
Microbiota and Resistome Analysis of Colostrum and Milk from Dairy Cows Treated with and without Dry Cow Therapies
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Microbiota Diversity Is Influenced by Antibiotic Treatment
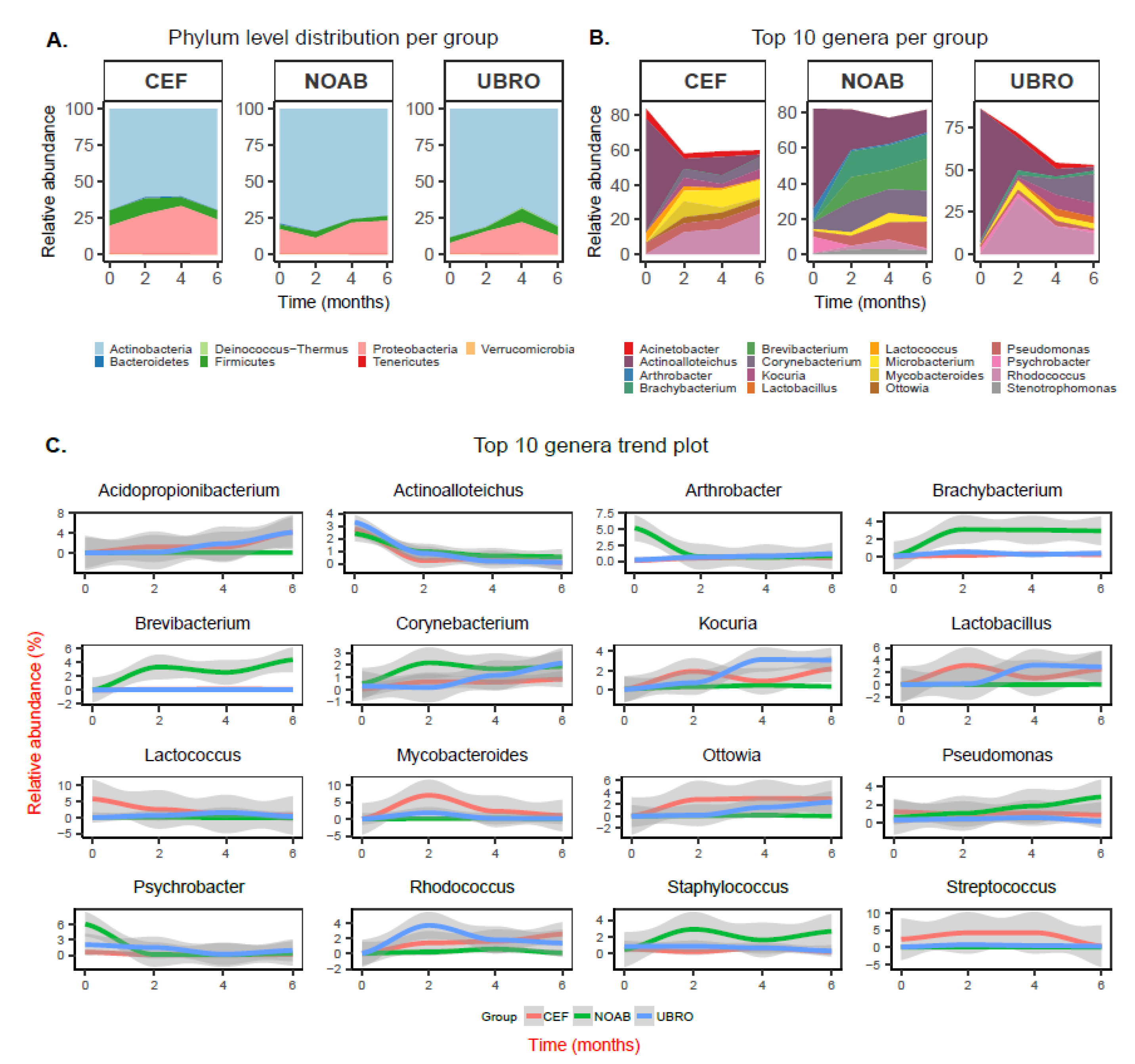
2.2. Taxonomic Composition Is Associated with Dry Cow Therapy Treatment
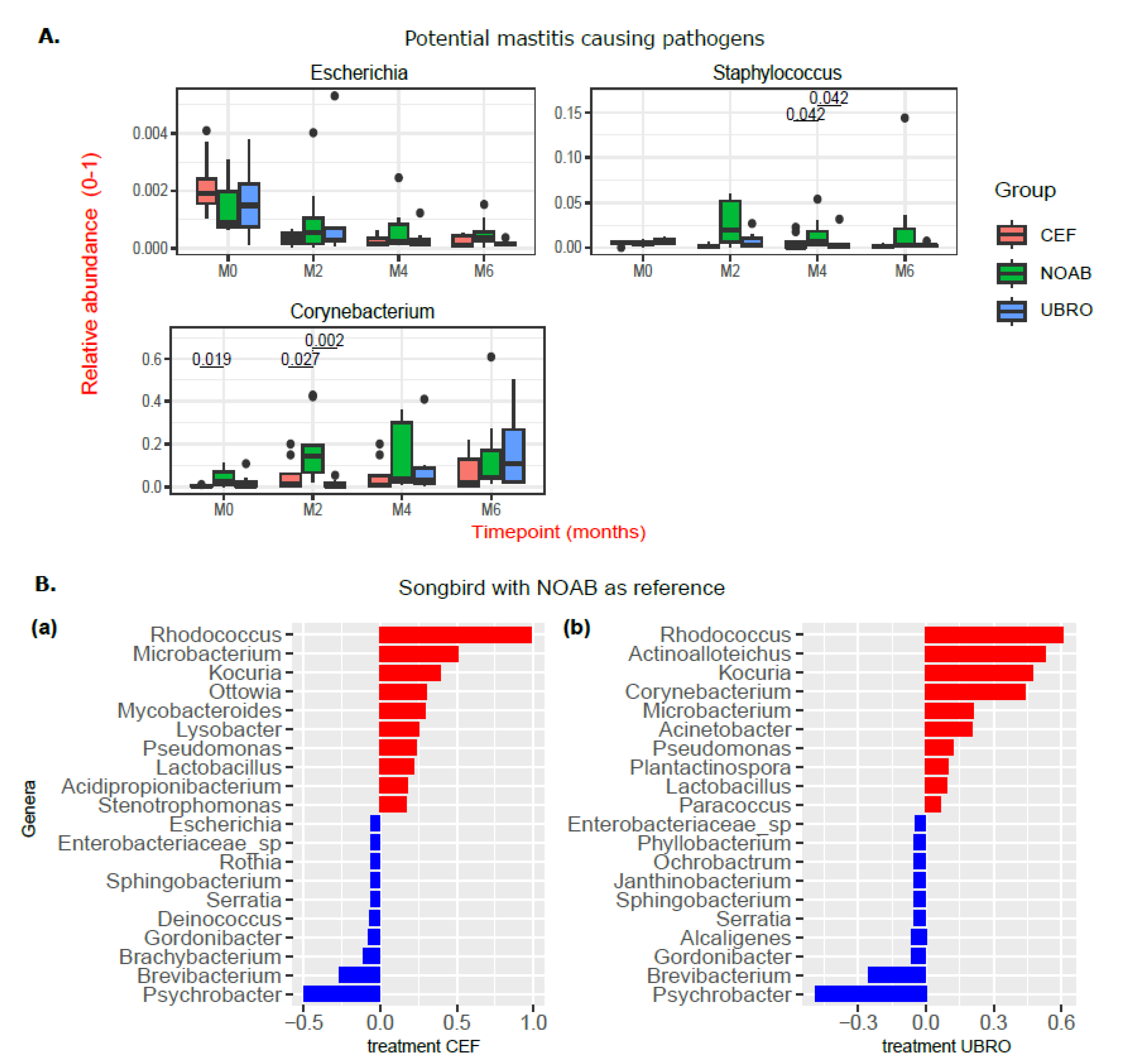
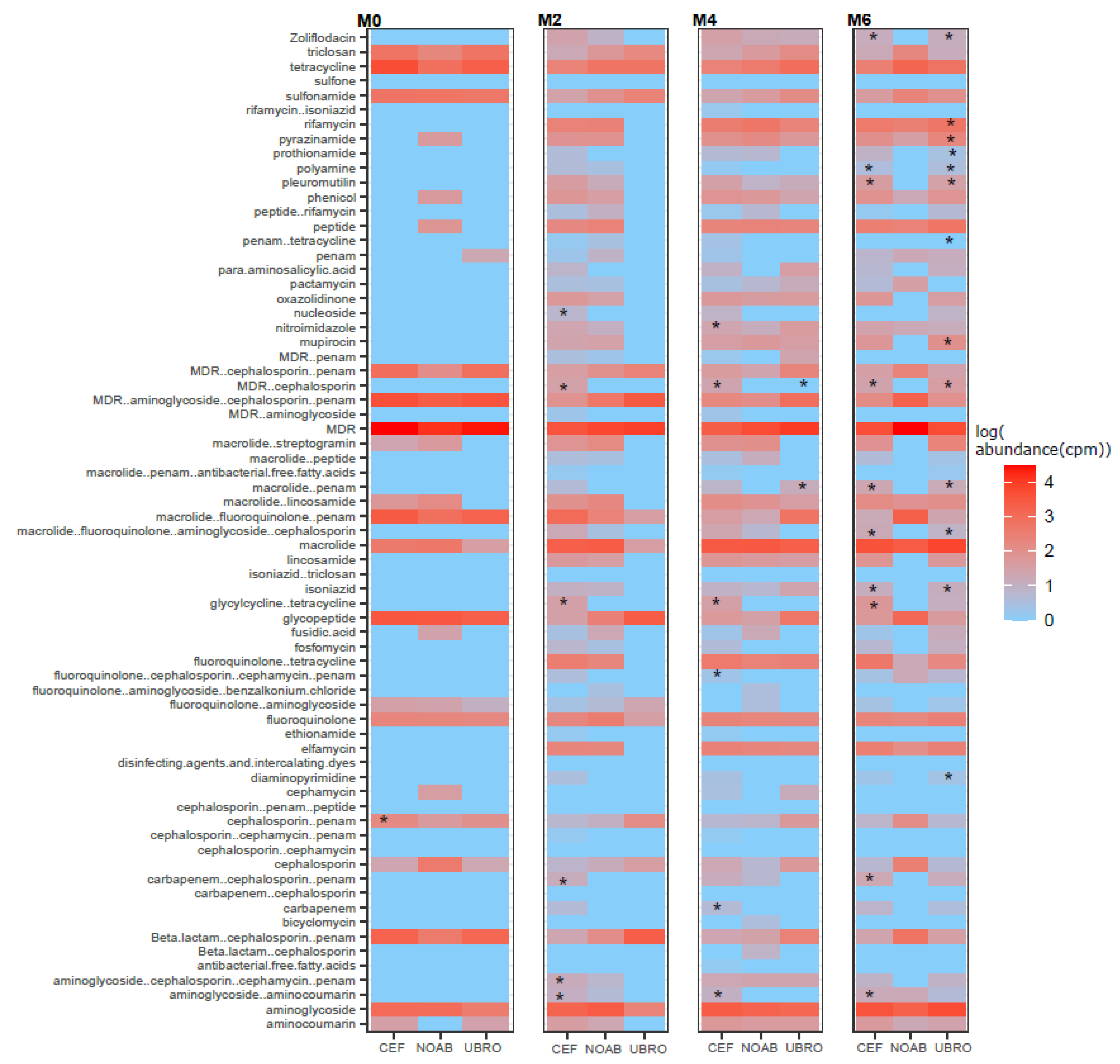
2.3. Impact of Dry Cow Therapy on Antibiotic Resistance Gene Reservoir
3. Discussion
4. Methods
4.1. Enrolment Criteria and Treatment for Cows
4.2. Sample and Data Collection
4.3. DNA Extraction
4.4. Amplification of DNA, Library Preparation and Shotgun Metagenomics Sequencing
4.5. Bioinformatics and Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, Y.; Malmuthuge, N.; Li, F.; Guan, L.L. Colostrum feeding shapes the hindgut microbiota of dairy calves during the first 12 h of life. FEMS Microbiol. Ecol. 2019, 95, fiy203. [Google Scholar] [CrossRef]
- Lopez, A.J.; Heinrichs, A.J. Invited review: The importance of colostrum in the newborn dairy calf. J. Dairy Sci. 2022, 105, 2733–2749. [Google Scholar] [CrossRef] [PubMed]
- Hammon, H.M.; Liermann, W.; Frieten, D.; Koch, C. Review: Importance of colostrum supply and milk feeding intensity on gastrointestinal and systemic development in calves. Animal 2020, 14 (Suppl. S1), S133–S143. [Google Scholar] [CrossRef] [PubMed]
- Amin, N.; Seifert, J. Dynamic progression of the calf’s microbiome and its influence on host health. Comput. Struct. Biotechnol. J. 2021, 19, 989–1001. [Google Scholar] [CrossRef]
- Oikonomou, G.; Addis, M.F.; Chassard, C.; Nader-Macias, M.E.F.; Grant, I.; Delbès, C.; Bogni, C.I.; Le Loir, Y.; Even, S. Milk Microbiota: What Are We Exactly Talking About? Front. Microbiol. 2020, 11, 60. [Google Scholar] [CrossRef] [PubMed]
- Porcellato, D.; Meisal, R.; Bombelli, A.; Narvhus, J.A. A core microbiota dominates a rich microbial diversity in the bovine udder and may indicate presence of dysbiosis. Sci. Rep. 2020, 10, 21608. [Google Scholar] [CrossRef] [PubMed]
- Ganda, E.K.; Bisinotto, R.S.; Lima, S.F.; Kronauer, K.; Decter, D.H.; Oikonomou, G.; Schukken, Y.H.; Bicalho, R.C. Longitudinal metagenomic profiling of bovine milk to assess the impact of intramammary treatment using a third-generation cephalosporin. Sci. Rep. 2016, 6, 37565. [Google Scholar] [CrossRef]
- Pikhtirova, A.; Pecka-Kiełb, E.; Zachwieja, A.; Bujok, J.; Zigo, F. Effects of non- aureus staphylococci on colostrum composition, properties and fatty acid profile in cow—A preliminary study. Pol. J. Vet. Sci. 2022, 25, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Berry, E.A.; Hillerton, J.E. The Effect of Selective Dry Cow Treatment on New Intramammary Infections. J. Dairy Sci. 2002, 85, 112–121. [Google Scholar] [CrossRef]
- Crispie, F.; Flynn, J.; Ross, R.P.; Hill, C.; Meaney, W.J. Dry cow therapy with a non-antibiotic intramammary teat seal—A review. Ir. Vet. J. 2004, 57, 412. [Google Scholar] [CrossRef]
- More, S.J.; Clegg, T.A.; McCoy, F. The use of national-level data to describe trends in intramammary antimicrobial usage on Irish dairy farms from 2003 to 2015. J. Dairy Sci. 2017, 100, 6400–6413. [Google Scholar] [CrossRef]
- Sneeringer, S.; MacDonald, J.; Key, N.; McBride, W.; Mathews, K. Economics of Antibiotic Use in U.S. Livestock Production; ERR-200; U.S. Department of Agriculture, Economic Research Service: Washington, DC, USA, 2015. [Google Scholar]
- Lam, T.J.G.M.; van den Borne, B.H.P.; Jansen, J.; Huijps, K.; van Veersen, J.C.L.; van Schaik, G.; Hogeveen, H. Improving bovine udder health: A national mastitis control program in the Netherlands. J. Dairy Sci. 2013, 96, 1301–1311. [Google Scholar] [CrossRef] [PubMed]
- USDA (US Department of Agriculture). Dairy 2007. Part III: Reference of Dairy Cattle Health and Management Practices in the United States, 2007. #N482.0908. USDA-Animal and Plant Health Inspection Service-Veterinary Services, Centers for Epidemiology and Animal Health (USDA-APHIS-VS, CEAH); National Animal Health Monitoring System: Fort Collins, CO, USA, 2008. [Google Scholar]
- ESVAC. 2020. Available online: https://www.ema.europa.eu/en/veterinary-regulatory/overview/antimicrobial-resistance/european-surveillance-veterinary-antimicrobial-consumption-esvac (accessed on 20 August 2022).
- EFSA Panel on Biological Hazards (BIOHAZ); Ricci, A.; Allende, A.; Bolton, D.; Chemaly, M.; Davies, R.; Fernández Escámez, P.S.; Girones, R.; Koutsoumanis, K.; Lindqvist, R.; et al. Risk for the development of Antimicrobial Resistance (AMR) due to feeding of calves with milk containing residues of antibiotics. EFSA J. 2017, 15, e04665. [Google Scholar] [CrossRef]
- Sachi, S.; Ferdous, J.; Sikder, M.; Hussani, S. Antibiotic residues in milk: Past, present, and future. J. Adv. Vet. Anim. Res. 2019, 6, 315. [Google Scholar] [CrossRef]
- Kurjogi, M.; Issa Mohammad, Y.H.; Alghamdi, S.; Abdelrahman, M.; Satapute, P.; Jogaiah, S. Detection and determination of stability of the antibiotic residues in cow’s milk. PLoS ONE 2019, 14, e0223475. [Google Scholar] [CrossRef] [PubMed]
- Layada, S.; Benouareth, D.-E.; Coucke, W.; Andjelkovic, M. Assessment of antibiotic residues in commercial and farm milk collected in the region of Guelma (Algeria). Int. J. Food Contam. 2016, 3, 19. [Google Scholar] [CrossRef]
- Andersson, D.I.; Hughes, D. Evolution of antibiotic resistance at non-lethal drug concentrations. Drug Resist. Updates 2012, 15, 162–172. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef]
- Liu, J.; Taft, D.H.; Maldonado-Gomez, M.X.; Johnson, D.; Treiber, M.L.; Lemay, D.G.; De Peters, E.J.; Mills, D.A. The fecal resistome of dairy cattle is associated with diet during nursing. Nat. Commun. 2019, 10, 4406. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.V.V.; Carroll, L.M.; Lima, S.; Foditsch, C.; Siler, J.D.; Bicalho, R.C.; Warnick, L.D. Impacts of feeding preweaned calves milk containing drug residues on the functional profile of the fecal microbiota. Sci. Rep. 2018, 8, 554. [Google Scholar] [CrossRef]
- Maynou, G.; Bach, A.; Terré; M. Feeding of waste milk to Holstein calves affects antimicrobial resistance of Escherichia coli and Pasteurella multocida isolated from fecal and nasal swabs. J. Dairy Sci. 2017, 100, 2682–2694. [Google Scholar] [CrossRef]
- Maynou, G.; Migura-Garcia, L.; Subirats, J.; Chester-Jones, H.; Ziegler, D.; Bach, A.; Terré, M. 1232 Impact of milk-feeding programs on fecal bacteria population and antimicrobial resistance genes in Escherichia coli isolated from feces in preweaned calves. J. Anim. Sci. 2016, 94 (Suppl. S5), 593. [Google Scholar] [CrossRef]
- Van Vleck Pereira, R.; Lima, S.; Siler, J.D.; Foditsch, C.; Warnick, L.D.; Bicalho, R.C. Ingestion of Milk Containing Very Low Concentration of Antimicrobials: Longitudinal Effect on Fecal Microbiota Composition in Preweaned Calves. PLoS ONE 2016, 11, e0147525. [Google Scholar] [CrossRef] [PubMed]
- Di Rocco, M.; Scollard, J.; Sayers, R.; Furey, A.; Danaher, M.; Jordan, K.; Lourenco, A. Migration of Cefquinome Antibiotic Residues from Milk to Dairy Products. Dairy 2021, 2, 658–670. [Google Scholar] [CrossRef]
- Zaheer, R.; Lakin, S.M.; Polo, R.O.; Cook, S.R.; Larney, F.J.; Morley, P.S.; Booker, C.W.; Hannon, S.J.; Van Domselaar, G.; Read, R.R.; et al. Comparative diversity of microbiomes and Resistomes in beef feedlots, downstream environments and urban sewage influent. BMC Microbiol. 2019, 19, 197. [Google Scholar] [CrossRef]
- Astorga, F.; Navarrete-Talloni, M.J.; Miró, M.P.; Bravo, V.; Toro, M.; Blondel, C.J.; Hervé-Claude, L.P. Antimicrobial resistance in E. coli isolated from dairy calves and bedding material. Heliyon 2019, 5, e02773. [Google Scholar] [CrossRef] [PubMed]
- Mungai, E.A.; Behravesh, C.B.; Gould, L.H. Increased Outbreaks Associated with Nonpasteurized Milk, United States, 2007–2012. Emerg. Infect. Dis. 2015, 21, 119–122. [Google Scholar] [CrossRef]
- Kyselková, M.; Jirout, J.; Vrchotová, N.; Schmitt, H.; Elhottová, D. Spread of tetracycline resistance genes at a conventional dairy farm. Front. Microbiol. 2015, 6, 536. [Google Scholar] [CrossRef]
- Oikonomou, G.; Bicalho, M.L.; Meira, E.; Rossi, R.E.; Foditsch, C.; Machado, V.S.; Teixeira, A.G.V.; Santisteban, C.; Schukken, Y.H.; Bicalho, R.C. Microbiota of Cow’s Milk; Distinguishing Healthy, Sub-Clinically and Clinically Diseased Quarters. PLoS ONE 2014, 9, e85904. [Google Scholar] [CrossRef]
- Zigo, F.; Farkašová, Z.; Výrostková, J.; Regecová, I.; Ondrašovičová, S.; Vargová, M.; Sasáková, N.; Pecka-Kielb, E.; Bursová, Š.; Kiss, D.S. Dairy Cows’ Udder Pathogens and Occurrence of Virulence Factors in Staphylococci. Animals 2022, 12, 470. [Google Scholar] [CrossRef]
- Boireau, C.; Cazeau, G.; Jarrige, N.; Calavas, D.; Madec, J.-Y.; Leblond, A.; Haenni, M.; Gay, É. Antimicrobial resistance in bacteria isolated from mastitis in dairy cattle in France, 2006–2016. J. Dairy Sci. 2018, 101, 9451–9462. [Google Scholar] [CrossRef] [PubMed]
- Saini, V.; McClure, J.T.; Léger, D.; Keefe, G.P.; Scholl, D.T.; Morck, D.W.; Barkema, H.W. Antimicrobial resistance profiles of common mastitis pathogens on Canadian dairy farms. J. Dairy Sci. 2012, 95, 4319–4332. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.T.; Marotz, C.; Washburne, A.; Silverman, J.; Zaramela, L.S.; Edlund, A.; Zengler, K.; Knight, R. Establishing microbial composition measurement standards with reference frames. Nat. Commun. 2019, 10, 2719. [Google Scholar] [CrossRef] [PubMed]
- Metzger, S.A.; Hernandez, L.L.; Skarlupka, J.H.; Suen, G.; Walker, T.M.; Ruegg, P.L. Influence of sampling technique and bedding type on the milk microbiota: Results of a pilot study. J. Dairy Sci. 2018, 101, 6346–6356. [Google Scholar] [CrossRef] [PubMed]
- Jovel, J.; Patterson, J.; Wang, W.; Hotte, N.; O’Keefe, S.; Mitchel, T.; Perry, T.; Kao, D.; Mason, A.L.; Madsen, K.L.; et al. Characterization of the Gut Microbiome Using 16S or Shotgun Metagenomics. Front. Microbiol. 2016, 7, 459. [Google Scholar] [CrossRef]
- Department of Health (2017). Ireland’s National Action Plan on Antimicrobial Resistance 2017–2020. (iNAP). Available online: http://health.gov.ie/national-patient-safety-office/patient-safety-surveillance/antimicrobialresistance-amr/ (accessed on 14 February 2020).
- Derakhshani, H.; Plaizier, J.C.; De Buck, J.; Barkema, H.W.; Khafipour, E. Composition of the teat canal and intramammary microbiota of dairy cows subjected to antimicrobial dry cow therapy and internal teat sealant. J. Dairy Sci. 2018a, 101, 10191–10205. [Google Scholar] [CrossRef] [PubMed]
- Hermansson, H.; Kumar, H.; Collado, M.C.; Salminen, S.; Isolauri, E.; Rautava, S. Breast Milk Microbiota Is Shaped by Mode of Delivery and Intrapartum Antibiotic Exposure. Front. Nutr. 2019, 6, 4. [Google Scholar] [CrossRef]
- Biscarini, F.; Cremonesi, P.; Castiglioni, B.; Stella, A.; Bronzo, V.; Locatelli, C.; Moroni, P. A Randomized Controlled Trial of Teat-Sealant and Antibiotic Dry-Cow Treatments for Mastitis Prevention Shows Similar Effect on the Healthy Milk Microbiome. Front. Vet. Sci. 2020, 7, 1–17. [Google Scholar] [CrossRef]
- Bonsaglia, E.C.R.; Gomes, M.S.; Canisso, I.F.; Zhou, Z.; Lima, S.F.; Rall, V.L.M.; Oikonomou, G.; Bicalho, R.C.; Lima, F.S. Milk microbiome and bacterial load following dry cow therapy without antibiotics in dairy cows with healthy mammary gland. Sci. Rep. 2017, 7, 8067. [Google Scholar] [CrossRef]
- Parker, K.I.; Compton, C.W.R.; Anniss, F.M.; Heuer, C.; McDougall, S. Quarter-Level Analysis of Subclinical and Clinical Mastitis in Primiparous Heifers Following the Use of a Teat Sealant or an Injectable Antibiotic, or Both, Precalving. J. Dairy Sci. 2008, 91, 169–181. [Google Scholar] [CrossRef]
- Porcellato, D.; Smistad, M.; Bombelli, A.; Abdelghani, A.; Jørgensen, H.J.; Skeie, S.B. Longitudinal Study of the Bulk Tank Milk Microbiota Reveals Major Temporal Shifts in Composition. Front. Microbiol. 2021, 12, 616429. [Google Scholar] [CrossRef]
- Rubiola, S.; Chiesa, F.; Dalmasso, A.; Di Ciccio, P.; Civera, T. Detection of Antimicrobial Resistance Genes in the Milk Production Environment: Impact of Host DNA and Sequencing Depth. Front. Microbiol. 2020, 11, 1983. [Google Scholar] [CrossRef] [PubMed]
- McHugh, A.J.; Feehily, C.; Fenelon, M.A.; Gleeson, D.; Hill, C.; Cotter, P.D. Tracking the Dairy Microbiota from Farm Bulk Tank to Skimmed Milk Powder. MSystems 2020, 5, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Huo, W.; Zhu, W.; Mao, S. Characterization of bacterial community of raw milk from dairy cows during subacute ruminal acidosis challenge by high-throughput sequencing. J. Sci. Food Agric. 2015, 95, 1072–1079. [Google Scholar] [CrossRef]
- Gulbe, G.; Valdovska, A. Microbiological content of cow milk depending upon season and herd size in latvian organic farms. Vet. Med. Res. Rural. Dev. 2014, 1, 161–168. [Google Scholar]
- Masoud, W.; Vogensen, F.K.; Lillevang, S.; Abu Al-Soud, W.; Sørensen, S.J.; Jakobsen, M. The fate of indigenous microbiota, starter cultures, Escherichia coli, Listeria innocua and Staphylococcus aureus in Danish raw milk and cheeses determined by pyrosequencing and quantitative real time (qRT)-PCR. Int. J. Food Microbiol. 2012, 153, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Watts, J. Chapter: Bovine Mastitis. In Diagnostic Procedure in Veterinary Bacteriology and Mycology; Academic Press Inc.: San Diego, CA, USA, 1990; p. 469. [Google Scholar] [CrossRef]
- Nickerson, S.C. MASTITIS PATHOGENS|Contagious Pathogens. In Encyclopedia of Dairy Sciences; Hubert Roginski, H., Ed.; Elsevier: Amsterdam, The Netherlands, 2002; pp. 1723–1728. ISBN 9780122272356. [Google Scholar] [CrossRef]
- Frétin, M.; Martin, B.; Rifa, E.; Isabelle, V.-M.; Pomiès, D.; Ferlay, A.; Montel, M.-C.; Delbès, C. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci. Rep. 2018, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Woudstra, S.; Lücken, A.; Wente, N.; Zhang, Y.; Leimbach, S.; Gussmann, M.K.; Kirkeby, C.; Krömker, V. Reservoirs of Corynebacterium spp. in the Environment of Dairy Cows. Pathogens 2023, 12, 139. [Google Scholar] [CrossRef]
- Lücken, A.; Wente, N.; Zhang, Y.; Woudstra, S.; Krömker, V. Corynebacteria in Bovine Quarter Milk Samples—Species and Somatic Cell Counts. Pathogens 2021, 10, 831. [Google Scholar] [CrossRef]
- Watts, J.L.; Lowery, D.E.; Teel, J.F.; Rossbach, S. Identification of Corynebacterium bovis and other Coryneforms Isolated from Bovine Mammary Glands. J. Dairy Sci. 2000, 83, 2373–2379. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, W.; Huang, J.; Luo, H.; Jin, Y.; Zhang, W.; Liu, Z.; Huang, Y. Actinoalloteichus hymeniacidonis sp. nov., an actinomycete isolated from the marine sponge Hymeniacidon perleve. Int. J. Syst. Evol. Microbiol. 2006, 10, 2309–2312. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Chen, J.; Wang, B.; Chen, T.; Chen, J.; Zhang, Y.; Liu, X.; Chen, Q. Activation and enhancement of caerulomycin A biosynthesis in marine-derived Actinoalloteichus sp. AHMU CJ021 by combinatorial genome mining strategies. Microb. Cell Factories 2020, 19, 159. [Google Scholar] [CrossRef] [PubMed]
- Boudjelal, F.; Zitouni, A.; Bouras, N.; Schumann, P.; Spröer, C.; Sabaou, N.; Klenk, H.-P. Actinoalloteichus hoggarensis sp. nov., an actinomycete isolated from Saharan soil. Int. J. Syst. Evol. Microbiol. 2015, 65 Pt 6, 2006–2010. [Google Scholar] [CrossRef] [PubMed]
- Schaffert, L.; Albersmeier, A.; Winkler, A.; Kalinowski, J.; Zotchev, S.B.; Rückert, C. Complete genome sequence of the actinomycete Actinoalloteichus hymeniacidonis type strain HPA 177T isolated from a marine sponge. Stand. Genom. Sci. 2016, 11, 91. [Google Scholar] [CrossRef] [PubMed]
- Lima, S.F.; Teixeira AG, V.; Lima, F.S.; Ganda, E.K.; Higgins, C.H.; Oikonomou, G.; Bicalho, R.C. The bovine colostrum microbiome and its association with clinical mastitis. J. Dairy Sci. 2017, 100, 3031–3042. [Google Scholar] [CrossRef] [PubMed]
- Doyle, C.J.; Gleeson, D.; O’Toole, P.W.; Cotter, P.D. Impacts of Seasonal Housing and Teat Preparation on Raw Milk Microbiota: A High-Throughput Sequencing Study. Appl. Environ. Microbiol. 2017, 83, e02694-16. [Google Scholar] [CrossRef] [PubMed]
- Derakhshani, H.; Plaizier, J.C.; De Buck, J.; Barkema, H.W.; Khafipour, E. Association of bovine major histocompatibility complex (BoLA) gene polymorphism with colostrum and milk microbiota of dairy cows during the first week of lactation. Microbiome 2018, 6, 203. [Google Scholar] [CrossRef] [PubMed]
- Blaustein, R.A.; Shelton, D.R.; Van Kessel JA, S.; Karns, J.S.; Stocker, M.D.; Pachepsky, Y.A. Irrigation waters and pipe-based biofilms as sources for antibiotic-resistant bacteria. Environ. Monit. Assess. 2016, 188, 56. [Google Scholar] [CrossRef]
- Li, X.; Watanabe, N.; Xiao, C.; Harter, T.; McCowan, B.; Liu, Y.; Atwill, E.R. Antibiotic-resistant E. coli in surface water and groundwater in dairy operations in Northern California. Environ. Monit. Assess. 2014, 186, 1253–1260. [Google Scholar] [CrossRef]
- Pan, M.; Chu, L.M. Leaching behavior of veterinary antibiotics in animal manure-applied soils. Sci. Total Environ. 2017, 579, 466–473. [Google Scholar] [CrossRef]
- Baker, M.; Williams, A.D.; Hooton, S.P.; Helliwell, R.; King, E.; Dodsworth, T.; Baena-Nogueras, R.M.; Warry, A.; Ortori, C.A.; Todman, H.; et al. Antimicrobial resistance in dairy slurry tanks: A critical point for measurement and control. bioRxiv 2022. [Google Scholar] [CrossRef]
- Pitta, D.W.; Indugu, N.; Toth, J.D.; Bender, J.S.; Baker, L.D.; Hennessy, M.L.; Vecchiarelli, B.; Aceto, H.; Dou, Z. The distribution of microbiomes and resistomes across farm environments in conventional and organic dairy herds in Pennsylvania. Environ. Microbiome 2020, 15, 21. [Google Scholar] [CrossRef] [PubMed]
- Sawant, A.A.; Hegde, N.V.; Straley, B.A.; Donaldson, S.C.; Love, B.C.; Knabel, S.J.; Jayarao, B.M. Antimicrobial-Resistant Enteric Bacteria from Dairy Cattle. Appl. Environ. Microbiol. 2007, 73, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Adkins, P.R.F.; Middleton, J.R. Laboratory Handbook on Bovine Mastitis; National Mastitis Council: New Prague, MN, USA, 2017. [Google Scholar]
- Rinke, C.; Low, S.; Woodcroft, B.J.; Raina, J.-B.; Skarshewski, A.; Le, X.H.; Butler, M.K.; Stocker, R.; Seymour, J.; Tyson, G.W.; et al. Validation of picogram- and femtogram-input DNA libraries for microscale metagenomics. PeerJ 2016, 4, e2486. [Google Scholar] [CrossRef] [PubMed]
- McArthur, A.G.; Waglechner, N.; Nizam, F.; Yan, A.; Azad, M.A.; Baylay, A.J.; Bhullar, K.; Canova, M.J.; De Pascale, G.; Ejim, L.; et al. The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 2013, 57, 3348–3357. [Google Scholar] [CrossRef] [PubMed]
- Calgaro, M.; Romualdi, C.; Waldron, L.; Risso, D.; Vitulo, N. Assessment of statistical methods from single cell, bulk RNA-seq, and metagenomics applied to microbiome data. Genome Biol. 2020, 21, 191. [Google Scholar] [CrossRef]
- McMurdie, J.P.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Patangia, D.V.; Grimaud, G.; Linehan, K.; Ross, R.P.; Stanton, C. Microbiota and Resistome Analysis of Colostrum and Milk from Dairy Cows Treated with and without Dry Cow Therapies. Antibiotics 2023, 12, 1315. https://doi.org/10.3390/antibiotics12081315
Patangia DV, Grimaud G, Linehan K, Ross RP, Stanton C. Microbiota and Resistome Analysis of Colostrum and Milk from Dairy Cows Treated with and without Dry Cow Therapies. Antibiotics. 2023; 12(8):1315. https://doi.org/10.3390/antibiotics12081315
Chicago/Turabian StylePatangia, Dhrati V., Ghjuvan Grimaud, Kevin Linehan, R. Paul Ross, and Catherine Stanton. 2023. "Microbiota and Resistome Analysis of Colostrum and Milk from Dairy Cows Treated with and without Dry Cow Therapies" Antibiotics 12, no. 8: 1315. https://doi.org/10.3390/antibiotics12081315
APA StylePatangia, D. V., Grimaud, G., Linehan, K., Ross, R. P., & Stanton, C. (2023). Microbiota and Resistome Analysis of Colostrum and Milk from Dairy Cows Treated with and without Dry Cow Therapies. Antibiotics, 12(8), 1315. https://doi.org/10.3390/antibiotics12081315