
Diverse Role of blaCTX-M and Porins in Mediating Ertapenem Resistance among Carbapenem-Resistant Enterobacterales

 ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. ErMs Predominantly Harbor blaCTX-M among NCPE
2.2. ErMs E. coli Associates with Mobile Genetic Elements Interposed by blaCTX-M
2.3. Carbapenemase and blaCTX-M Hastens Meropenem Hydrolysis in CPE and NCPE
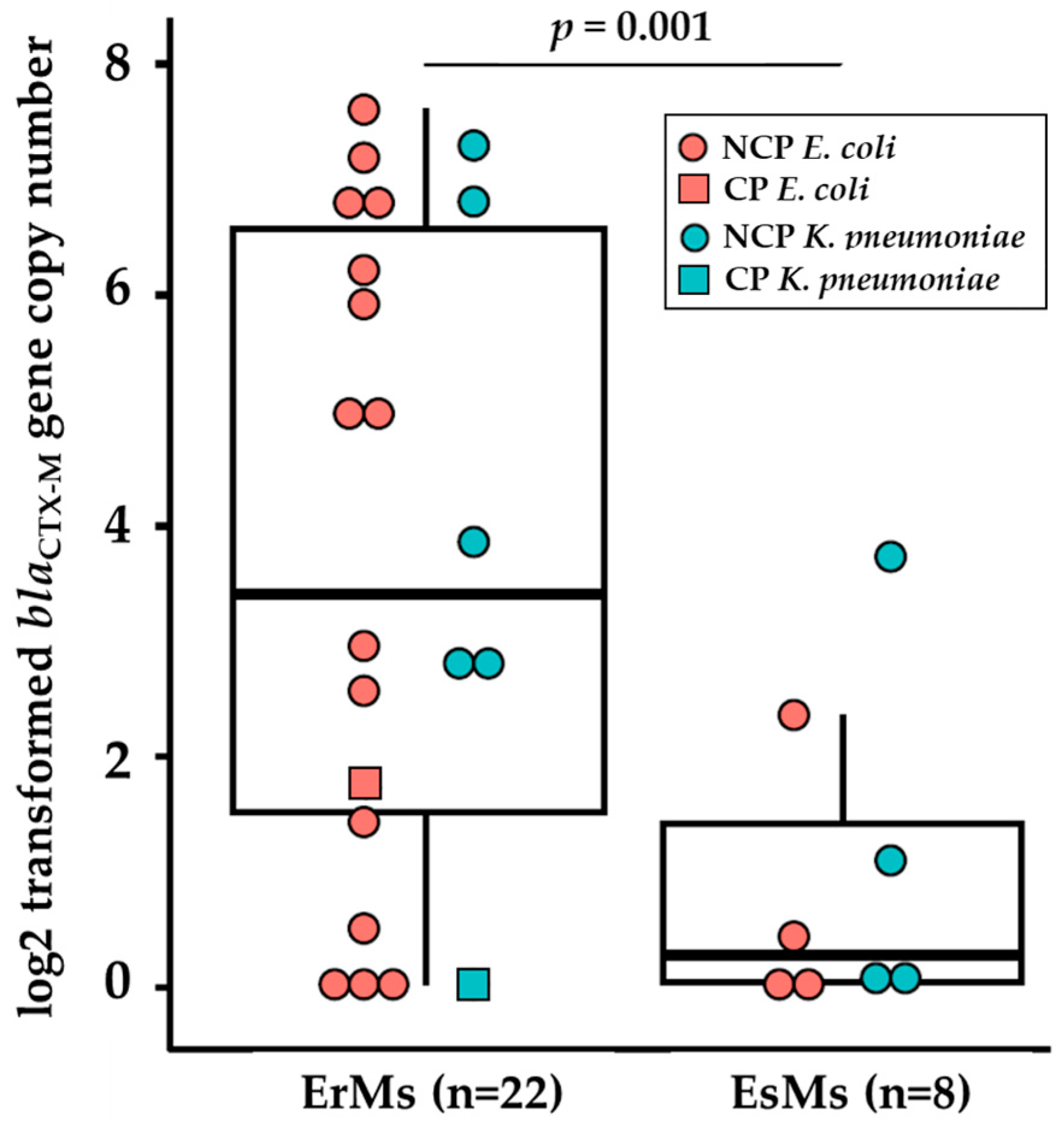
2.4. Ertapenem-Resistant E. coli and K. pneumoniae Carry Elevated Copies of blaCTX-M Genes
2.5. Porin Alterations Are Frequent among Ertapenem-Resistant NCPE E. coli

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Carbapenemase Status | blaCTX-M Δ∆Ct A | Contig Coverage B at K12 ompC | OmpC Band C |
|---|---|---|---|---|
| EC12 | CP (blaKPC) | +1.7 | No gap (Full coverage) | Detected |
| EC14 | CP (blaKPC) | +0.5 | No gap (Low at c.539–c.545) | Detected |
| EC13 | CP (blaKPC) | +5.0 | No gap (Full coverage) | ND |
| EC75 | CP (blaKPC) | ND | No gap (Full coverage) | Detected |
| EC30 | NCP | +6.2 | 149 bp gap (c.424—c.531) | ND |
| EC31 | NCP | +2.9 | 29 bp gap (c.544–c.531) | ND |
| EC35 | NCP | +5.9 | 144 bp gap (c.429–c.531) | ND |
| EC2 | NCP | +4.9 | 149 bp gap (c.424–c.531) | ND |
| EC3 | NCP | +6.6 | 173 bp gap (c.416–c.515) | ND |
| EC32 | NCP | +1.4 | 150 bp gap (c.424–c.530) | ND |
| EC33 | NCP | +7.5 | 149 bp gap (c.424–c.530) | ND |
| EC34 | NCP | ND | 139 bp gap D (c.434–c.531) | ND |
| EC36 | NCP | +7.1 | 140 bp gap (c.434–c.530) | ND |
| EC4 | NCP | +6.8 | No gap (Full coverage) | ND |
| EC6 | NCP | +2.5 | 141 bp gap (c.431–c.532) | ND |
| EC66 | NCP | ND | 149 bp gap (c.424–c.531) | ND |

2.6. Ertapenem-Resistant E. coli Lack OmpC Outer Membrane Protein
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates and Antimicrobial Susceptibility Testing
4.2. Whole Genome Sequencing
4.3. Immunodetection and Sample Preparation
4.4. qPCR of β-lactamase Genes
4.5. Sample Preparation for LC-MS/MS Analysis
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Implementation Manual to Prevent and Control the Spread of Carbapenem-Resistant Organisms at the National and Health Care Facility Level: Interim Practical Manual Supporting Implementation of the Guidelines for the Prevention and Control of Carbapenem-Resistant Enterobacteriaceae, Acinetobacter Baumannii and Pseudomonas Aeruginosa in Health Care Facilities; World Health Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Lasko, M.J.; Nicolau, D.P. Carbapenem-Resistant Enterobacterales: Considerations for Treatment in the Era of New Antimicrobials and Evolving Enzymology. Curr. Infect. Dis. Rep. 2020, 22, 6. [Google Scholar] [CrossRef]
- Tracking CRE in the United States|HAI|CDC. Available online: https://www.cdc.gov/hai/organisms/cre/trackingcre.html (accessed on 31 January 2023).
- Karlsson, M.; Lutgring, J.D.; Ansari, U.; Lawsin, A.; Albrecht, V.; McAllister, G.; Daniels, J.; Lonsway, D.; McKay, S.; Beldavs, Z.; et al. Molecular Characterization of Carbapenem-Resistant Enterobacterales Collected in the United States. Microb. Drug Resist. 2022, 28, 389–397. [Google Scholar] [CrossRef]
- Lee, Y.Q.; Sri La Sri Ponnampalavanar, S.; Chong, C.W.; Karunakaran, R.; Vellasamy, K.M.; Abdul Jabar, K.; Kong, Z.X.; Lau, M.Y.; Teh, C.S.J. Characterisation of Non-Carbapenemase-Producing Carbapenem-Resistant Klebsiella Pneumoniae Based on Their Clinical and Molecular Profile in Malaysia. Antibiotics 2022, 11, 1670. [Google Scholar] [CrossRef]
- Chea, N.; Bulens, S.N.; Kongphet-Tran, T.; Lynfield, R.; Shaw, K.M.; Vagnone, P.S.; Kainer, M.A.; Muleta, D.B.; Wilson, L.; Vaeth, E.; et al. Improved Phenotype-Based Definition for Identifying Carbapenemase Producers among Carbapenem-Resistant Enterobacteriaceae. Emerg. Infect. Dis. 2015, 21, 1611–1616. [Google Scholar] [CrossRef]
- Black, C.A.; So, W.; Dallas, S.S.; Gawrys, G.; Benavides, R.; Aguilar, S.; Chen, C.-J.; Shurko, J.F.; Lee, G.C. Predominance of Non-Carbapenemase Producing Carbapenem-Resistant Enterobacterales in South Texas. Front. Microbiol. 2021, 11, 623574. [Google Scholar] [CrossRef] [PubMed]
- Tamma, P.D.; Aitken, S.L.; Bonomo, R.A.; Mathers, A.J.; van Duin, D.; Clancy, C.J. Infectious Diseases Society of America Antimicrobial-Resistant Treatment Guidance: Gram-Negative Bacterial Infections. Infectious Diseases Society of America 2023; Version 3.0. Available online: https://www.idsociety.org/practice-guideline/amr-guidance/ (accessed on 9 December 2023).
- Ma, P.; He, L.L.; Pironti, A.; Laibinis, H.H.; Ernst, C.M.; Manson, A.L.; Bhattacharyya, R.P.; Earl, A.M.; Livny, J.; Hung, D.T. Genetic Determinants Facilitating the Evolution of Resistance to Carbapenem Antibiotics. eLife 2021, 10, e67310. [Google Scholar] [CrossRef] [PubMed]
- Shropshire, W.C.; Aitken, S.L.; Pifer, R.; Kim, J.; Bhatti, M.M.; Li, X.; Kalia, A.; Galloway-Peña, J.; Sahasrabhojane, P.; Arias, C.A.; et al. IS26-Mediated Amplification of blaOXA-1 and blaCTX-M-15 with Concurrent Outer Membrane Porin Disruption Associated with de Novo Carbapenem Resistance in a Recurrent Bacteraemia Cohort. J. Antimicrob. Chemother. 2021, 76, 385–395. [Google Scholar] [CrossRef]
- Zankari, E.; Allesøe, R.; Joensen, K.G.; Cavaco, L.M.; Lund, O.; Aarestrup, F.M. PointFinder: A Novel Web Tool for WGS-Based Detection of Antimicrobial Resistance Associated with Chromosomal Point Mutations in Bacterial Pathogens. J. Antimicrob. Chemother. 2017, 72, 2764–2768. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef] [PubMed]
- Clausen, P.T.L.C.; Aarestrup, F.M.; Lund, O. Rapid and Precise Alignment of Raw Reads against Redundant Databases with KMA. BMC Bioinform. 2018, 19, 307. [Google Scholar] [CrossRef] [PubMed]
- Martínez-García, E.; Navarro-Lloréns, J.M.; Tormo, A. Identification of an Unknown Promoter, OUTIIp, within the IS10R Element. J. Bacteriol. 2003, 185, 2046–2050. [Google Scholar] [CrossRef]
- Pinet, E.; Franceschi, C.; Davin-Regli, A.; Zambardi, G.; Pagès, J.-M. Role of the Culture Medium in Porin Expression and Piperacillin-Tazobactam Susceptibility in Escherichia Coli. J. Med. Microbiol. 2015, 64, 1305–1314. [Google Scholar] [CrossRef]
- van Boxtel, R.; Wattel, A.A.; Arenas, J.; Goessens, W.H.F.; Tommassen, J. Acquisition of Carbapenem Resistance by Plasmid-Encoded-AmpC-Expressing Escherichia Coli. Antimicrob. Agents Chemother. 2017, 61, 10–1128. [Google Scholar] [CrossRef]
- Delgado, A.; Gawrys, G.W.; Duhon, B.M.; Koeller, J.M.; Lee, G.C. Impact of an Antimicrobial Stewardship Initiative on Ertapenem Use and Carbapenem Susceptibilities at Four Community Hospitals. J. Infect. Dis. Ther. 2017, 5, 341. [Google Scholar] [CrossRef]
- Shropshire, W.C.; Dinh, A.Q.; Earley, M.; Komarow, L.; Panesso, D.; Rydell, K.; Gómez-Villegas, S.I.; Miao, H.; Hill, C.; Chen, L.; et al. Accessory Genomes Drive Independent Spread of Carbapenem-Resistant Klebsiella Pneumoniae Clonal Groups 258 and 307 in Houston, TX. mBio 2022, 13, e00497-22. [Google Scholar] [CrossRef]
- Livermore, D.M.; Day, M.; Cleary, P.; Hopkins, K.L.; Toleman, M.A.; Wareham, D.W.; Wiuff, C.; Doumith, M.; Woodford, N. OXA-1 β-Lactamase and Non-Susceptibility to Penicillin/β-Lactamase Inhibitor Combinations among ESBL-Producing Escherichia Coli. J. Antimicrob. Chemother. 2019, 74, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Ahuja, S.K.; Manoharan, M.S.; Lee, G.C.; McKinnon, L.R.; Meunier, J.A.; Steri, M.; Harper, N.; Fiorillo, E.; Smith, A.M.; Restrepo, M.I.; et al. Immune Resilience despite Inflammatory Stress Promotes Longevity and Favorable Health Outcomes Including Resistance to Infection. Nat. Commun. 2023, 14, 3286. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.C.; Restrepo, M.I.; Harper, N.; Manoharan, M.S.; Smith, A.M.; Meunier, J.A.; Sanchez-Reilly, S.; Ehsan, A.; Branum, A.P.; Winter, C.; et al. Immunologic Resilience and COVID-19 Survival Advantage. J. Allergy Clin. Immunol. 2021, 148, 1176–1191. [Google Scholar] [CrossRef] [PubMed]
- Teshome, B.F.; Lee, G.C.; Reveles, K.R.; Attridge, R.T.; Koeller, J.; Wang, C.; Mortensen, E.M.; Frei, C.R. Application of a Methicillin-Resistant Staphylococcus Aureus Risk Score for Community-Onset Pneumonia Patients and Outcomes with Initial Treatment. BMC Infect. Dis. 2015, 15, 380. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.C.; Lawson, K.A.; Burgess, D.S. Clinical Epidemiology of Carbapenem-Resistant Enterobacteriaceae in Community Hospitals: A Case-Case-Control Study. Ann. Pharmacother. 2013, 47, 1115–1121. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.C.; Hall, R.G.; Boyd, N.K.; Dallas, S.D.; Du, L.C.; Treviño, L.B.; Retzloff, C.; Treviño, S.B.; Lawson, K.A.; Wilson, J.P.; et al. Predictors of Community-Associated Staphylococcus Aureus, Methicillin-Resistant and Methicillin-Susceptible Staphylococcus Aureus Skin and Soft Tissue Infections in Primary-Care Settings. Epidemiol. Infect. 2016, 144, 3198–3204. [Google Scholar] [CrossRef]
- Gawrys, G.W.; Tun, K.; Jackson, C.B.; Astorga, B.; Fetchick, R.J.; Septimus, E.; Lee, G.C. The Impact of Rapid Diagnostic Testing, Surveillance Software, and Clinical Pharmacist Staffing at a Large Community Hospital in the Management of Gram-Negative Bloodstream Infections. Diagn. Microbiol. Infect. Dis. 2020, 98, 115084. [Google Scholar] [CrossRef]
- Bandy, S.M.; Jackson, C.B.; Black, C.A.; Godinez, W.; Gawrys, G.W.; Lee, G.C. Molecular Rapid Diagnostics Improve Time to Effective Therapy and Survival in Patients with Vancomycin-Resistant Enterococcus Bloodstream Infections. Antibiotics 2023, 12, 210. [Google Scholar] [CrossRef]
- Lee, G.C.; Long, S.W.; Musser, J.M.; Beres, S.B.; Olsen, R.J.; Dallas, S.D.; Nunez, Y.O.; Frei, C.R. Comparative Whole Genome Sequencing of Community-Associated Methicillin-Resistant Staphylococcus Aureus Sequence Type 8 from Primary Care Clinics in a Texas Community. Pharmacother. J. Hum. Pharmacol. Drug Ther. 2015, 35, 220–228. [Google Scholar] [CrossRef]
- Lee, G.C.; Dallas, S.D.; Wang, Y.; Olsen, R.J.; Lawson, K.A.; Wilson, J.; Frei, C.R. Emerging Multidrug Resistance in Community-Associated Staphylococcus Aureus Involved in Skin and Soft Tissue Infections and Nasal Colonization. J. Antimicrob. Chemother. 2017, 72, 2461–2468. [Google Scholar] [CrossRef] [PubMed]
- Hasman, H.; Saputra, D.; Sicheritz-Ponten, T.; Lund, O.; Svendsen, C.A.; Frimodt-Møller, N.; Aarestrup, F.M. Rapid Whole-Genome Sequencing for Detection and Characterization of Microorganisms Directly from Clinical Samples. J. Clin. Microbiol. 2014, 52, 139–146. [Google Scholar] [CrossRef]
- Larsen, M.V.; Cosentino, S.; Lukjancenko, O.; Saputra, D.; Rasmussen, S.; Hasman, H.; Sicheritz-Pontén, T.; Aarestrup, F.M.; Ussery, D.W.; Lund, O. Benchmarking of Methods for Genomic Taxonomy. J. Clin. Microbiol. 2014, 52, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Larsen, M.V.; Cosentino, S.; Rasmussen, S.; Friis, C.; Hasman, H.; Marvig, R.L.; Jelsbak, L.; Sicheritz-Pontén, T.; Ussery, D.W.; Aarestrup, F.M.; et al. Multilocus Sequence Typing of Total-Genome-Sequenced Bacteria. J. Clin. Microbiol. 2012, 50, 1355–1361. [Google Scholar] [CrossRef]
- Bartual, S.G.; Seifert, H.; Hippler, C.; Luzon, M.A.D.; Wisplinghoff, H.; Rodríguez-Valera, F. Development of a Multilocus Sequence Typing Scheme for Characterization of Clinical Isolates of Acinetobacter Baumannii. J. Clin. Microbiol. 2005, 43, 4382–4390. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, D.; Fawley, W.; Kachrimanidou, M.; Bowden, R.; Crook, D.W.; Fung, R.; Golubchik, T.; Harding, R.M.; Jeffery, K.J.M.; Jolley, K.A.; et al. Multilocus Sequence Typing of Clostridium Difficile. J. Clin. Microbiol. 2010, 48, 770–778. [Google Scholar] [CrossRef]
- Lemee, L.; Dhalluin, A.; Pestel-Caron, M.; Lemeland, J.-F.; Pons, J.-L. Multilocus Sequence Typing Analysis of Human and Animal Clostridium Difficile Isolates of Various Toxigenic Types. J. Clin. Microbiol. 2004, 42, 2609–2617. [Google Scholar] [CrossRef]
- Wirth, T.; Falush, D.; Lan, R.; Colles, F.; Mensa, P.; Wieler, L.H.; Karch, H.; Reeves, P.R.; Maiden, M.C.J.; Ochman, H.; et al. Sex and Virulence in Escherichia Coli: An Evolutionary Perspective. Mol. Microbiol. 2006, 60, 1136–1151. [Google Scholar] [CrossRef]
- Jaureguy, F.; Landraud, L.; Passet, V.; Diancourt, L.; Frapy, E.; Guigon, G.; Carbonnelle, E.; Lortholary, O.; Clermont, O.; Denamur, E.; et al. Phylogenetic and Genomic Diversity of Human Bacteremic Escherichia Coli Strains. BMC Genom. 2008, 9, 560. [Google Scholar] [CrossRef]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual Molecular Dynamics. J. Mol. Graph. 1996, 14, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Adler, M.; Anjum, M.; Andersson, D.I.; Sandegren, L. Influence of Acquired β-Lactamases on the Evolution of Spontaneous Carbapenem Resistance in Escherichia Coli. J. Antimicrob. Chemother. 2013, 68, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Rivoarilala, O.L.; Garin, B.; Andriamahery, F.; Collard, J.M. Rapid in Vitro Detection of CTX-M Groups 1, 2, 8, 9 Resistance Genes by LAMP Assays. PLoS ONE 2018, 13, e0200421. [Google Scholar] [CrossRef] [PubMed]



| Name | Overall (n 99) (%Susceptible) | ErMs (n 29) (%Susceptible) |
|---|---|---|
| Amikacin | 91 | 65 |
| Aztreonam | 9 | 12 |
| Ceftazidime–avibactam | 77 | 88 |
| Ciprofloxacin | 9 | 12 |
| Colistin | 95 | 96 |
| Doripenem | 53 | 88 |
| Doxycycline | 44 | 38 |
| Ertapenem | 4 | 0 |
| Cefepime | 16 | 23 |
| Cefotaxime | 9 | 12 |
| Gentamicin | 39 | 48 |
| Imipenem | 46 | 73 |
| Imipenem–relebactam | 98 | 96 |
| Levofloxacin | 16 | 0 |
| Meropenem | 44 | 100 |
| Meropenem–vaborbactam | 88 | 88 |
| Minocycline | 63 | 68 |
| Polymyxin B | 95 | 96 |
| Piperacillin–tazobactam | 19 | 35 |
| Trimethoprim-sulfamethoxazole | 23 | 15 |
| Ceftazidime | 14 | 19 |
| Tigecycline | 98 | 96 |
| Ticarcillin–clavulanic acid | 11 | 15 |
| Tobramycin | 30 | 23 |
| Species | Carbapenem Phenotype | Carbapenemase Status | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| n (%) | Overall (N = 76) | K. pneumoniae (n = 47) | E. coli (n = 29) | p | ErMr (n = 47) | ErMs (n = 29) | p | CPE (n = 35) | NCPE (n = 41) | p |
| ErMs | 29 (38) | 8 (17) | 21 (72) | <0.001 | 5 (14) | 24 (59) | <0.001 | |||
| NCPE | 41 (54) | 19 (40) | 22 (76) | 0.01 | 17 (36) | 24 (83) | <0.001 | |||
| CPE | 35 (46) | 28 (60) | 7 (24) | 0.01 | 30 (64) | 5 (17) | <0.001 | |||
| blaMBL A | 5 (7) | 3 (6) | 2 (7) | 1.00 | 4 (9) | 1 (3) | 0.70 | 5 (14) | 0 (0) | 0.04 |
| blaKPC B | 28 (37) | 23 (49) | 5 (17) | 0.01 | 24 (51) | 4 (14) | 0.002 | 28 (80) | 0 (0) | <0.001 |
| blaOXA-48 C | 2 (3) | 2 (4) | 0 (0) | 0.70 | 2 (4) | 0 (0) | 0.70 | 2 (6) | 0 (0) | 0.41 |
| blaOXA-1/-9 | 33 (43) | 22 (47) | 11 (38) | 0.60 | 22 (47) | 11 (38) | 0.60 | 21 (60) | 12 (29) | 0.01 |
| blaESBL D | 52 (68) | 32 (68) | 20 (69) | 1.00 | 28 (60) | 24 (83) | 0.06 | 21 (60) | 31 (76) | 0.23 |
| blaCTX-M E | 47 (62) | 27 (57) | 20 (69) | 0.45 | 23 (49) | 24 (83) | 0.01 | 16 (46) | 31 (76) | 0.02 |
| blaCTX-M-15 | 43 (57) | 27 (57) | 16 (55) | 1.00 | 22 (47) | 21 (72) | 0.05 | 16 (46) | 27 (66) | 0.13 |
| blaSHV-12 | 7 (9) | 7 (15) | 0 (0) | 0.08 | 4 (9) | 3 (10) | 1.00 | 4 (11) | 3 (7) | 0.83 |
| blapenicillinase F | 60 (79) | 42 (89) | 18 (62) | 0.01 | 40 (85) | 20 (69) | 0.17 | 30 (86) | 30 (73) | 0.29 |
| blaAmpC G | 12 (16) | 5 (11) | 7 (24) | 0.21 | 7 (15) | 5 (17) | 1.00 | 3 (8) | 9 (22) | 0.20 |
| ID | β-Lactamase Profile A | Meropenem Hydrolysis (ng/mL-h) | Vaborbactam Hydrolysis (ng/mL-h) B | |
|---|---|---|---|---|
| Carbapenemase | Non-Carbapenemase β-Lactamase | |||
| EC68 | none | blaCMY-133, blaTEM-1 | −0.5 | −0.1 |
| EC5 | none | blaCTX-M-15, blaOXA-1 | −0.8 | No loss |
| EC201 | none | blaCTX-M-15, blaOXA-1 | −1.0 | No loss |
| KP56 | blaKPC-2 | blaOXA-9,blaTEM-1, blaSHV-182 | −1.0 | No loss |
| EC74 | blaKPC-3 | none | −1.3 | No loss |
| KP15 | blaKPC-2 | blaCTX-M-15, blaOXA-9,blaTEM-1, blaSHV-100 | −2.0 | No loss |
| EC23 | blaNDM-5 | blaCTX-M-15, blaOXA-1, blaTEM-1, blaSHV-27 | −2.8 | No loss |
| EC22 | blaNDM-5 | blaCTX-M-15, blaOXA-1, blaTEM-1, blaSHV-27 | LLQ at t1 | No loss |
| KP26 | blaNDM-1 | blaCTX-M-15, blaCMY-6, blaOXA-1, blaTEM-1, blaSHV-155 | LLQ at t1 | No loss |
| Phenotype | ID | rpsL | blaCTX-M | blaTEM | blaSHV | blaOXA-1/9 | blaCMY | blaKPC | ompC/ ompK36 | ompF/ ompK35 | tolC/ oqxA |
|---|---|---|---|---|---|---|---|---|---|---|---|
| ErMs | EC12 | 1.0 | 2.3 | 0.4 | 0.6 | 1.6 | 0.5 | 0.4 | 0.4 | ||
| EC30 | 48.8 | 2.8 | 1.4 | 1.3 | 1.7 | ||||||
| EC31 | 5.2 | 0.9 | 0.6 | 0.6 | 0.8 | ||||||
| EC35 | 39.9 | 1.2 | 0.1 | 0.7 | 0.9 | 1.0 | |||||
| KP10 | 1.0 | 10.7 | 4.7 | 0.5 | 4.3 | 0.6 | 0.7 | 0.7 | |||
| KP38 | 20.4 | 4.3 | 1.6 | 1.7 | 1.8 | 1.9 | |||||
| KP45 | 9.1 | 4.9 | 9.2 | 1.2 | 1.3 | 1.0 | |||||
| KP54 | 0.2 | 0.2 | 0.2 | 0.2 | 0.2 | 0.3 | 0.2 | ||||
| Mean ΔCt | 17.1 | 2.4 | 1.4 | 0.6 | 0.0 | 0.2 | 0.9 | 0.9 | 1.0 | ||
| EsMs | EC87 | 1.0 | 1.6 | 0.7 | 0.6 | 0.5 | 0.6 | ||||
| EC88 | 8.1 | 0.8 | 0.6 | 1.9 | |||||||
| EC89 | 2.1 | 2.0 | 0.6 | 0.7 | 1.5 | ||||||
| EC92 | 1.4 | 0.6 | 0.5 | 0.4 | 1.2 | ||||||
| KP85 | 1.0 | 1.3 | 1.2 | 0.8 | 0.5 | 0.5 | 0.5 | ||||
| KP86 | 2.7 | 0.7 | 0.3 | 0.4 | 0.6 | ||||||
| KP90 | 20 | 8.6 | 1.0 | 3.6 | 0.8 | 1.0 | 1.6 | ||||
| KP91 | 1.1 | 0.1 | 0.4 | 0.4 | |||||||
| Mean ΔCt | 4.8 | 1.5 | 0.3 | 0.7 | 0.1 | 0.0 | 0.5 | 0.6 | 1.1 | ||
| E. coli | K. pneumoniae | |||||
|---|---|---|---|---|---|---|
| CP-ErMr (n = 2) | NCP-ErMs (n = 16) | p | CP-ErMr (n = 28) | NCP-ErMs (n = 8) | p | |
| No major alteration(s) | 2 (100) | 0 (0) | 0.002 | 1 (3.6) | 0 (0) | 1.00 |
| Any major alteration(s) | 0 (0) | 16 (100) | 0.002 | 27 (96) | 8 (100) | 1.00 |
| ompC/ompK35 | 0 (0) | 14 (88) | 0.05 | 27 (96) | 8 (100) | 1.00 |
| ompF/ompK36 | 0 (0) | 8 (50) | 0.55 | 20 (71) | 4 (50) | 0.47 |
| Insertion/Deletion | 0 (0) | 10 (63) | 0.85 | 27 (96) | 8 (100) | 1.00 |
| ompC/ompK35 | 0 (0) | 10 (63) | 0.85 | 27 (96) | 8 (100) | 1.00 |
| ompF/ompK36 | 0 (0) | 0 (0) | ND | 0 (0) | 0 (0) | ND |
| Frameshift | 0 (0) | 16 (100) | 0.002 | 27 (96) | 8 (100) | 1.00 |
| ompC/ompK35 | 0 (0) | 14 (88) | 0.05 | 24 (85) | 8 (100) | 0.62 |
| ompF/ompK36 | 0 (0) | 8 (50) | 0.55 | 0 (0) | 0 (0) | ND |
| Premature Stop | 0 (0) | 1 (6.2) | 1.00 | 25 (89) | 8 (100) | 0.80 |
| ompC/ompK35 | 0 (0) | 1 (6.2) | 1.00 | 23 (82) | 7 (87) | 1.00 |
| ompF/ompK36 | 0 (0) | 0 (0) | ND | 20 (71) | 4 (50) | 0.47 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Black, C.A.; Benavides, R.; Bandy, S.M.; Dallas, S.D.; Gawrys, G.; So, W.; Moreira, A.G.; Aguilar, S.; Quidilla, K.; Smelter, D.F.; et al. Diverse Role of blaCTX-M and Porins in Mediating Ertapenem Resistance among Carbapenem-Resistant Enterobacterales. Antibiotics 2024, 13, 185. https://doi.org/10.3390/antibiotics13020185
Black CA, Benavides R, Bandy SM, Dallas SD, Gawrys G, So W, Moreira AG, Aguilar S, Quidilla K, Smelter DF, et al. Diverse Role of blaCTX-M and Porins in Mediating Ertapenem Resistance among Carbapenem-Resistant Enterobacterales. Antibiotics. 2024; 13(2):185. https://doi.org/10.3390/antibiotics13020185
Chicago/Turabian StyleBlack, Cody A., Raymond Benavides, Sarah M. Bandy, Steven D. Dallas, Gerard Gawrys, Wonhee So, Alvaro G. Moreira, Samantha Aguilar, Kevin Quidilla, Dan F. Smelter, and et al. 2024. "Diverse Role of blaCTX-M and Porins in Mediating Ertapenem Resistance among Carbapenem-Resistant Enterobacterales" Antibiotics 13, no. 2: 185. https://doi.org/10.3390/antibiotics13020185
APA StyleBlack, C. A., Benavides, R., Bandy, S. M., Dallas, S. D., Gawrys, G., So, W., Moreira, A. G., Aguilar, S., Quidilla, K., Smelter, D. F., Reveles, K. R., Frei, C. R., Koeller, J. M., & Lee, G. C. (2024). Diverse Role of blaCTX-M and Porins in Mediating Ertapenem Resistance among Carbapenem-Resistant Enterobacterales. Antibiotics, 13(2), 185. https://doi.org/10.3390/antibiotics13020185