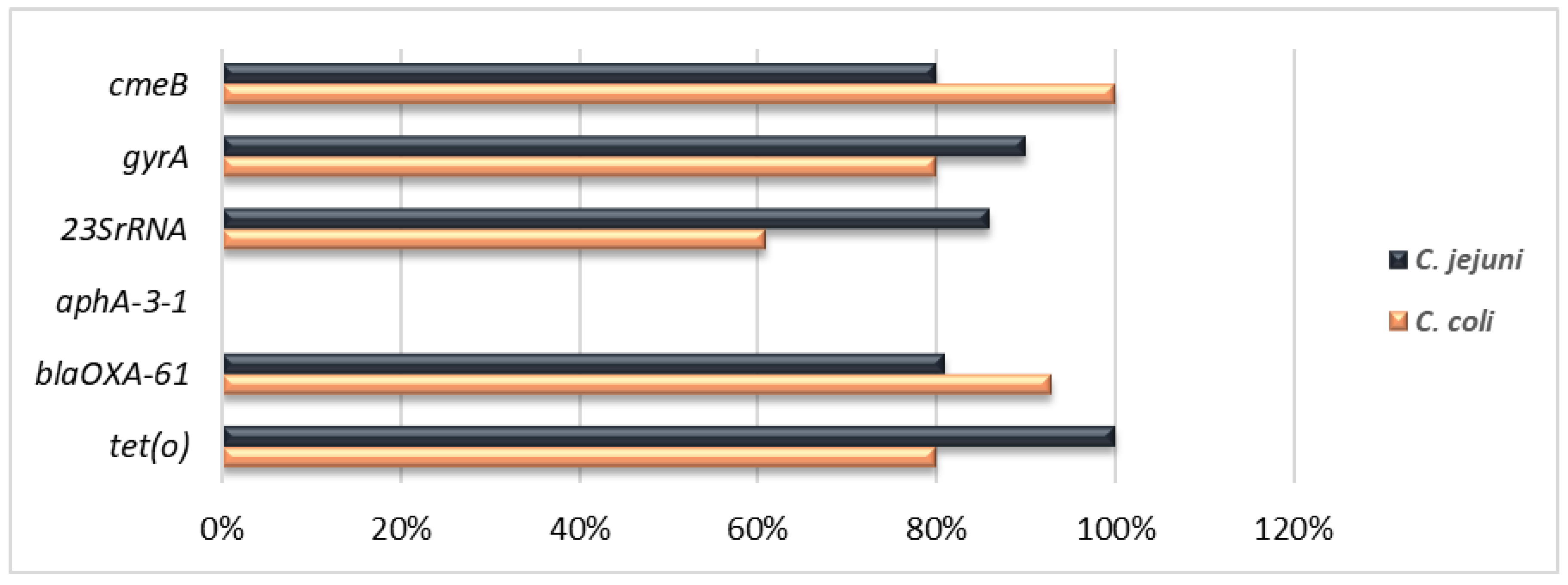
2.4. Detection of Antimicrobial Resistance Genes
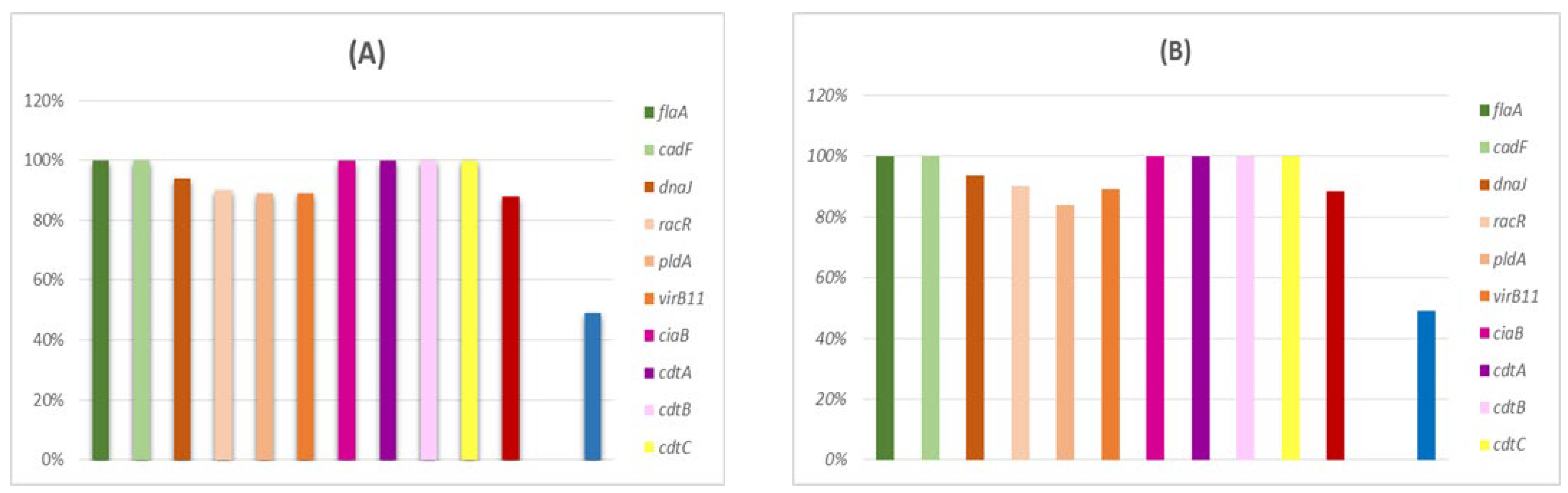
2.5. Prevalence of Genes Encodingvirulence Factors
3. Discussion
3.1. Prevalence of Campylobacter and Risk Factors
3.2. Antimicrobial Resistances and Implicated Genes
3.3. Occurrence of Virulence Genes
4. Materials and Methods
4.1. Samples Collection
4.2. Audit of Cleaning and Disinfection Procedures in Poultry Farm Buildings
4.3. Sample Transportation and Processing
4.4. Culture and Growth Conditions
4.5. DNA Extraction
4.6. Antimicrobial Susceptibility Test
4.7. Detection of Mutation(s) in the QRDR of gyrA and 23S rRNA Genes by PCR Mismatch Amplification Mutation Assay (MAMA-PCR)
4.8. Detection of Their Resistance Determinants
4.9. Detection of Virulence Genes
4.10. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Igwaran, A.; Okoh, A.I. Human Campylobacteriosis: A Public Health Concern of Global Importance. Heliyon 2019, 5, e0281. [Google Scholar] [CrossRef] [PubMed]
- Bolton, D.J. Campylobacter Virulence and Survival Factors. Food Microbiol. 2015, 48, 99–108. [Google Scholar] [CrossRef]
- García-Fernández, A.; Dionisi, A.M.; Arena, S.; Iglesias-Torrens, Y.; Carattoli, A.; Luzzi, I. Human Campylobacteriosis in Italy: Emergence of Multi-Drug Resistance to Ciprofloxacin, Tetracycline, and Erythromycin. Front. Microbiol. 2018, 9, 1906. [Google Scholar] [CrossRef] [PubMed]
- Food, E.; Authority, S. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2017. EFSA J. 2018, 16, e05500. [Google Scholar]
- Hermans, D.; Pasmans, F.; Messens, W.; Martel, A.; Van Immerseel, F.; Rasschaert, G.; Heyndrickx, M.; Van Deun, K.; Haesebrouck, F. Poultry as a Host for the Zoonotic Pathogen Campylobacter jejuni. Vector Borne Zoonotic Dis. 2012, 12, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Pielsticker, C.; Glünder, G.; Rautenschlein, S. Colonization Properties of Campylobacter Jejuni in Chickens. Eur. J. Microbiol. Immunol. 2012, 2, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Jacobs-Reitsma, W.F. Aspects of Epidemiology of Campylobacter in Poultry. Vet. Q 1997, 19, 113–117. [Google Scholar] [CrossRef]
- Van De Giessen, A.W.; Tilburg, J.J.H.C.; Ritmeester, W.S.; Van Der Plas, J. Reduction of Campylobacter Infections in Broiler Flocks by Application of Hygiene Measures. Epidemiol. Infect. 1998, 121, 57–66. [Google Scholar] [CrossRef]
- Puterflam, J.; Bouvarel, I.R.; Ragot, O.; Drouet, M. Contamination des élévages de poulets de chair par Campylobacter: Est-ce une fatalité. Sci. Technol. Avic. 2005, 53, 12–19. [Google Scholar]
- Van De Giessen, A.W.; Bloemberg, B.P.M.; Ritmeester, W.S.; Tilburg, J.J.H.C. Epidemiological Study on Risk Factors and Risk Reducing Measures for Campylobacter Infections in Dutch Broiler Flocks. Epidemiol. Infect. 1996, 117, 245–250. [Google Scholar] [CrossRef]
- Shane, S.M. The Significance of Campylobacter Jejuni Infection in Poultry: A Review. Avian Pathol. 1992, 21, 189–213. [Google Scholar] [CrossRef] [PubMed]
- FAO. CÔTE D’IVOIRE: Revue Du Secteur Avicole; FAO: Rome, Italy, 2008; p. 77. [Google Scholar]
- Van Immerseel, F.; Boyen, F.; Gantois, I.; Timbermont, L.; Bohez, L.; Pasmans, F.; Haesebrouck, F.; Ducatelle, R. Supplementation of Coated Butyric Acid in the Feed Reduces Colonization and Shedding of Salmonella in Poultry. Poult. Sci. 2005, 84, 1851–1856. [Google Scholar] [CrossRef] [PubMed]
- Badji, A.; Lo, M.M.; Alambedji, R.B. Facteurs de Risque Liés à l’apparition de La Maladie de Gumboro En Aviculture Semi-Intensive Au Sénégal. Int. J. Biol. Chem. Sci. 2017, 11, 1551. [Google Scholar] [CrossRef]
- Hendrix, S.L.; Clark, A.; Nygaard, I.; Aragaki, A.; Barnabei, V.; McTiernan, A. Pelvic Organ Prolapse in the Women’s Health Initiative: Gravity and Gravidity. Am. J. Obstet. Gynecol. 2002, 186, 1160–1166. [Google Scholar] [CrossRef] [PubMed]
- Kaci, M.; Sadoun, T.; Cimmino, S. Crystallinity Measurements of Unstabilized and HALS-Stabilized LDPE Films Exposed to Natural Weathering by FT-IR, DSC and WAXS Analyses. Int. J. Polym. Anal. Charact. 2001, 6, 455–464. [Google Scholar] [CrossRef]
- Jussiau, R.; Papet, A.; Rigal, J. Amélioration Génétique des Animaux D’élevage (Edition 2013); Educagri Editions: Dijon, France, 2013. [Google Scholar]
- Wei, B.; Kang, M.; Jang, H.K. Genetic Characterization and Epidemiological Implications of Campylobacter Isolates from Wild Birds in South Korea. Transbound. Emerg. Dis. 2019, 66, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.K.; Rigby, D.; Burton, M.; Millman, C.; Williams, N.J.; Jones, T.R.; Wigley, P.; O’Brien, S.J.; Cross, P. Restaurant Cooking Trends and Increased Risk for Campylobacter Infection. Emerg. Infect. Dis. 2016, 22, 1208–1215. [Google Scholar] [CrossRef]
- Novoa Rama, E.; Bailey, M.; Jones, D.R.; Gast, R.K.; Anderson, K.; Brar, J.; Taylor, R.; Oliver, H.F.; Singh, M. Prevalence, Persistence, and Antimicrobial Resistance of Campylobacter Spp. from Eggs and Laying Hens Housed in Five Commercial Housing Systems. Foodborne Pathog. Dis. 2018, 15, 506–516. [Google Scholar] [CrossRef]
- Sulonen, J.; Kärenlampi, R.; Holma, U.; Hänninen, M.L. Campylobacter in Finnish Organic Laying Hens in Autumn 2003 and Spring 2004. Poult. Sci. 2007, 86, 1223–1228. [Google Scholar] [CrossRef]
- Dipineto, L.; Gargiulo, A.; Russo, T.P.; De Luca Bossa, L.M.; Borrelli, L.; Menna, L.F.; Fioretti, A. Campylobacter Jejuni, Campylobacter Coli, and Cytolethal Distending Toxin Genes in Laying Hens. Avian Dis. 2011, 55, 103–105. [Google Scholar] [CrossRef]
- Kalupahana, R.S.; Kottawatta, K.S.A.; Kanankege, K.S.T.; Van Bergen, M.A.P.; Abeynayake, P.; Wagenaar, J.A. Colonization of Campylobacter Spp. in Broiler Chickens and Laying Hens Reared in Tropical Climates with Low-Biosecurity Housing. Appl. Environ. Microbiol. 2013, 79, 393–395. [Google Scholar] [CrossRef] [PubMed]
- Van Asselt, M.; Poortvliet, P.M.; Ekkel, E.D.; Kemp, B.; Stassen, E.N. Risk Perceptions of Public Health and Food Safety Hazards in Poultry Husbandry by Citizens, Poultry Farmers and Poultry Veterinarians. Poult. Sci. 2018, 97, 607–619. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, K.; Yamamoto, T.; Satake, S.; Takayama, S.; Kubota, S.; Negishi, H.; Kojima, A.; Asai, T.; Sawada, T.; Takahashi, T.; et al. Comparison of Campylobacter Isolated from Humans and Food-Producing Animals in Japan. J. Appl. Microbiol. 2006, 100, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Neves, R.A.F.; Santiago, T.C.; Carvalho, W.F.; Silva, E.d.S.; da Silva, P.M.; Nascimento, S.M. Impacts of the Toxic Benthic Dinoflagellate Prorocentrum Lima on the Brown Mussel Perna Perna: Shell-Valve Closure Response, Immunology, and Histopathology. Mar. Environ. Res. 2019, 146, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Pergola, S.; Franciosini, M.P.; Comitini, F.; Ciani, M.; De Luca, S.; Bellucci, S.; Menchetti, L.; Casagrande Proietti, P. Genetic Diversity and Antimicrobial Resistance Profiles of Campylobacter Coli and Campylobacter Jejuni Isolated from Broiler Chicken in Farms and at Time of Slaughter in Central Italy. J. Appl. Microbiol. 2017, 122, 1348–1356. [Google Scholar] [CrossRef]
- Mezher, Z.; Saccares, S.; Marcianò, R.; de Santis, P.; Flores Rodas, E.M.; De Angelis, V.; Condoleo, R. Occurrence of Campylobacter Spp. In Poultry Meat at Retail and Processing Plants’ Levels in Central Italy. Ital. J. Food Saf. 2016, 5, 47–49. [Google Scholar] [CrossRef]
- Sahin, O.; Kassem, I.I.; Shen, Z.; Lin, J.; Rajashekara, G.; Zhang, Q. Campylobacter in Poultry: Ecology and Potential Interventions. Avian Dis. 2015, 59, 185–200. [Google Scholar] [CrossRef]
- Marinou, I.; Bersimis, S.; Ioannidis, A.; Nicolaou, C.; Mitroussia-Ziouva, A.; Legakis, N.J.; Chatzipanagiotou, S. Identification and Antimicrobial Resistance of Campylobacter Species Isolated from Animal Sources. Front. Microbiol. 2012, 3, 58. [Google Scholar] [CrossRef]
- Puterflam, J.; Michel, J.; Hocking, P.M.; Picard, M. Effects of Genotype and Feed Restriction on the Time-Budgets of Broiler Breeders at Different Ages. Appl. Anim. Behav. Sci. 2006, 98, 100–113. [Google Scholar] [CrossRef]
- Snelling, W.J.; Matsuda, M.; Moore, J.E.; Dooley, J.S.G. Under the Microscope: Campylobacter Jejuni. Lett. Appl. Microbiol. 2005, 41, 297–302. [Google Scholar] [CrossRef]
- Guérin, F.; Abril, G. Significance of Pelagic Aerobic Methane Oxidation in the Methane and Carbon Budget of a Tropical Reservoir. J. Geophys. Res. 2007, 112, 1–14. [Google Scholar] [CrossRef]
- Fani, F.; Aminshahidi, M.; Firoozian, N.; Rafaatpour, N. Prevalence, Antimicrobial Resistance, and Virulence-Associated Genes of Campylobacter Isolates from Raw Chicken Meat in Shiraz, Iran. Iran. J. Vet. Res. 2019, 20, 283–288. [Google Scholar] [PubMed]
- Morgan, R.B.; Sierra–Arguello, Y.M.; Perdoncini, G.; Borges, K.A.; Furian, T.Q.; Gomes, M.J.P.; Lima, D.; Salle, C.T.P.; Moraes, H.L.S.; Nascimento, V.P. Comparison of Transport Crates Contamination with Campylobacter spp. before and after the Cleaning and Disinfection Procedure in Broiler Slaughterhouses. Poult. Sci. 2022, 101, 101909. [Google Scholar] [CrossRef] [PubMed]
- Peyrat, M.B.; Soumet, C.; Maris, P.; Sanders, P. Phenotypes and Genotypes of Campylobacter Strains Isolated after Cleaning and Disinfection in Poultry Slaughterhouses. Vet. Microbiol. 2008, 128, 313–326. [Google Scholar] [CrossRef]
- Hasan, M.M.; Talukder, S.; Mandal, A.K.; Tasmim, S.T.; Parvin, M.S.; Ali, M.Y.; Sikder, M.H.; Islam, M.T. Prevalence and Risk Factors of Campylobacter Infection in Broiler and Cockerel Flocks in Mymensingh and Gazipur Districts of Bangladesh. Prev. Vet. Med. 2020, 180, 105034. [Google Scholar] [CrossRef]
- Insights, C. Relationships between Virulence Genes and Antibiotic Resistance Phenotypes/Genotypes in Campylobacter Spp. Foods 2022, 11, 3554. [Google Scholar]
- Gharbi, M.; Béjaoui, A.; Ben Hamda, C.; Jouini, A.; Ghedira, K.; Zrelli, C.; Hamrouni, S.; Aouadhi, C.; Bessoussa, G.; Ghram, A.; et al. Prevalence and Antibiotic Resistance Patterns of Campylobacter Spp. Isolated from Broiler Chickens in the North of Tunisia. Biomed Res. Int. 2018, 2018, 7943786. [Google Scholar] [CrossRef]
- Elhadidy, M.; Miller, W.G.; Arguello, H.; álvarez-Ordóñez, A.; Duarte, A.; Dierick, K.; Botteldoorn, N. Genetic Basis and Clonal Population Structure of Antibiotic Resistance in Campylobacter Jejuni Isolated from Broiler Carcasses in Belgium. Front. Microbiol. 2018, 9, 1014. [Google Scholar] [CrossRef]
- Lynch, C.T.; Lynch, H.; Burke, S.; Hawkins, K.; Buttimer, C.; Mc Carthy, C.; Egan, J.; Whyte, P.; Bolton, D.; Coffey, A.; et al. Antimicrobial Resistance Determinants Circulating among Thermophilic Campylobacter Isolates Recovered from Broilers in Ireland Over a One-Year Period. Antibiotics 2020, 9, 308. [Google Scholar] [CrossRef]
- Fraqueza, M.J.; Martins, A.; Borges, A.C.; Fernandes, M.H.; Fernandes, M.J.; Vaz, Y.; Bessa, R.J.B.; Barreto, A.S. Antimicrobial Resistance among Campylobacter Spp. Strains Isolated from Different Poultry Production Systems at Slaughterhouse Level. Poult. Sci. 2014, 93, 1578–1586. [Google Scholar] [CrossRef]
- Bolinger, H.K.; Zhang, Q.; Miller, W.G.; Kathariou, S. Lack of Evidence for Erm(B) Infiltration into Erythromycin-Resistant Campylobacter Coli and Campylobacter Jejuni from Commercial Turkey Production in Eastern North Carolina: A Major Turkey-Growing Region in the United States. Foodborne Pathog. Dis. 2018, 15, 698–700. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Guan, X.; Zeng, H.; Li, J.; Huang, X.; Wen, Y.; Zhao, Q.; Huang, X.; Yan, Q.; Huang, Y. Prevalence, Antimicrobial Resistance Profiles and Virulence-Associated Genes of Thermophilic Campylobacter Spp. Isolated from Ducks in a Chinese Slaughterhouse. Food Control 2019, 104, 157–166. [Google Scholar] [CrossRef]
- Oh, J.-Y.; Kwon, Y.-K.; Wei, B.; Jang, H.-K.; Lim, S.-K.; Kim, C.-H.; Jung, S.-C.; Kang, M.-S. Epidemiological Relationships of Campylobacter Jejuni Strains Isolated from Humans and Chickens in South Korea. J. Microbiol. 2017, 55, 13–20. [Google Scholar] [CrossRef]
- Otigbu, A.C.; Clarke, A.M.; Fri, J.; Akanbi, E.O.; Njom, H.A. Antibiotic Sensitivity Profiling and Virulence Potential of Campylobacter Jejuni Isolates from Estuarine Water in the Eastern Cape Province, South Africa. Int. J. Environ. Res. Public Health 2018, 15, 925. [Google Scholar] [CrossRef]
- Ramires, T.; de Oliveira, M.G.; Kleinubing, N.R.; de Fátima Rauber Würfel, S.; Mata, M.M.; Iglesias, M.A.; Lopes, G.V.; Dellagostin, O.A.; da Silva, W.P. Genetic Diversity, Antimicrobial Resistance, and Virulence Genes of Thermophilic Campylobacter Isolated from Broiler Production Chain. Brazilian J. Microbiol. 2020, 51, 2021–2032. [Google Scholar] [CrossRef] [PubMed]
- Pillay, S.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Shobo, C.O.; Perrett, K.; Bester, L.A.; Essack, S.Y. Characterisation of Campylobacter Spp. Isolated from Poultry in KwaZulu-Natal, South Africa. Antibiotics 2020, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Niwa, H.; Itoh, K. Age-Dependent Variation of Virulence-Associated Genes Retained in Campylobacter Jejuni Isolated from Chickens in a Poultry Farm. J. Vet. Med. Sci. 2009, 71, 1247–1249. [Google Scholar] [CrossRef]
- Khoshbakht, R.; Tabatabaei, M.; Hosseinzadeh, S.; Shekarforoush, S.S.; Aski, H.S. Distribution of Nine Virulence-Associated Genes in Campylobacter Jejuni and C. Coli Isolated from Broiler Feces in Shiraz, Southern Iran. Foodborne Pathog. Dis. 2013, 10, 764–770. [Google Scholar] [CrossRef]
- ISO 10272-1 (Annex E); Microbiology of Food And Animal Feeding Stuffs: Horizontal Method for Detection and Enu-Mera-Tion of Campylobacter Spp. I. Detection Method. International Organization for Standardization: Geneva, Switzerland, 2006.
- Gharbi, M.; Béjaoui, A.; Hamda, C.B.; Ghedira, K.; Ghram, A.; Maaroufi, A. Distribution of Virulence and Antibiotic Resistance Genes in Campylobacter Jejuni and Campylobacter Coli Isolated from Broiler Chickens in Tunisia. J. Microbiol. Immunol. Infect. 2022, 55, 1273–1282. [Google Scholar] [CrossRef]
- Linton, D.; Owen, R.J.; Stanley, J. Rapid Identification by PCR of the Genus Campylobacter and of Five Campylobacter Species Enteropathogenic for Man and Animals. Res. Microbiol. 1996, 147, 707–718. [Google Scholar] [CrossRef]
- Denis, M.; Soumet, C.; Rivoal, K.; Ermel, G.; Blivet, D.; Salvat, G.; Colin, P. Development of a M-PCR Assay for Simultaneous Identification of Campylobacter Jejuni and C. Coli. Lett. Appl. Microbiol. 1999, 29, 406–410. [Google Scholar] [CrossRef] [PubMed]
- CA-SFM. Comité de l’Antibiogramme-Société Française de Microbiologie, V 1.0, Mai. Int. J. Antimicrob. Agents. 2022, 21, 119. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Zirnstein, G.; Helsel, L.; Li, Y.; Swaminathan, B.; Besser, J. Characterization of GyrA Mutations Associated with Fluoroquinolone Resistance in Campylobacter Coli by DNA Sequence Analysis and MAMA PCR. FEMS Microbiol. Lett. 2000, 190, 1–7. [Google Scholar] [CrossRef]
- Zirnstein, G.; Li, Y.; Swaminathan, B.; Angulo, F. Ciprofloxacin Resistance in Campylobacter Jejuni Isolates: Detection of GyrA Resistance Mutations by Mismatch Amplification Mutation Assay PCR and DNA Sequence Analysis. J. Clin. Microbiol. 1999, 37, 3276–3280. [Google Scholar] [CrossRef]
- Alonso, R.; Mateo, E.; Churruca, E.; Martinez, I.; Girbau, C.; Fernández-Astorga, A. MAMA-PCR Assay for the Detection of Point Mutations Associated with High-Level Erythromycin Resistance in Campylobacter Jejuni and Campylobacter Coli Strains. J. Microbiol. Methods 2005, 63, 99–103. [Google Scholar] [CrossRef]
- Mchugh, M.L. Lessons in Biostatistics The Chi-Square Test of Independence. Biochem. Med. 2013, 23, 143–149. [Google Scholar] [CrossRef]
- Denis, M.; Refrégier-Petton, J.; Laisney, M.J.; Ermel, G.; Salvat, G. Campylobacter contamination in French chicken production from farm to consumers. Use of a PCR assay for detection and identification of Campylobacter jejuni and Camp. coli. J. Appl. Microbiol. 2001, 91, 255–267. [Google Scholar] [CrossRef]
- Asakura, M.; Samosornsuk, W.; Hinenoya, A.; Misawa, N.; Nishimura, K.; Matsuhisa, A.; Yamasaki, S. Development of a cytolethal distending toxin (cdt) gene-based species-specific multiplex PCR assay for the detection and identification of Campylobacter jejuni, Campylobacter coli and Campylobacter fetus. FEMS Immun. Med. Microbiol. 2008, 52, 260–266. [Google Scholar] [CrossRef]
- Pratt, A.; Korolik, V. Tetracycline resistance of Australian Campylobacter jejuni and Campylobacter coli isolates. J. Antimicrobial Chemother. 2005, 55, 452–460. [Google Scholar] [CrossRef]
- Olah, P.A.; Doetkott, C.; Fakhr, M.K.; Logue, C.M. Prevalence of the Campylobacter multi-drug efflux pump (CmeABC) in Campylobacter spp. Isolated from freshly processed Turkeys. Food Microbiol. 2006, 23, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Obeng, A.S.; Rickard, H.; Sexton, M.; Pang, Y.; Peng, H.; Barton, M. Antimicrobial susceptibilities and resistance genes in Campylobacter strains isolated from poultry and pigs in Australia. J. Appl. Microbiol. 2012, 113, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Datta, S.; Niwa, H.; Itoh, K. Prevalence of 11 pathogenic genes of Campylobacter jejuni by PCR in strains isolated from humans, poultry meat and broiler and bovine faeces. J. Med. Microbiol. 2003, 52 Pt 4, 345–348. [Google Scholar] [CrossRef] [PubMed]
- Konkel, M.E.; Gray, S.A.; Kim, B.J.; Garvis, S.G.; Yoon, J. Identification of the enteropathogens Campylobacter jejuni and Campylobacter coli based on the cadF virulence gene and its product. J. Clin. Microbiol. 1999, 37, 510–517. [Google Scholar] [CrossRef] [PubMed]
- Hickey, T.E.; McVeigh, A.L.; Scott, D.A.; Michielutti, R.E.; Bixby, A.; Carroll, S.A.; Bourgeois, A.L.; Guerry, P. Campylobacter jejuni cytolethal distending toxin mediates release of interleukin-8 from intestinal epithelial cells. Infection Immun. 2000, 68, 6535–6541. [Google Scholar] [CrossRef]
- Gonzalez, I.; Grant, K.A.; Richardson, P.T.; Park, S.F.; Collins, M.D. Specific identification of the enteropathogens Campylobacter jejuni and Campylobacter coli by using a PCR test based on the ceuE gene encoding a putative virulence determinant. J. Clin. Microbiol. 1997, 35, 759–763. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Before Disinfection | After Disinfection | ||||||
|---|---|---|---|---|---|---|---|
| Nb. Samples | NB % Isolates | Nb.Samples | NB Isolates | ||||
| C. coli | C. jejuni | C. coli | C. jejuni | ||||
| Buildings | Trays | 20 | 0 (0%) | 0 (0%) | 20 | 0 (0%) | 0 (0%) |
| Nestings boxes | 20 | 0 (0%) | 0 (0%) | 20 | 0 (0%) | 0 (0%) | |
| Plates | 20 | 12 (60%) | 0 (0%) | 20 | 0 (0%) | 0 (0%) | |
| floors | 20 | 20 (100%) | 0 (0%) | 20 | 0 (0%) | 0 (0%) | |
| Worker’s locker room | Boots | 6 | 6 (100%) | 0 | 6 | 3 (50%) | 0 (0%) |
| Slides | 6 | 0 | 0 | 6 | 0 (0%) | 0 (0%) | |
| Blouses | 6 | 0 | 0 | 6 | 0 (0%) | 0 (0%) | |
| Total | 98 | 38/98 (38.7%) | 98 | 3/98 (3%) | 0 (0%) | ||
| Source | Specie | No. | Antimicrobial Resistance Rates % (Number of Isolates) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ERY | AM | AMC | Cip | NAL | CHL | Tet | GEN | |||
| Layer hens | C. jejuni | 89 | 100% (89) | 47.19% (42) | 58.42% * (52) | 100% (89) | 100% (89) | 100% (89) | 100% (89) | 1.12% (1) |
| C. coli | 37 | 100% (37) | 81% * (30) | 20.22% (18) | 100% (37) | 100% (37) | 100% (37) | 100% (37) | 0% (0) | |
| Environment | C. coli | 41 | 100% (41) | 80.48% (33) | 21.95% (9) | 100% (41) | 100% (41) | 100% (41) | 100% (41) | 0% (0) |
| Total | 167 | 100% (167) | 62.87% (105) | 47.3% (79) | 100% (167) | 100% (167) | 100% (167) | 100% (167) | 0.6% (1) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gharbi, M.; Béjaoui, A.; Hamrouni, S.; Arfaoui, A.; Maaroufi, A. Persistence of Campylobacter spp. in Poultry Flocks after Disinfection, Virulence, and Antimicrobial Resistance Traits of Recovered Isolates. Antibiotics 2023, 12, 890. https://doi.org/10.3390/antibiotics12050890
Gharbi M, Béjaoui A, Hamrouni S, Arfaoui A, Maaroufi A. Persistence of Campylobacter spp. in Poultry Flocks after Disinfection, Virulence, and Antimicrobial Resistance Traits of Recovered Isolates. Antibiotics. 2023; 12(5):890. https://doi.org/10.3390/antibiotics12050890
Chicago/Turabian StyleGharbi, Manel, Awatef Béjaoui, Safa Hamrouni, Amel Arfaoui, and Abderrazak Maaroufi. 2023. "Persistence of Campylobacter spp. in Poultry Flocks after Disinfection, Virulence, and Antimicrobial Resistance Traits of Recovered Isolates" Antibiotics 12, no. 5: 890. https://doi.org/10.3390/antibiotics12050890
APA StyleGharbi, M., Béjaoui, A., Hamrouni, S., Arfaoui, A., & Maaroufi, A. (2023). Persistence of Campylobacter spp. in Poultry Flocks after Disinfection, Virulence, and Antimicrobial Resistance Traits of Recovered Isolates. Antibiotics, 12(5), 890. https://doi.org/10.3390/antibiotics12050890