
Unveiling the Relationship between Ceftobiprole and High-Molecular-Mass (HMM) Penicillin-Binding Proteins (PBPs) in Enterococcus faecalis

 and
and 
Abstract
:1. Introduction
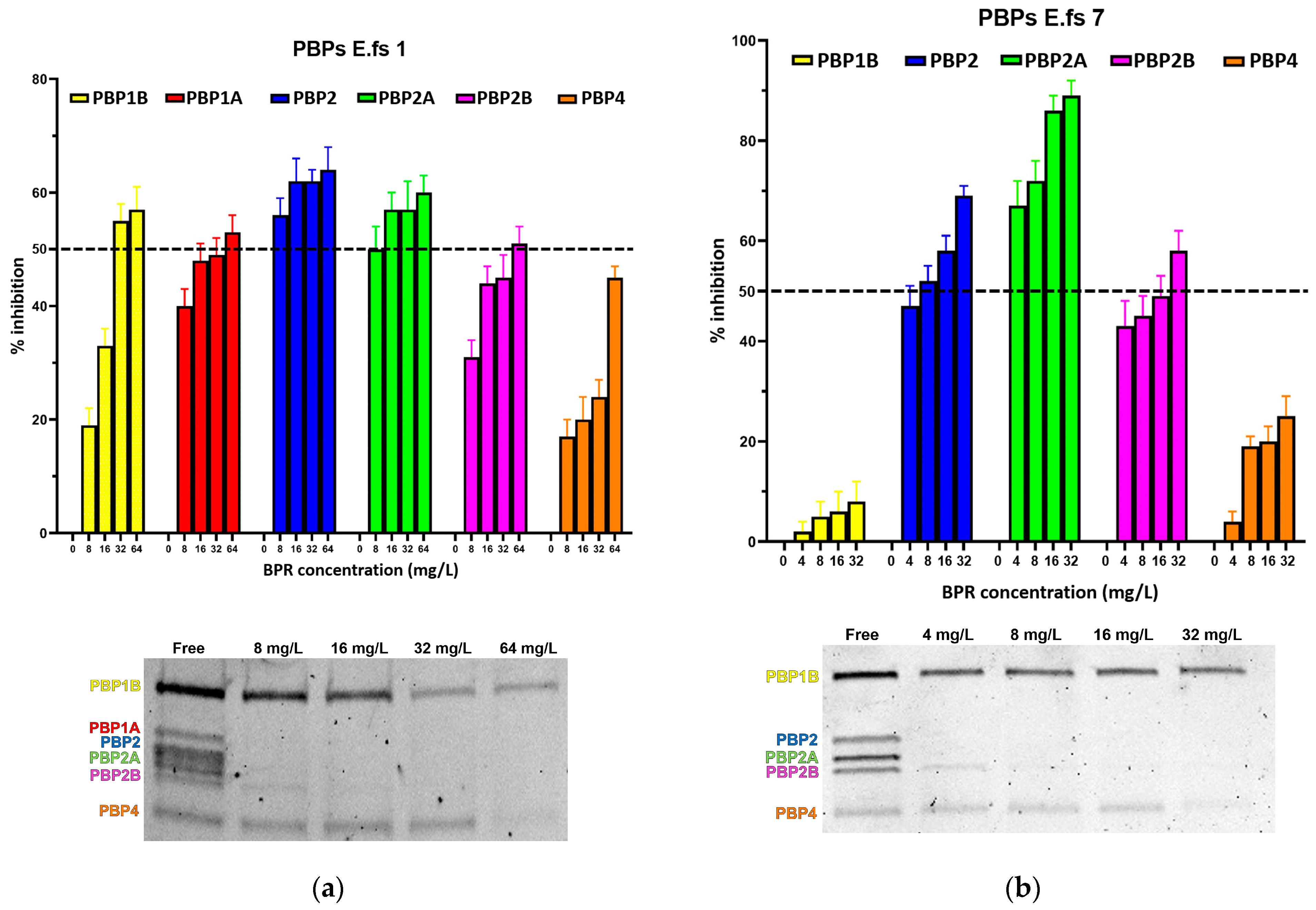
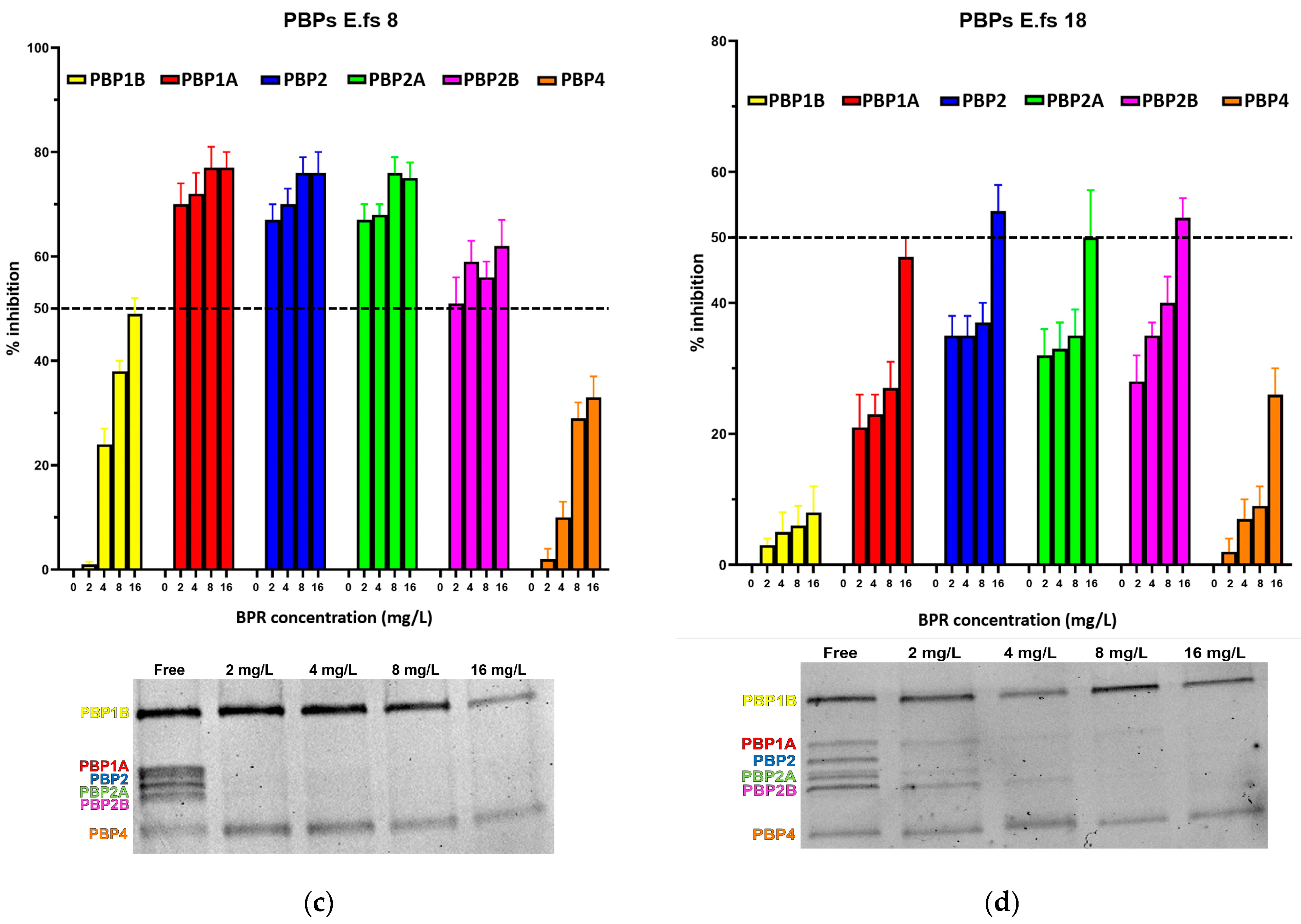
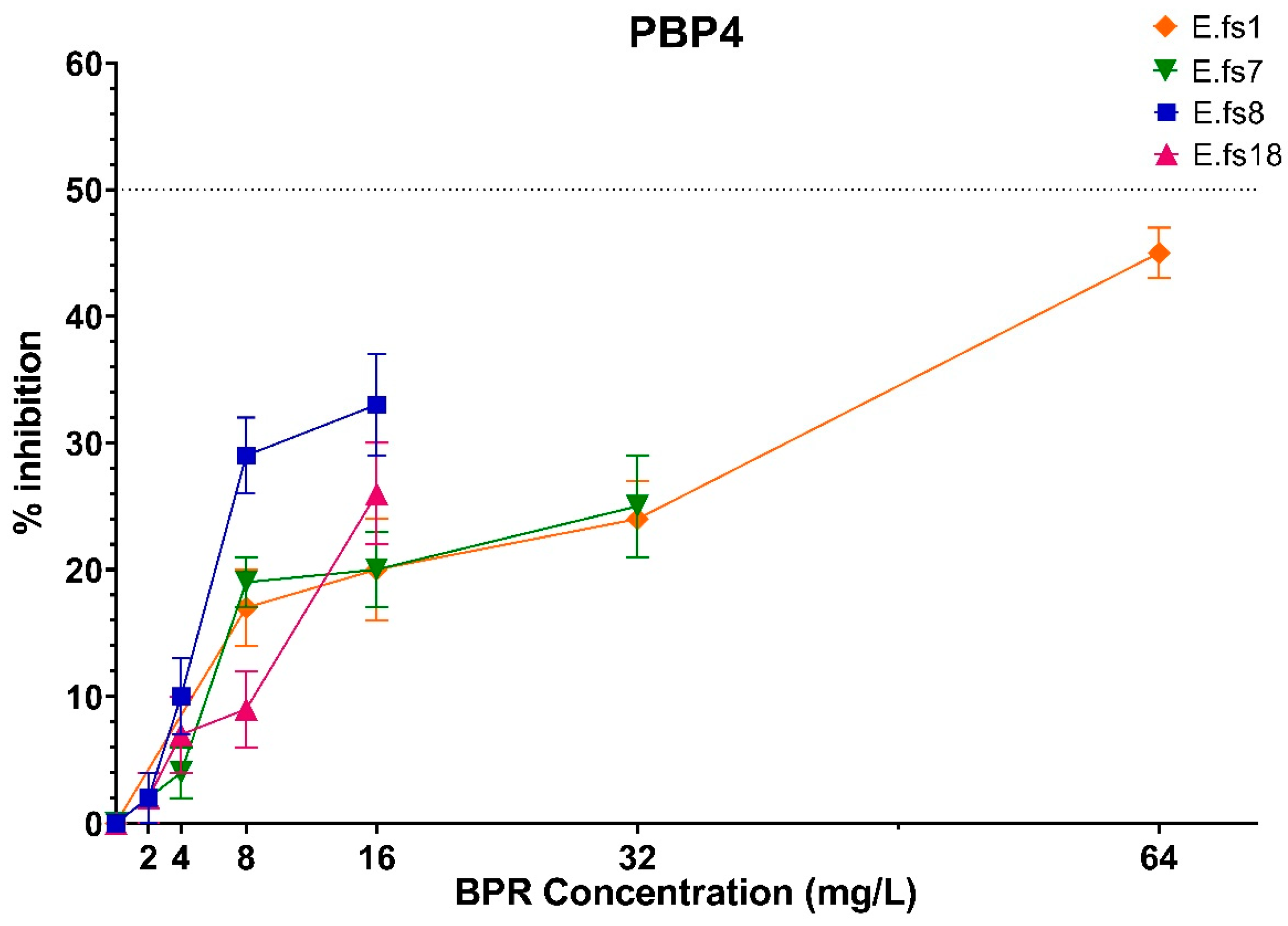
2. Results
2.1. Binding Profiles of HMM PBPs
2.2. Analysis of β-Lactamase Production
3. Discussion
4. Materials and Methods
4.1. Selected Strains
4.2. Study of PBP4/BPR Affinity and Competition Assay
4.3. β-Lactamase Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hollenbeck, B.L.; Rice, L.B. Intrinsic and acquired resistance mechanisms in Enterococcus. Virulence 2012, 3, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Weiner, L.M.; Webb, A.K.; Limbago, B.; Dudeck, M.A.; Patel, J.; Kallen, A.; Edwards, J.R.; Sievert, D.M. Antimicrobial-resistant pathogens associated with healthcare-associated infections: Summary of data reported to the National Healthcare Safety Network at the Centers for Disease Control and Prevention, 2011–2014. Infect. Control Hosp. Epidemiol. 2016, 37, 1288–1301. [Google Scholar] [CrossRef]
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti Infect Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef] [PubMed]
- Born, P.; Breukink, E.; Vollmer, W. In vitro synthesis of cross-linked murein and its attachment to sacculi by PBP1A from Escherichia coli. J. Biol. Chem. 2006, 281, 26985–26993. [Google Scholar] [CrossRef] [PubMed]
- Moon, T.M.; D’Andréa, É.D.; Lee, C.W.; Soares, A.; Jakoncic, J.; Desbonnet, C.; Garcia-Solache, M.; Rice, L.B.; Page, R.; Peti, W. The structures of penicillin-binding protein 4 (PBP4) and PBP5 from Enterococci provide structural insights into β-lactam resistance. J. Biol. Chem. 2018, 293, 18574–18584. [Google Scholar] [CrossRef] [PubMed]
- Zapun, A.; Vernet, T.; Pinho, M. The different shapes of cocci. FEMS Microbiol. Rev. 2008, 32, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Arbeloa, A.; Segal, H.; Hugonnet, J.E.; Josseaume, N.; Dubost, L.; Brouard, J.P.; Gutmann, L.; Mengin-Lecreulx, D.; Arthur, M. Role of class A penicillin-binding proteins in PBP5-mediated β-lactam resistance in Enterococcus faecalis. J. Bacteriol. 2004, 186, 1221–1228. [Google Scholar] [CrossRef]
- Duez, C.; Zorzi, W.; Sapunaric, F.; Amoroso, A.; Thamm, I.; Coyette, J. The penicillin resistance of Enterococcus faecalis JH2-2R results from an overproduction of the low-affinity penicillin-binding protein PBP4 and does not involve a psr-like gene. Microbiology 2001, 147, 2561–2569. [Google Scholar] [CrossRef]
- Zapun, A.; Contreras-Martel, C.; Vernet, T. Penicillin binding proteins and beta-lactam resistance. FEMS Microbiol. Rev. 2008, 32, 361–385. [Google Scholar] [CrossRef]
- Ghuysen, J.M. Serine β -lactamases and penicillin-binding proteins. Annu. Rev. Microbiol. 1991, 45, 37–67. [Google Scholar] [CrossRef]
- Lazzaro, L.M.; Cassisi, M.; Stefani, S.; Campanile, F. Impact of PBP4 Alterations on β-Lactam Resistance and Ceftobiprole Non-Susceptibility Among Enterococcus faecalis Clinical Isolates. Front Cell Infect. Microbiol. 2022, 11, 816657. [Google Scholar] [CrossRef]
- Rice, L.B.; Desbonnet, C.; Tait-Kamradt, A.; Garcia-Solache, M.; Lonks, J.; Moon, T.M.; D’Andréa, É.D.; Page, R.; Peti, W. Structural and regulatory changes in PBP4 trigger decreased β-lactam susceptibility in Enterococcus faecalis. mBio 2018, 9, e00361-18. [Google Scholar] [CrossRef]
- Sauvage, E.; Kerff, F.; Terrak, M.; Ayala, J.A.; Charlier, P. The penicillin-binding proteins: Structure and role in peptidoglycan biosynthesis. FEMS Microbiol. Rev. 2008, 32, 234–258. [Google Scholar] [CrossRef]
- Djorić, D.; Little, J.L.; Kristich, C.J. Multiple Low-Reactivity Class B Penicillin-Binding Proteins Are Required for Cephalosporin Resistance in Enterococci. Antimicrob. Agents Chemother. 2020, 64, e02273-19. [Google Scholar] [CrossRef]
- Conceição, N.; da Silva, L.E.; Darini, A.L.; Pitondo-Silva, A.; de Oliveira, A.G. Penicillin Resistant, Ampicillin-Susceptible Enterococcus faecalis of Hospital Origin: Pbp4 Gene Polymorphism and Genetic Diversity. Infect. Genet. Evol. 2014, 28, 289–295. [Google Scholar] [CrossRef]
- Gawryszewska, I.; Zabicka, D.; Hryniewicz, W.; Sadowy, E. Penicillin-Resistant, Ampicillin-Susceptible Enterococcus faecalis in Polish Hospitals. Microb. Drug Resist. 2021, 27, 291–300. [Google Scholar] [CrossRef]
- Infante, V.H.P.; Conceic, N.; Goncalves de Oliveira, A.; Costa Darini, A.L. Evaluation of polymorphisms in pbp4 gene and genetic diversity in penicillin-resistant, ampicillin-susceptible Enterococcus faecalis from hospitals in different states in Brazil. FEMS Microbiol. Lett. 2016, 363, fnw044. [Google Scholar] [CrossRef]
- Werth, B.J.; Abbott, A.N. The combination of ampicillin plus ceftaroline is synergistic against Enterococcus faecalis. J. Antimicrob. Chemother. 2015, 70, 2414–2417. [Google Scholar] [CrossRef]
- Arias, C.A.; Singh, K.V.; Panesso, D.; Murray, B.E. Time-kill and synergism studies of ceftobiprole against Enterococcus faecalis, including beta-lactamase-producing and vancomycin-resistant isolates. Antimicrob. Agents Chemother. 2007, 51, 2043–2047. [Google Scholar] [CrossRef] [PubMed]
- Sarti, M.; Campanile, F.; Sabia, C.; Santagati, M.; Gargiulo, R.; Stefani, S. Polyclonal diffusion of beta-lactamase-producing Enterococcus faecium. J. Clin. Microbiol. 2012, 50, 169–172. [Google Scholar] [CrossRef]
- The European Committee on Antimicrobial Susceptibility Testing. Breakpoint Tables for Interpretation of MICs and Zone Diameters. EUCAST. 2021, Version 11.0. Available online: https://www.eucast.org/clinical_breakpoints (accessed on 1 January 2021).
- Bourgogne, A.; Garsin, D.A.; Qin, X.; Singh, K.V.; Sillanpaa, J.; Yerrapragada, S.; Ding, Y.; Dugan-Rocha, S.; Buhay, C.; Shen, H.; et al. Large scale variation in Enterococcus faecalis illustrated by the genome analysis of strain OG1RF. Genome Biol. 2008, 9, 1–16. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| PBP | PBP (Other Name) | PBP Class | a Locus_Tag | N. Amino Acids (aa) | b Molecular Weight (kDa) | c Enzymatic Activity | Gene | GenBank CP025020.1 (Position) | N. Nucleotides (bp) |
|---|---|---|---|---|---|---|---|---|---|
| PBP1B | PBPZ | class A | CVT43_07655 | 803 | 88.40413 | TPase/Gtase | pbp1b | 1509774-1512185 | 2412 |
| PBP1A | PONA | class A | CVT43_04970 | 778 | 85.35237 | TPase/Gtase | pbp1a | 431566-433752 | 2331 |
| PBP2 | PBPB | class B | CVT43_03860 | 742 | 81.71274 | TPase—Cell division protein FtsI | pbp2 | 747766-749994 | 2229 |
| PBP2A | PBPF | class A | CVT43_02290 | 728 | 79.56664 | TPase/Gtase | pbp2a | 966441-968777 | 2187 |
| PBP2B | PBPA | class B | CVT43_11360 | 711 | 77.85899 | TPase | pbp2b | 2276447-2278582 | 2136 |
| PBP4 | PBP4(5) | class B | CVT43_10030 | 680 | 74.01754 | TPase | pbp4 | 2010904-2012946 | 2043 |
| PBP | PBP | class C | CVT43_05000 | 498 | 57.71227 | D,D Carboxypeptidase M32 | pbp M32 | 972441-973937 | 1497 |
| VanY | PBP | class C | CVT43_11375 | 236 | 27.04 | D-Ala-D-Ala carboxypeptidase VanY | pbp vanY | 2281379-2282089 | 711 |
| Strain | IC50 (μg/mL) PBP | MIC BPR (mg/L) | |||||
|---|---|---|---|---|---|---|---|
| 1B | 1A | 2 | 2A | 2B | 4 | ||
| E.fs1 | 28.59 | 38.45 | 7.30 | 8.42 | 57.64 | N.A. | 16 |
| E.fs7 | N.A. | N.A. | 7 | 3.07 | 16.3 | N.A. | 8 |
| E.fs8 | N.A. | 1.53 | 1.53 | 0.31 | 2.2 | N.A. | 4 |
| E.fs18 | N.A. | N.A. | 14.2 | 15.19 | 14.2 | N.A. | 4 |
| Code | MDR-Phenotype | MIC Values (mg/L) | Deletion in Promoter Region b | Amino Acid Substitutions in PBP4 d | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| PBP Active Sites | ||||||||||||||||||||||
| P 1 | AMP | AML | IMI | BPR 2 | CPT 2 | VA | TEC | 50T | 223I | 418T | 475L | 536A | 573D | 605Y | 606V | 639L | 665T | 666D | 678T | |||
| E.fs1 | PRAS-BPR-NS-HLAR | 16 | 1 | 0.5 | 4 | 16 | >256 | 0.5 | 2 | 2013028_2013029 delA c | I | - | A | - | T | - | - | - | - | - | P | - |
| E.fs7 | PRAS-BPR-NS-HLAR | 64 | 4 | 4 | 4 | 8 | >256 | 1 | 2 | 2013028_2013029 delA c | I | - | - | - | - | - | - | A | F | I | - | A |
| E.fs8 | PRAS-BPR-NS-VRE a | 16 | 4 | 1 | 2 | 4 | 32 | >256 | >256 | 2013028_2013029 delA c | I | - | - | Q | - | - | - | - | - | - | - | - |
| E.fs18 | PRAS-BPR-NS | 16 | 2 | 1 | 2 | 4 | 4 | 1 | 2 | 2013028_2013029 delA c | - | - | - | - | - | - | H | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Conti, P.; Lazzaro, L.M.; Longo, F.; Lenzo, F.; Giardina, A.; Fortuna, S.A.; Stefani, S.; Campanile, F. Unveiling the Relationship between Ceftobiprole and High-Molecular-Mass (HMM) Penicillin-Binding Proteins (PBPs) in Enterococcus faecalis. Antibiotics 2024, 13, 65. https://doi.org/10.3390/antibiotics13010065
Conti P, Lazzaro LM, Longo F, Lenzo F, Giardina A, Fortuna SA, Stefani S, Campanile F. Unveiling the Relationship between Ceftobiprole and High-Molecular-Mass (HMM) Penicillin-Binding Proteins (PBPs) in Enterococcus faecalis. Antibiotics. 2024; 13(1):65. https://doi.org/10.3390/antibiotics13010065
Chicago/Turabian StyleConti, Paola, Lorenzo Mattia Lazzaro, Fabio Longo, Federica Lenzo, Alessandra Giardina, Sebastiano Alberto Fortuna, Stefania Stefani, and Floriana Campanile. 2024. "Unveiling the Relationship between Ceftobiprole and High-Molecular-Mass (HMM) Penicillin-Binding Proteins (PBPs) in Enterococcus faecalis" Antibiotics 13, no. 1: 65. https://doi.org/10.3390/antibiotics13010065
APA StyleConti, P., Lazzaro, L. M., Longo, F., Lenzo, F., Giardina, A., Fortuna, S. A., Stefani, S., & Campanile, F. (2024). Unveiling the Relationship between Ceftobiprole and High-Molecular-Mass (HMM) Penicillin-Binding Proteins (PBPs) in Enterococcus faecalis. Antibiotics, 13(1), 65. https://doi.org/10.3390/antibiotics13010065