
Chemical Analysis and Investigation of Antimicrobial and Antibiofilm Activities of Prangos trifida (Apiaceae)

 ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Chemical Composition of P. trifida
2.1.1. Composition of Essential Oils
2.1.2. Composition of Headspace Volatiles
2.1.3. Fatty Acid, Phytosterol and Triterpene Composition of CH2Cl2 Extracts
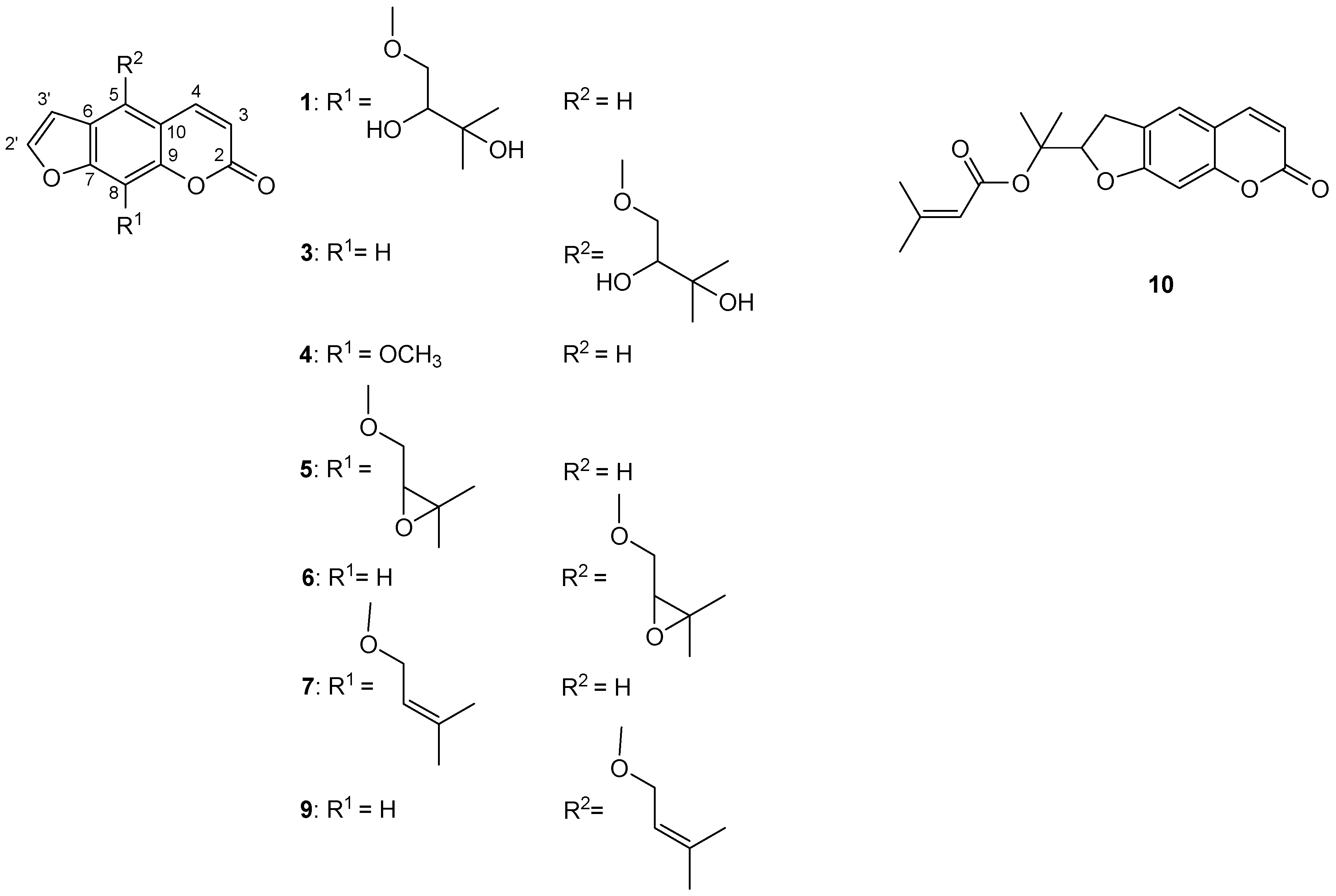
2.1.4. Coumarin Composition of CH2Cl2 Extracts and Their MeOH Fractions (MFDEs)
2.2. Antimicrobial and Antibiofilm Activities of P. trifida
2.2.1. Antimicrobial Activity of Essential Oils
2.2.2. Antibiofilm Activity of Essential Oils
2.2.3. Antimicrobial Activity of MFDEs
2.2.4. Antibiofilm Activity of MFDEs
2.2.5. Testing MFDEs in Congo Red and Ergosterol Binding Assays in Candida albicans
3. Discussion
3.1. Chemical Composition of P. trifida
3.1.1. Composition of Volatile Constituents
3.1.2. Chemical Composition of CH2Cl2 Extracts
3.2. Antimicrobial and Antibiofilm Activities, and Potential Mechanisms of Selected P. trifida Isolated Products
3.2.1. Antimicrobial and Antibiofilm Activities of Essential Oils
3.2.2. Antimicrobial and Antibiofilm Activities of MFDEs
3.2.3. Potential Mechanisms of Action against Candida albicans of MFDEs
4. Materials and Methods
4.1. Plant Material
4.2. Isolation of the Essential Oils
4.3. Chemical Analysis of the Essential Oils
4.4. Static Headspace (HS) Extraction and Chemical Analysis of HS Volatiles
4.5. Obtaining of CH2Cl2 Extracts
4.6. Saponification and Transesterification of CH2Cl2 Extracts
4.7. Analysis of the FAME, Phytosterols and Triterpenes in CH2Cl2 Extracts
4.8. Obtaining of Oxypeucedanin (6) and Prantschimgin (10) from CH2Cl2 Extracts
4.9. Obtaining of Methanol Fractions of CH2Cl2 Extracts (MFDEs)
4.10. LC–MS Analysis of Coumarins in MFDEs
4.11. Investigation of Antibacterial and Antifungal Activities of Essential Oils and MFDEs
4.12. Inhibition of Biofilm Formation of Essential Oils and MFDEs
4.13. Congo Red Binding Assay
4.14. Ergosterol Binding Assay in C. albicans
4.15. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stojković, D.; Petrović, J.; Carević, T.; Soković, M.; Liaras, K. Synthetic and Semisynthetic Compounds as Antibacterials Targeting Virulence Traits in Resistant Strains: A Narrative Updated Review. Antibiotics 2023, 12, 963. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Ćirić, A.; Stojković, D. Emerging Antifungal Targets and Strategies. Int. J. Mol. Sci. 2022, 23, 2756. [Google Scholar] [CrossRef] [PubMed]
- Chassagne, F.; Samarakoon, T.; Porras, G.; Lyles, J.T.; Dettweiler, M.; Marquez, L.; Salam, A.M.; Shabih, S.; Farrokhi, D.R.; Quave, C.L. A Systematic Review of Plants With Antibacterial Activities: A Taxonomic and Phylogenetic Perspective. Front. Pharmacol. 2021, 11, 586548. [Google Scholar] [CrossRef] [PubMed]
- Kostić, M.; Ivanov, M.; Markovic, T.; Sanković Babić, S.; Barros, L.; Calhelha, R.; Sokovic, M.; Ciric, A. An in vitro study of the Origanum minutiflorum O. Schwarz & P. H. Davis and Coriandrum sativum L. essential oils as chronic tonsillitis therapeutics: Antibacterial, antibiofilm, antioxidant, and cytotoxic activities. J. Essent. Oil Res. 2022, 34, 533–543. [Google Scholar] [CrossRef]
- Plants of the World Online. Available online: https://powo.science.kew.org (accessed on 14 December 2023).
- Mottaghipisheh, J.; Kiss, T.; Tóth, B.; Csupor, D. The Prangos genus: A comprehensive review on traditional use, phytochemistry, and pharmacological activities. Phytochem. Rev. 2020, 19, 1449–1470. [Google Scholar] [CrossRef]
- Bruno, M.; Ilardi, V.; Lupidi, G.; Quassinti, L.; Bramucci, M.; Fiorini, D.; Venditti, A.; Maggi, F. The nonvolatile and volatile metabolites of Prangos ferulacea and their biological properties. Planta Med. 2019, 85, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Badalamenti, N.; Maresca, V.; Di Napoli, M.; Bruno, M.; Basile, A.; Zanfardino, A. Chemical composition and biological activities of Prangos ferulacea essential oils. Molecules 2022, 27, 7430. [Google Scholar] [CrossRef]
- Azarkish, P.; Moghaddam, M.; Ghasemi Pirbalouti, A.; Khakdan, F. Variability in the essential oil of different wild populations of Prangos platychlaena collected from Southwestern Iran. Plant Biosyst. 2021, 155, 1100–1110. [Google Scholar] [CrossRef]
- Zengin, G.; Mahomoodally, M.F.; Yıldıztugay, E.; Jugreet, S.; Khan, S.U.; Dall’Acqua, S.; Mollica, A.; Bouyahya, A.; Montesano, D. Chemical composition, biological activities and in silico analysis of essential oils of three endemic Prangos species from Turkey. Molecules 2022, 27, 1676. [Google Scholar] [CrossRef]
- Karahisar, E.; Köse, Y.B.; İşcan, G.; Kurkcuoglu, M.; Tugay, O. Chemical Composition and Anticandidal Activity of Essential Oils Obtained from Different Parts of Prangos heyniae H. Duman & MF Watson. Rec. Nat. Prod. 2022, 16, 74–83. [Google Scholar] [CrossRef]
- Sevin, G.; Alan, E.; Demir, S.; Albayrak, G.; Demiroz, T.; Yetik-Anacak, G.; Baykan, S. Comparative evaluation of relaxant effects of three Prangos species on mouse corpus cavernosum: Chemical characterization and the relaxant mechanisms of action of P. pabularia and (+)-oxypeucedanin. J. Ethnopharmacol. 2022, 284, 114823. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.; Yazıcı-Tütüniş, S.; Bilgin, M.; Tan, E.; Miski, M. Antibacterial Activities of Pyrenylated Coumarins from the Roots of Prangos hulusii. Molecules 2017, 22, 1098. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Sinan, K.I.; Ak, G.; Mahomoodally, M.F.; Paksoy, M.Y.; Picot-Allain, C.; Glamočlija, J.; Soković, M.; Jekő, J.; Cziáky, Z.; et al. Chemical profile, antioxidant, antimicrobial, enzyme inhibitory, and cytotoxicity of seven Apiaceae species from Turkey: A comparative study. Ind. Crops Prod. 2020, 153, 112572. [Google Scholar] [CrossRef]
- Dall’Acqua, S.; Sut, S.; Zengin, G.; Peron, G.; Elbasan, F.; Yildiztugay, E.; Bibi Sadeer, N.; Mahomoodally, M.F. Phytochemical screening, antioxidant, and enzyme inhibitory properties of three Prangos species (P. heyniae, P. meliocarpoides var. meliocarpoides, and P. uechtritzii) depicted by comprehensive LC-MS and multivariate data analysis. Antioxidants 2022, 11, 1712. [Google Scholar] [CrossRef] [PubMed]
- Nosrati, M.; Ranjbar, R. Investigation of the antibacterial and biofilm inhibitory activities of Prangos acaulis (DC.) Bornm in nanoparticulated formulation. Nanotechnology 2022, 33, 385103. [Google Scholar] [CrossRef] [PubMed]
- Sarghaleh, S.J.; Behbahani, B.A.; Hojjati, M.; Vasiee, A.; Noshad, M. Evaluation of the constituent compounds, antioxidant, anticancer, and antimicrobial potential of Prangos ferulacea plant extract and its effect on Listeria monocytogenes virulence gene expression. Front. Microbiol. 2023, 14, 1202228. [Google Scholar] [CrossRef]
- Herrnstadt, I.; Heyn, C.C. A monographic study of the genus Prangos (Umbelliferae). Boissiera 1977, 26, 1–91. [Google Scholar]
- Niketić, M. Cachrys alpina L. In The Red Data Book of Flora of Serbia. Extinct and Critically Endangered Taxa; Stevanović, V., Ed.; Ministry of Environment of the Republic of Serbia, Faculty of Biology, University of Belgrade, Institution for Protection of Nature of the Republic of Serbia: Beograd, Serbia, 1999; pp. 174–176+454–455, (In Serbian and English). [Google Scholar]
- Hand, R. Apiaceae. In Euro+Med Plantbase—The Information Resource for Euro-Mediterranean Plant Diversity; Dipartimento di Scienzeambientali e Biodiversità ed Orto botanico, Università degli Studi di Palermo: Palermo, Italy, 2011; Available online: https://europlusmed.org/cdm_dataportal/taxon/368c1360-c3ad-41c6-8a61-e46168b44d47#footnote-T (accessed on 10 September 2023).
- Lyskov, D.F. Systematics of the Genus Prangos (Umbelliferae, Apioideae) and Related Taxa: Comparison of Morphological-Anatomic and Molecular Data. Ph.D. Thesis, Faculty of Biology, M.V. Lomonosov Moscow State University, Moscow, Russia, 2015. (In Russian). [Google Scholar]
- Lyskov, D.; Samigullin, T. European Prangos species complexes: When classic morphological features are not enough to distinguish similar species. In Abstract Book, Proceedings of the IX Apiales Symposium, The Gold Coast Marina Club, Guangzhou, China, 31 July–2 August 2017; Oskolski, A., Nuraliev, M., Tilney, P., Eds.; University of Johannesburg: Johannesburg, South Africa; Komarov Botanical Institute: St. Petersburg, Russia; Lomonosov Moscow State University: Moscow, Russia, 2017; p. 23. [Google Scholar]
- Baser, K.H.C.; Demirci, B.; Akalin, E.; Özhatay, N. Composition of the Essential Oil of Cachrys alpine Bieb. J. Essent. Oil Res. 2004, 16, 167–168. [Google Scholar] [CrossRef]
- Palá-Paül, J.; Velasco-Negueruela, A.; Pérez-Alonso, J.; Maqueda, J.; Sanz, J. Volatile oil Constituents from Different Parts of Cachrys trifida L. J. Essent. Oil Res. 2004, 16, 347–349. [Google Scholar] [CrossRef]
- Korotkov, O.I.; Shevchuk, O.M.; Shatko, V.G.; Timashova, L.A.; Feskov, S.A. Some biochemical characteristics of Prangos trifida (Mill.) Herrnst. & Heyn. Bull. State Nikit. Botan. Gard. 2018, 76–83. [Google Scholar] [CrossRef]
- Di Napoli, M.; Castagliuolo, G.; Badalamenti, N.; Vaglica, A.; Ilardi, V.; Varcamonti, M.; Bruno, M.; Zanfardino, A. Chemical composition, antimicrobial and antioxidant activities of the essential oil of Italian Prangos trifida (Mill.) Herrnst. & Heyn. Nat. Prod. Res. 2022, 37, 3772–3786. [Google Scholar] [CrossRef]
- Maresca, V.; Badalamenti, N.; Ilardi, V.; Bruno, M.; Basile, A. The Antioxidant Properties and Protective Capacity of Prangos trifida and Cachrys cristata Essential Oils against Cd Stress in Lunularia cruciata and Brassica napus. Antioxidants 2023, 12, 793. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; De Las Heras, B.; Silván, A.M.; Pascual, R.; Bermejo, P.; Rodriguez, B.; Villar, A.M. Effects of furocoumarins from Cachrys trifida on some macrophage functions. J. Pharm. Pharmacol. 2001, 53, 1163–1168. [Google Scholar] [CrossRef] [PubMed]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4.1 ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2017. [Google Scholar]
- Khalighi-Sigaroodi, F.; Hadjiakhoondi, A.; Shafiee, A.; Mozaffarian, V.A.; Shahverdi, A.R.; Alavi, S.H. Phytochemical analysis of Ferulogo bernardii Tomk & M. Pimen. DARU J. Pharm. Sci. 2006, 14, 214–221. [Google Scholar]
- Wei, Y.; Ito, Y. Preparative isolation of imperatorin, oxypeucedanin and isoimperatorin from traditional Chinese herb “bai zhi” Angelica dahurica (Fisch. ex Hoffm) Benth. et Hook using multidimensional high-speed counter-current chromatography. J. Chromatogr. A 2006, 1115, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Sajjadi, S.E.; Jamali, M.; Shokoohinia, Y.; Abdi, G.; Shahbazi, B.; Fattahi, A. Antiproliferative evaluation of terpenoids and terpenoid coumarins from Ferulago macrocarpa (Fenzl) Boiss. fruits. Pharmacogn. Res. 2015, 7, 322–328. [Google Scholar] [CrossRef]
- Ušjak, L.J.; Drobac, M.M.; Niketić, M.S.; Petrović, S.D. Chemosystematic Significance of Essential Oil Constituents and Furanocoumarins of Underground Parts and Fruits of Nine Heracleum L. Taxa from Southeastern Europe. Chem. Biodivers. 2018, 15, e1800412. [Google Scholar] [CrossRef] [PubMed]
- Kiyonga, A.N.; Hong, G.; Kim, H.S.; Suh, Y.G.; Jung, K. Facile and Rapid Isolation of Oxypeucedanin Hydrate and Byakangelicin from Angelica dahurica by Using [Bmim] Tf2N Ionic Liquid. Molecules 2021, 26, 830. [Google Scholar] [CrossRef]
- Frérot, E.; Decorzant, E. Quantification of Total Furocoumarins in Citrus Oils by HPLC Coupled with UV, Fluorescence, and Mass Detection. J. Agric. Food Chem. 2004, 52, 6879–6886. [Google Scholar] [CrossRef]
- Dewick, P.M. Medicinal Natural Products: A Biosynthetic Approach, 3rd ed.; John Wiley & Sons: Chichester, UK, 2009; pp. 161–165. [Google Scholar]
- Kusano, M.; Kobayashi, M.; Iizuka, Y.; Fukushima, A.; Saito, K. Unbiased profiling of volatile organic compounds in the headspace of Allium plants using an in-tube extraction device. BMC Res. Notes 2016, 9, 133. [Google Scholar] [CrossRef]
- Skalicka-Woźniak, K.; Głowniak, K. Pressurized liquid extraction of coumarins from fruits of Heracleum leskowii with application of solvents with different polarity under increasing temperature. Molecules 2012, 17, 4133–4141. [Google Scholar] [CrossRef]
- Bagci, E. Fatty acids and tocochromanol patterns of some Turkish Apiaceae (Umbelliferae) plants; a chemotaxonomic approach. Acta Bot. Gallica 2007, 154, 143–151. [Google Scholar] [CrossRef]
- Küçükboyacι, N.; Ayaz, F.; Adιgüzel, N.; Bani, B.; Gören, A.C. Fatty Acid Methyl Ester Composition of Some Turkish Apiaceae Seed Oils: New Sources for Petroselinic Acid. Nat. Prod. Commun. 2016, 11, 1934578X1601101118. [Google Scholar] [CrossRef]
- Saini, R.K.; Song, M.H.; Yu, J.W.; Shang, X.; Keum, Y.S. Phytosterol Profiling of Apiaceae Family Seeds Spices Using GC-MS. Foods 2021, 10, 2378. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, G.A.; Belenovskaya, L.M. Prantschimgin—A new coumarin from the roots of Prangos tschimganica. Chem. Nat. Compd. 1966, 2, 190–192. [Google Scholar] [CrossRef]
- Basilico, M.Z.; Basilico, J.C. Inhibitory effects of some spice essential oils on Aspergillus ochraceus NRRL 3174 growth and ochratoxin A production. Lett. Appl. Microbiol. 1999, 29, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Bayman, P.; Baker, J.L.; Doster, M.A.; Michailides, T.J.; Mahoney, N.E. Ochratoxin production by the Aspergillus ochraceus group and Aspergillus alliaceus. Appl. Environ. Microbiol. 2002, 68, 2326–2329. [Google Scholar] [CrossRef] [PubMed]
- Mazaheri, T.; Cervantes-Huamán, B.R.; Bermúdez-Capdevila, M.; Ripolles-Avila, C.; Rodríguez-Jerez, J.J. Listeria monocytogenes biofilms in the food industry: Is the current hygiene program sufficient to combat the persistence of the pathogen? Microorganisms 2021, 9, 181. [Google Scholar] [CrossRef]
- Zhao, L.; Poh, C.N.; Wu, J.; Zhao, X.; He, Y.; Yang, H. Effects of electrolysed water combined with ultrasound on inactivation kinetics and metabolite profiles of Escherichia coli biofilms on food contact surface. Innov. Food Sci. Emerg. Technol. 2022, 76, 102917. [Google Scholar] [CrossRef]
- Idrees, M.; Sawant, S.; Karodia, N.; Rahman, A. Staphylococcus aureus biofilm: Morphology, genetics, pathogenesis and treatment strategies. Int. J. Environ. Res. Public Health 2021, 18, 7602. [Google Scholar] [CrossRef]
- Villa, K.; Sopha, H.; Zelenka, J.; Motola, M.; Dekanovsky, L.; Beketova, D.C.; Macak, J.M.; Ruml, T.; Pumera, M. Enzyme-photocatalyst tandem microrobot powered by urea for Escherichia coli biofilm eradication. Small 2022, 18, 2106612. [Google Scholar] [CrossRef]
- Rios, J.L.; Recio, M.C. Medicinal plants and antimicrobial activity. J. Ethnopharmacol. 2005, 100, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Miladi, H.; Zmantar, T.; Kouidhi, B.; Al Qurashi, Y.M.; Bakhrouf, A.; Chaabouni, Y.; Mahdouani, K.; Chaieb, K. Synergistic effect of eugenol, carvacrol, thymol, p-cymene and γ-terpinene on inhibition of drug resistance and biofilm formation of oral bacteria. Microb. Pathog. 2017, 112, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Reichling, J. Anti-biofilm and Virulence Factor-Reducing Activities of Essential Oils and Oil Components as a Possible Option for Bacterial Infection Control. Planta Med. 2020, 86, 520–537. [Google Scholar] [CrossRef] [PubMed]
- Purkait, S.; Bhattacharya, A.; Bag, A.; Chattopadhyay, R.R. Evaluation of antibiofilm efficacy of essential oil components β-caryophyllene, cinnamaldehyde and eugenol alone and in combination against biofilm formation and preformed biofilms of Listeria monocytogenes and Salmonella typhimurium. Lett. Appl. Microbiol. 2020, 71, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kerekes, E.B.; Deák, É.; Takó, M.; Tserennadmid, R.; Petkovits, T.; Vágvölgyi, C.; Krisch, J. Anti-biofilm forming and anti-quorum sensing activity of selected essential oils and their main components on food-related micro-organisms. J. Appl. Microbiol. 2013, 115, 933–942. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Tabassum, N.; Jeong, G.J.; Jung, W.K.; Kim, Y.M. Inhibition of Mixed Biofilms of Candida albicans and Staphylococcus aureus by β-Caryophyllene-Gold Nanoparticles. Antibiotics 2023, 12, 726. [Google Scholar] [CrossRef]
- Khoury, M.; El Beyrouthy, M.; Eparvier, V.; Ouaini, N.; Stien, D. Chemical diversity and antimicrobial activity of the essential oils of four Apiaceae species growing wild in Lebanon. J. Essent. Oil Res. 2018, 30, 25–31. [Google Scholar] [CrossRef]
- Yousefi, K.; Hamedeyazdan, S.; Hodaei, D.; Lotfipour, F.; Baradaran, B.; Orangi, M.; Fathiazad, F. An in vitro ethnopharmacological study on Prangos ferulacea: A wound healing agent. Bioimpacts 2017, 7, 75–82. [Google Scholar] [CrossRef]
- Özek, G.; Özek, T.; İşcan, G.; Başer, K.H.C.; Hamzaoglu, E.; Duran, A. Comparison of hydrodistillation and microdistillation methods for the analysis of fruit volatiles of Prangos pabularia Lindl., and evaluation of its antimicrobial activity. S. Afr. J. Bot. 2007, 73, 563–569. [Google Scholar] [CrossRef]
- Brusotti, G.; Ibrahim, M.F.; Dentamaro, A.; Gilardoni, G.; Tosi, S.; Grisoli, P.; Dacarro, C.; Guglielminetti, M.L.; Hussain, F.H.S.; Caccialanza, G.; et al. Chemical composition and antimicrobial activity of the volatile fractions from leaves and flowers of the wild Iraqi Kurdish plant Prangos peucedanifolia Fenzl. Chem. Biodivers. 2013, 10, 274–280. [Google Scholar] [CrossRef]
- Uzel, A.; Dirmenci, T.; Çelik, A.; Arabaci, T. Composition and antimicrobial activity of Prangos platychlaena and P. uechtritzii. Chem. Nat. Compd. 2006, 42, 169–171. [Google Scholar] [CrossRef]
- Latgé, J.P.; Chamilos, G. Aspergillus fumigatus and Aspergillosis in 2019. Clin. Microbiol. Rev. 2019, 33, 10–1128. [Google Scholar] [CrossRef] [PubMed]
- Karakaya, S.; Şimşek, D.; Özbek, H.; Güvenalp, Z.; Altanlar, N.; Kazaz, C.; Kiliç, C.S. Antimicrobial Activities of Extracts and Isolated Coumarins from the Roots of Four Ferulago Species Growing in Turkey. Iran. J. Pharm. Res. 2019, 18, 1516–1529. [Google Scholar] [CrossRef] [PubMed]
- Ulubelen, A.; Topcu, G.; Tan, N.; Ölçal, S.; Johansson, C.; Üçer, M.; Birman, H.; Tamer, Ş. Biological activities of a Turkish medicinal plant, Prangos platychlaena. J. Ethnopharmacol. 1995, 45, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Mileski, K.S.; Trifunović, S.S.; Ćirić, A.D.; Šakić, Z.M.; Ristić, M.S.; Todorović, N.M.; Matevski, V.S.; Marin, P.D.; Tešević, V.V.; Džamić, A.M. Research on Chemical Composition and Biological Properties Including Antiquorum Sensing Activity of Angelica pancicii Vandas Aerial Parts and Roots. J. Agric. Food Chem. 2017, 65, 10933–10949. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.C.A.; De Brito Bezerra, A.P.; De Sousa, J.P.; De Oliveira Lima, E. Investigating the antifungal activity and mechanism(s) of geraniol against Candida albicans strains. Med. Mycol. 2015, 53, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Sudha, K.G.; Thirumalaivasan, N.; Ahamed, M.; Pandiaraj, S.; Rajeswari, V.D.; Vinayagam, Y.; Thiruvengadam, M.; Govindasamy, R. Green Synthesis of Magnesium Oxide Nanoparticles by Using Abrus precatorius Bark Extract and Their Photocatalytic, Antioxidant, Antibacterial, and Cytotoxicity Activities. Bioengineering 2023, 10, 302. [Google Scholar] [CrossRef]
- Gayathiri, E.; Prakash, P.; Kumaravel, P.; Jayaprakash, J.; Ragunathan, M.G.; Sankar, S.; Pandiaraj, S.; Thirumalaivasan, N.; Thiruvengadam, M.; Govindasamy, R. Computational approaches for modeling and structural design of biological systems: A comprehensive review. Prog. Biophys. Mol. Biol. 2023, 185, 17–32. [Google Scholar] [CrossRef]
- Pemán, J.; Cantón, E.; Espinel-Ingroff, A. Antifungal drug resistance mechanisms. Expert Rev. Anti-Infect. Ther. 2009, 7, 453–460. [Google Scholar] [CrossRef]
- EDQM. European Pharmacopoeia 11.0; European Directorate for the Quality of Medicine & Health Care of the Council of Europe (EDQM): Strasbourg, France, 2023. [Google Scholar]
- Arsenijević, J.; Marković, J.; Šoštarić, I.; Ražić, S. A chemometrics as a powerful tool in the elucidation of the role of metals in the biosynthesis of volatile organic compounds in Hungarian thyme samples. Plant Physiol. Biochem. 2013, 71, 298–306. [Google Scholar] [CrossRef]
- Ušjak, L.; Sofrenić, I.; Tešević, V.; Drobac, M.; Niketić, M.; Petrović, S. Fatty acids, sterols, and triterpenes of the fruits of 8 Heracleum taxa. Nat. Prod. Commun. 2019, 14, 1934578X19856788. [Google Scholar] [CrossRef]
- Tang, J.J.; Zhao, N.; Gao, Y.Q.; Han, R.; Wang, X.Y.; Tian, J.M.; Gao, J.M. Phytosterol profiles and iridoids of the edible Eucommia ulmoides Oliver seeds and their anti-inflammatory potential. Food Biosci. 2021, 43, 101295. [Google Scholar] [CrossRef]
- Kostić, M.; Smiljković, M.; Petrović, J.; Glamočlija, J.; Barros, L.; Ferreira, I.C.F.R.; Ćirić, A.; Soković, M. Chemical, nutritive composition and a wide range of bioactive properties of honey mushroom: Armillaria mellea (Vahl: Fr.) Kummer. Food Funct. 2017, 8, 3239–3249. [Google Scholar] [CrossRef] [PubMed]
- Ivanov, M.; Kostić, M.; Stojković, D.; Soković, M. Rosmarinic acid–Modes of antimicrobial and antibiofilm activities of common plant polyphenol. S. Afr. J. Bot. 2022, 146, 521–527. [Google Scholar] [CrossRef]
- Ivanov, M.; Gašić, U.; Stojković, D.; Kostić, M.; Mišić, D.; Soković, M. New Evidence for Artemisia absinthium L. Application in Gastrointestinal Ailments: Ethnopharmacology, Antimicrobial Capacity, Cytotoxicity, and Phenolic Profile. Evid. Based Complement. Alternat. Med. 2021, 2021, 9961089. [Google Scholar] [CrossRef]


{kind=link}
| RIexp 1 | RIlit 2 | Compound 3 | Essential Oils, % 4 | Headspace Volatiles, % 4 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Root | Leaf | Stem | Fruit | Root | Leaf | Stem | Fruit | |||
| 934 | 924 | α-Thujene | - | - | - | - | tr | tr | tr | 0.4 |
| 935 | 932 | α-Pinene | 1.3 | 11.3 | 5.7 | 7.0 | 5.0 | 17.3 | 21.7 | 19.5 |
| 950 | 946 | Camphene | tr | 0.5 | 0.9 | tr | tr | 0.6 | 1.1 | tr |
| 975 | 969 | Sabinene | tr | 0.4 | tr | 4.5 | tr | 0.5 | tr | 12.5 |
| 979 | 974 | β-Pinene | - | 0.4 | tr | 0.5 | - | 0.5 | tr | - |
| 992 | 988 | Myrcene | - | 1.0 | tr | 0.6 | 1.5 | 1.2 | tr | 1.2 |
| 992 | 984 | 2-Pentyl furan | 2.6 | - | tr | - | - | - | - | - |
| 1004 | 998 | n-Octanal | 0.3 | - | 0.4 | 0.3 | - | - | - | - |
| 1007 | 1002 | α-Phellandrene | tr | tr | - | 0.8 | tr | tr | - | 1.7 |
| 1026 | 1020 | p-Cymene | 11.5 | 4.9 | 14.1 | 25.4 | 11.1 | 3.3 | 40.3 | 23.3 |
| 1030 | 1024 | Limonene | 4.7 | 6.2 | 3.4 | 14.4 | 11.6 | 6.9 | 21.6 | 23.9 |
| 1037 | 1032 | (Z)-β-Ocimene | tr | 1.3 | tr | tr | - | 3.0 | - | - |
| 1050 | 1044 | (E)-β-Ocimene | tr | 23.2 | 1.7 | 0.2 | tr | 40.9 | 2.5 | - |
| 1059 | 1054 | γ-Terpinene | 2.4 | tr | tr | 11.4 | 4.7 | - | tr | 15.4 |
| 1068 | 1065 | cis-Sabinene hydrate | - | - | tr | 0.2 | - | - | - | - |
| 1092 | 1086 | Terpinolene | 36.2 | 18.1 | 2.7 | 1.8 | 58.5 | 18.6 | 6.1 | 1.5 |
| 1092 | 1089 | p-Cymenene | tr | tr | 1.9 | tr | - | - | - | - |
| 1127 | 1122 | α-Campholenal | tr | tr | 1.5 | tr | - | - | - | - |
| 1140 | 1135 | trans-Pinocarveol | - | tr | 1.8 | tr | - | - | - | - |
| 1142 | 1137 | cis-Verbenol | - | - | 0.8 | - | - | - | - | - |
| 1142 | 1137 | (E)-Epoxy-ocimene | - | 0.6 | - | - | - | 0.2 | - | - |
| 1147 | - | 4,8-Epoxy-p-menth-1-ene | 3.0 | 5.1 | 8.1 | 0.5 | 2.5 | 1.9 | 3.3 | tr |
| 1163 | 1160 | Pinocarvone | - | - | 0.6 | tr | - | - | - | - |
| 1178 | - | 1,8-Menthadien-4-ol | tr | 1.2 | 6.6 | 0.9 | - | - | - | - |
| 1190 | 1179 | p-Cymen-8-ol | 6.2 | 13.0 | 21.8 | 1.3 | 5.3 | 1.5 | 3.4 | tr |
| 1199 | 1195 | Myrtenal | - | tr | 0.9 | tr | - | - | - | - |
| 1220 | 1215 | trans-Carveol | - | tr | 1.1 | tr | - | - | - | - |
| 1245 | 1239 | Carvone | tr | tr | 1.3 | - | - | - | - | - |
| 1286 | 1287 | Bornyl acetate | 0.9 | tr | 1.0 | tr | - | - | - | - |
| 1317 | 1315 | (2E,4E)-Decadienal | 1.1 | - | tr | - | - | - | - | - |
| 1357 | 1352 | 2,3,6-Trimethyl benzaldehyde | 0.6 | 0.8 | tr | tr | - | - | - | - |
| 1392 | 1389 | β-Elemene | 2.6 | tr | tr | 4.2 | tr | - | tr | 0.3 |
| 1420 | 1417 | (E)-Caryophyllene | 0.7 | 1.2 | 1.9 | 1.2 | tr | 0.1 | tr | tr |
| 1434 | 1434 | γ-Elemene | - | - | - | 0.3 | - | - | - | - |
| 1455 | 1452 | α-Humulene | tr | tr | tr | 2.1 | - | - | - | - |
| 1482 | 1484 | Germacrene D | - | - | - | 0.5 | - | - | - | - |
| 1487 | 1489 | β-Selinene | 2.2 | - | tr | 0.6 | - | - | - | - |
| 1497 | 1498 | α-Selinene | 2.8 | - | tr | - | - | - | - | - |
| 1498 | 1498 | α-Selinene + Bicyclogermacrene | - | - | - | 1.4 | - | - | - | - |
| 1511 | 1505 | β-Bisabolene | 8.7 | tr | tr | 1.8 | - | - | - | - |
| 1543 | 1545 | Selina-3,7(11)-diene | - | - | - | 0.8 | - | - | - | - |
| 1561 | 1559 | Germacrene B | - | - | - | 5.6 | - | - | - | - |
| 1566 | 1561 | (E)-Nerolidol | - | - | - | 3.0 | - | - | - | - |
| 1581 | 1577 | Spathulenol | - | - | - | 1.2 | - | - | - | - |
| 1586 | 1582 | Caryophyllene oxide | 1.6 | 5.7 | 13.1 | 2.0 | - | - | - | - |
| 1611 | 1608 | Humulene epoxide II | - | tr | 0.7 | 0.7 | - | - | - | - |
| 1638 | - | Caryophylla-dien-5-ol isomer | - | tr | 1.2 | tr | - | - | - | - |
| 1656 | 1658 | Selin-11-en-4-α-ol | 0.9 | - | tr | 0.2 | - | - | - | - |
| 1844 | - | Hexahydrofarnesyl acetone | - | 0.3 | 0.6 | tr | - | - | - | - |
| 1866 | - | Pentadecanoic acid | 0.8 | - | - | - | - | - | - | - |
| 1971 | 1959 | Hexadecanoic acid | 3.8 | - | - | - | - | - | - | - |
| 2137 | - | Octadecadienoic acid isomer | 0.6 | - | - | - | - | - | - | - |
| Monoterpene hydrocarbons | 56.2 | 67.2 | 30.5 | 66.5 | 92.3 | 93.0 | 93.3 | 99.4 | ||
| Oxygenated monoterpenes | 10.0 | 19.9 | 45.6 | 2.9 | 7.7 | 3.6 | 6.7 | tr | ||
| Sesquiterpene hydrocarbons | 17.0 | 1.2 | 1.9 | 18.4 | tr | 0.1 | tr | 0.3 | ||
| Oxygenated sesquiterpenes | 2.5 | 6.0 | 15.6 | 7.1 | - | - | - | - | ||
| Others | 9.9 | 0.8 | 0.4 | 0.3 | - | - | - | - | ||
| Total identified compounds | 95.6 | 95.1 | 94.0 | 95.2 | 100.0 | 96.7 | 100.0 | 99.7 | ||
| No of identified compounds | 32 | 32 | 39 | 43 | 15 | 16 | 15 | 14 | ||
| Rt 1 | Fatty Acid 2 | Roots Extract, % 3 | Fruits Extract, % 3 |
|---|---|---|---|
| 15.03 | Lauric acid (C12:0) | tr | - |
| 18.30 | Myristic acid (C14:0) | 0.2 | 0.1 |
| 20.02 | Pentadecanoic acid (C15:0) | 1.5 | 0.1 |
| 21.75 | Palmitic acid (C16:0) | 14.9 | 5.6 |
| 22.83 | Palmitoleic acid (C16:1n7c) | 0.6 | 0.3 |
| 23.41 | Heptadecanoic acid (C17:0) | 0.4 | 0.1 |
| 25.04 | Stearic acid (C18:0) | 1.0 | 1.0 |
| 25.98 | Petroselinic acid (C18:1n12c) | - | 49.9 |
| 26.03 | Oleic acid (C18:1n9c) | 14.2 | 10.2 |
| 26.12 | cis-Vaccenic acid (C18:1n7c) | 1.9 | 1.0 |
| 27.37 | Linoleic acid (C18:2n6c) | 51.8 | 28.3 |
| 28.08 | Arachidic acid (C20:0) | 0.6 | 0.3 |
| 28.84 | α-Linolenic acid (C18:3n3) | 3.6 | 0.8 |
| 29.52 | Heneicosanoic acid (C21:0) | tr | 0.1 |
| 30.92 | Behenic acid (C22:0) | 0.7 | 0.4 |
| 32.33 | Tricosanoic acid (C23:0) | 0.3 | 0.2 |
| 33.80 | Lignoceric acid (C24:0) | 0.9 | 0.3 |
| Saturated fatty acids | 20.4 | 8.2 | |
| Monounsaturated fatty acids | 16.8 | 61.3 | |
| Polyunsaturated fatty acids | 55.4 | 29.1 | |
| Total identified fatty acids | 92.6 | 98.7 |
| Rt 1 | Compound 2 | UFRE, % 3 | UFFE, % 3 |
|---|---|---|---|
| 78.24 | Campesterol | 2.7 | 1.3 |
| 79.26 | Stigmasterol | 18.5 | 26.3 |
| 80.51 | Δ5,23-Stigmastadienol | 0.9 | - |
| 80.64 | β-Amyrin | - | 4.1 |
| 81.23 | β-Sitosterol + α-spinasterol | - | 52.7 |
| 81.29 | β-Sitosterol | 60.0 | - |
| 81.48 | Stigmastanol | 3.3 | - |
| 83.10 | Δ7-Stigmastenol | 2.8 | 7.1 |
| 83.71 | Δ7-Avenasterol | 1.1 | 1.7 |
| Total identified | 89.4 | 93.3 | |
| Total sterols/triterpenes in unsaponifiable fractions | 93.7 | 69.6 |
| Rt, min | Coumarin (Registered Number) 1 | λmax, nm | m/z (100 V) | m/z (250 V) |
|---|---|---|---|---|
| 13.17 | Heraclenol (1) 2 | 218, 248, 302 | 631.1 (36.5) 4, 327.0 (100.0), 322.1 (17.1), 287.0 (39.2), 269.10 (13.0), 121.2 (25.1) | 327.1 (100.0), 240.9 (12.7), 203.0 (24.5), 173.0 (10.8), 147.10 (24.9), 129.0 (12.7) |
| 13.81 | 2′,3′-Dihydrofuranocoumarin derivative (2) | 222, 334 | 269.1 (33.5), 247.1 (100.0), 121.1 (16.6) | 269.0 (100.0), 247.1 (51.5), 229.1 (40.1), 213.1 (12.6), 175.0 (58.3), 147.0 (31.6) |
| 14.66 | Oxypeucedanin hydrate (3) | 222, 250, 266, 310 | 631.1 (100.0), 327.1 (12.2), 305.1 (94.8) | 327.0 (100.0), 203.0 (51.3), 147.0 (18.1) |
| 15.48 | Xanthotoxin (4) 3 | 218, 248, 302 | 455.0 (49.3), 239.0 (22.4), 217.1 (100.0) | 217.0 (13.5), 202.0 (72.1), 174.1 (100.0), 161.0 (15.6), 146.0 (13.5), 131.0 (12.4), 118.1 (14.8) |
| 18.01 | Heraclenin (5) 2 | 218, 248, 302 | 595.1 (84.1), 309.0 (81.8), 304.1 (15.2), 287.1 (100.0), 269.1 (70.5), 203.0 (10.6) | 309.1 (100.0), 224.0 (15.7), 203 (61.7), 173.0 (24.7), 157.0 (16.7), 147.1 (63.9), 129.1 (29.4), 89.1 (19.6) |
| 19.69 | Oxypeucedanin (6) | 222, 250, 266, 310 | 595.1 (64.5), 287.0 (100.0) | 309.0 (100.0), 203.0 (57.4), 147.1 (37.1) |
| 22.05 | Imperatorin (7) | 218, 248, 302 | 563.1 (100.0), 293.0 (33.9), 271.1 (87.9), 203.0 (43.0) | 293.0 (12.3), 224.0 (43.3), 203.1 (35.6), 175.0 (11.5), 147.0 (100.0), 129.0 (26.3), 91.1 (13.7) |
| 23.90 | 2′,3′-Dihydrofuranocoumarin derivative (8) 2 | 222, 334 | 679.2 (93.8), 351.1 (12.6), 329.1 (100.0) | 679.2 (11.8), 351.1 (100.0), 229.1 (46.2) |
| 24.29 | Isoimperatorin (9) | 222, 250, 266, 310 | 563.1 (100.0), 293.0 (11.8), 271.1 (93.3), 203.0 (33.7) | 293.0 (100.0), 224.0 (16.1), 203.0 (35.9), 147.1 (86.9), 91.1 (28.5) |
| 24.39 | Prantschimgin (10) | 222, 334 | 679.2 (75.4), 329.1 (100.0) | 679.2 (11.5), 351.1 (100.0), 247.1 (11.9), 229.1 (40.2) |
| Rt, min | Coumarin (Registered Number) 1 | Content in MFDEs and Expressed on the Crude (Whole) CH2Cl2 Extracts, mg/g | |||
|---|---|---|---|---|---|
| Root MFDE | Root Extract | Fruit MFDE | Fruit Extract | ||
| 13.17 | Heraclenol (1) 2 | - | - | 4.1 ± 1.8 | 2.8 ± 1.2 |
| 13.81 | 2′,3′-Dihydrofuranocoumarin derivative (2) 3 | 3.1 ± 0.2 | 2.8 ± 0.1 | 6.8 ± 0.6 | 4.7 ± 0.4 |
| 14.66 | Oxypeucedanin hydrate (3) 4 | 22.4 ± 0.1 | 20.3 ± 0.1 | 87.5 ± 5.9 | 60.8 ± 4.1 |
| 15.48 | Xanthotoxin (4) 2 | 11.0 ± 0.9 | 10.0 ± 0.8 | - | - |
| 18.01 | Heraclenin (5) 2 | - | - | 33.8 ± 10.3 | 23.5 ± 7.2 |
| 19.69 | Oxypeucedanin (6) | 18.4 ± 3.4 | 16.7 ± 3.1 | 346.0 ± 33.7 | 240.4 ± 23.4 |
| 22.05 | Imperatorin (7) | 36.6 ± 2.5 | 33.2 ± 2.3 | 32.5 ± 4.2 | 22.6 ± 2.9 |
| 23.90 | 2′,3′-Dihydrofuranocoumarin derivative (8) 3 | - | - | 54.6 ± 0.0 | 37.9 ± 0.0 |
| 24.29 | Isoimperatorin (9) 2 | 9.4 ± 0.9 | 8.5 ± 0.8 | 28.3 ± 8.3 | 19.7 ± 5.8 |
| 24.39 | Prantschimgin (10) | 118.7 ± 0.1 | 107.9 ± 0,1 | 96.3 ± 1.1 | 66.9 ± 0.7 |
| Essential Oil/Positive Control | Root | Leaf | Stem | Fruit | E211 | E224 | Standard Drugs 1 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC |
| Staphylococcus aureus (ATCC 11632) | 0.20 | 0.39 | 0.78 | 1.56 | 0.20 | 0.39 | 6.25 | 12.5 | 4.00 | 4.00 | 1.00 | 1.00 | 0.1 | 0.2 |
| Bacillus cereus (clinical isolate) | 0.20 | 0.39 | 0.78 | 1.56 | 0.20 | 0.39 | 0.78 | 1.56 | 0.50 | 0.50 | 2.00 | 4.00 | 0.025 | 0.05 |
| Listeria monocytogenes (NCTC 7973) | 3.13 | 6.25 | 3.13 | 6.25 | 1.56 | 3.13 | 6.25 | 12.5 | 1.00 | 2.00 | 0.50 | 1.00 | 0.15 | 0.3 |
| Salmonella Typhimurium (ATCC 13311) | 1.56 | 3.13 | 1.56 | 3.13 | 0.78 | 1.56 | 3.13 | 6.25 | 1.00 | 2.00 | 1.00 | 1.00 | 0.1 | 0.2 |
| Escherichia coli (ATCC 25922) | 6.25 | 12.5 | 1.56 | 3.13 | 1.56 | 3.13 | 6.25 | 12.5 | 1.00 | 2.00 | 0.50 | 1.00 | 0.1 | 0.2 |
| Enterobacter cloacae (ATCC 35030) | 1.56 | 3.13 | 1.56 | 3.13 | 1.56 | 3.13 | 6.25 | 12.5 | 2.00 | 4.00 | 0.50 | 0.50 | 0.025 | 0.05 |
| Fungi | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC |
| Aspergillus fumigatus (ATCC 9197) | 0.10 | 0.20 | 0.39 | 0.78 | 0.20 | 0.39 | 0.20 | 0.39 | 1.00 | 2.00 | 1.00 | 1.00 | 0.025 | 0.05 |
| Aspergillus versicolor (ATCC 11730) | 0.78 | 1.56 | 0.78 | 1.56 | 0.39 | 0.78 | 0.78 | 1.56 | 2.00 | 4.00 | 1.00 | 1.00 | 0.006 | 0.0125 |
| Aspergillus niger (ATCC 6275) | 0.20 | 0.39 | 0.39 | 0.78 | 0.20 | 0.39 | 0.39 | 0.78 | 1.00 | 2.00 | 1.00 | 1.00 | 0.10 | 0.20 |
| Penicillium funiculosum (ATCC 36839) | 0.10 | 0.20 | 0.10 | 0.20 | 0.10 | 0.20 | 0.10 | 0.20 | 1.00 | 2.00 | 0.50 | 0.50 | 0.0125 | 0.025 |
| Penicillium verrucosum (food isolate) | 0.10 | 0.20 | 0.10 | 0.20 | 0.10 | 0.20 | 0.10 | 0.20 | 2.00 | 4.00 | 1.00 | 1.00 | 0.06 | 0.0125 |
| Trichoderma harzianum (TH-IS005-12) | 0.10 | 0.20 | 0.10 | 0.20 | 0.20 | 0.39 | 0.39 | 0.39 | 1.00 | 2.00 | 0.50 | 0.50 | 0.2 | 0.4 |
| Candida albicans (ATCC 10231) | 1.56 | 3.13 | 1.56 | 3.13 | 3.13 | 6.25 | 1.56 | 3.13 | 1.00 | 2.00 | 1.00 | 2.00 | 0.0125 | 0.025 |
| Essential oils | Root | Leaf | Stem | Fruit | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC |
| Staphylococcus aureus | 39.4 ± 4.6 c | 37.7 ± 5.3 c | 24.2 ± 2.7 | 55.4 ± 3.8 d | 28.2 ± 2.9 b | NA1 | 22.6 ± 4.2 b | 35.5 ± 4.3 c | NA | 14.9 ± 1.1 a | 2.5 ± 0.6 a | NA |
| Listeria monocytogenes | 79.3 ± 6.6 b | 80.2 ± 7.2 c | 21 ± 2.8 b | 87.1 ± 4.1 c | 82.6 ± 6.4 c | 50.3 ± 1.1 c | 17.1 ± 1.1 a | 32.4 ± 2.4 b | 22.9 ± 1.5 b | 89.1 ± 1.7 c | 15.7 ± 4.1 a | 2.8 ± 0.4 a |
| Escherichia coli | 51.1 ± 3.3 c | 27.3 ± 2.7 b | 15.0 ± 1.0 a | 62.5 ± 5.4 d | 13.1 ± 1.5 a | NA | 2.3 ± 0.1 a | NA | NA | 45.2 ± 6.8 b | 46.3 ± 6.2 c | 27.2 ± 3.2 b |
| Fungus | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC |
| Candida albicans | 2.5 ± 0.3 b | NA | NA | 1.2 ± 0.1 a | NA | NA | NA | NA | NA | 3.3 ± 0.1 c | NA | NA |
| MFDE/Positive Control | Root | Fruit | Streptomycin | Ketoconazole | ||||
|---|---|---|---|---|---|---|---|---|
| Bacteria | MIC | MBC | MIC | MBC | MIC | MBC | MIC | MBC |
| Staphylococcus aureus (ATCC 11632) | 1 | 2 | 2 | 4 | 0.1 | 0.2 | - | - |
| Bacillus cereus (clinical isolate) | 0.125 | 0.25 | 0.125 | 0.25 | 0.025 | 0.05 | - | - |
| Listeria monocytogenes (NCTC 7973) | 1 | 2 | 1 | 2 | 0.15 | 0.3 | - | - |
| Salmonella Typhimurium (ATCC 13311) | 1 | 2 | 1 | 2 | 0.1 | 0.2 | - | - |
| Escherichia coli (ATCC 25922) | 4 | 8 | 4 | 8 | 0.1 | 0.2 | - | - |
| Enterobacter cloacae (ATCC 35030) | 4 | 8 | 2 | 4 | 0.025 | 0.05 | - | - |
| Fungi | MIC | MFC | MIC | MFC | MIC | MFC | MIC | MFC |
| Aspergillus fumigatus (ATCC 9197) | 0.25 | 0.5 | 0.25 | 0.5 | - | - | 0.025 | 0.05 |
| Aspergillus versicolor (ATCC 11730) | 0.5 | 1 | 0.5 | 1 | - | - | 0.006 | 0.0125 |
| Aspergillus niger (ATCC 6275) | 0.0625 | 0.125 | 0.0625 | 0.125 | - | - | 0.10 | 0.20 |
| Penicillium funiculosum (ATCC 36839) | 0.5 | 1 | 0.5 | 1 | - | - | 0.0125 | 0.025 |
| Penicillium verrucosum var. cyclopium (food isolate) | 1 | 2 | 1 | 2 | - | - | 0.06 | 0.0125 |
| Trichoderma harzianum (TH-IS005-12) | 1 | 2 | 1 | 2 | - | - | 0.2 | 0.4 |
| Candida albicans (ATCC 10231) | 1 | 2 | 1 | 2 | - | - | 0.0125 | 0.025 |
| MFDE | Root | Fruit | ||||
|---|---|---|---|---|---|---|
| Bacteria | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC |
| Staphylococcus aureus (ATCC 11632) | 2.7 ± 1.6 | NA 1 | NA | NA | 28.3 ± 3.7 | NA |
| Listeria monocytogenes (NCTC 7973) | 3.2 ± 3.4 | 15.2 ± 4.3 | 27.6 ± 2.2 b | NA | NA | 1.3 ± 0.9 a |
| Escherichia coli (ATCC 25922) | NA | NA | NA | 2.3 ± 1.8 | NA | NA |
| Fungus | MIC | ½MIC | ¼MIC | MIC | ½MIC | ¼MIC |
| Candida albicans (ATCC 10231) | 53.5 ± 6.0 a | 32.3 ± 5.2 b | 26.8 ± 10.2 b | 48.6 ± 5.1 a | 23.0 ± 3.9 a | 14.3 ± 7.9 a |
| MFDE | MIC | ½MIC | ¼MIC |
|---|---|---|---|
| Root | 23.7 ± 8.1 1,a | 14.4 ± 5.4 a | 33.7 ± 6.1 b |
| Fruit | 31.5 ± 5.1 b | 21.5 ± 7.0 b | 13.1 ± 3.7 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ušjak, L.; Stojković, D.; Carević, T.; Milutinović, V.; Soković, M.; Niketić, M.; Petrović, S. Chemical Analysis and Investigation of Antimicrobial and Antibiofilm Activities of Prangos trifida (Apiaceae). Antibiotics 2024, 13, 41. https://doi.org/10.3390/antibiotics13010041
Ušjak L, Stojković D, Carević T, Milutinović V, Soković M, Niketić M, Petrović S. Chemical Analysis and Investigation of Antimicrobial and Antibiofilm Activities of Prangos trifida (Apiaceae). Antibiotics. 2024; 13(1):41. https://doi.org/10.3390/antibiotics13010041
Chicago/Turabian StyleUšjak, Ljuboš, Dejan Stojković, Tamara Carević, Violeta Milutinović, Marina Soković, Marjan Niketić, and Silvana Petrović. 2024. "Chemical Analysis and Investigation of Antimicrobial and Antibiofilm Activities of Prangos trifida (Apiaceae)" Antibiotics 13, no. 1: 41. https://doi.org/10.3390/antibiotics13010041
APA StyleUšjak, L., Stojković, D., Carević, T., Milutinović, V., Soković, M., Niketić, M., & Petrović, S. (2024). Chemical Analysis and Investigation of Antimicrobial and Antibiofilm Activities of Prangos trifida (Apiaceae). Antibiotics, 13(1), 41. https://doi.org/10.3390/antibiotics13010041