
The Home Environment Is a Reservoir for Methicillin-Resistant Coagulase-Negative Staphylococci and Mammaliicocci

Abstract
:1. Introduction
2. Results
2.1. Isolates
2.2. Occurrence of MRCoNS and MRM
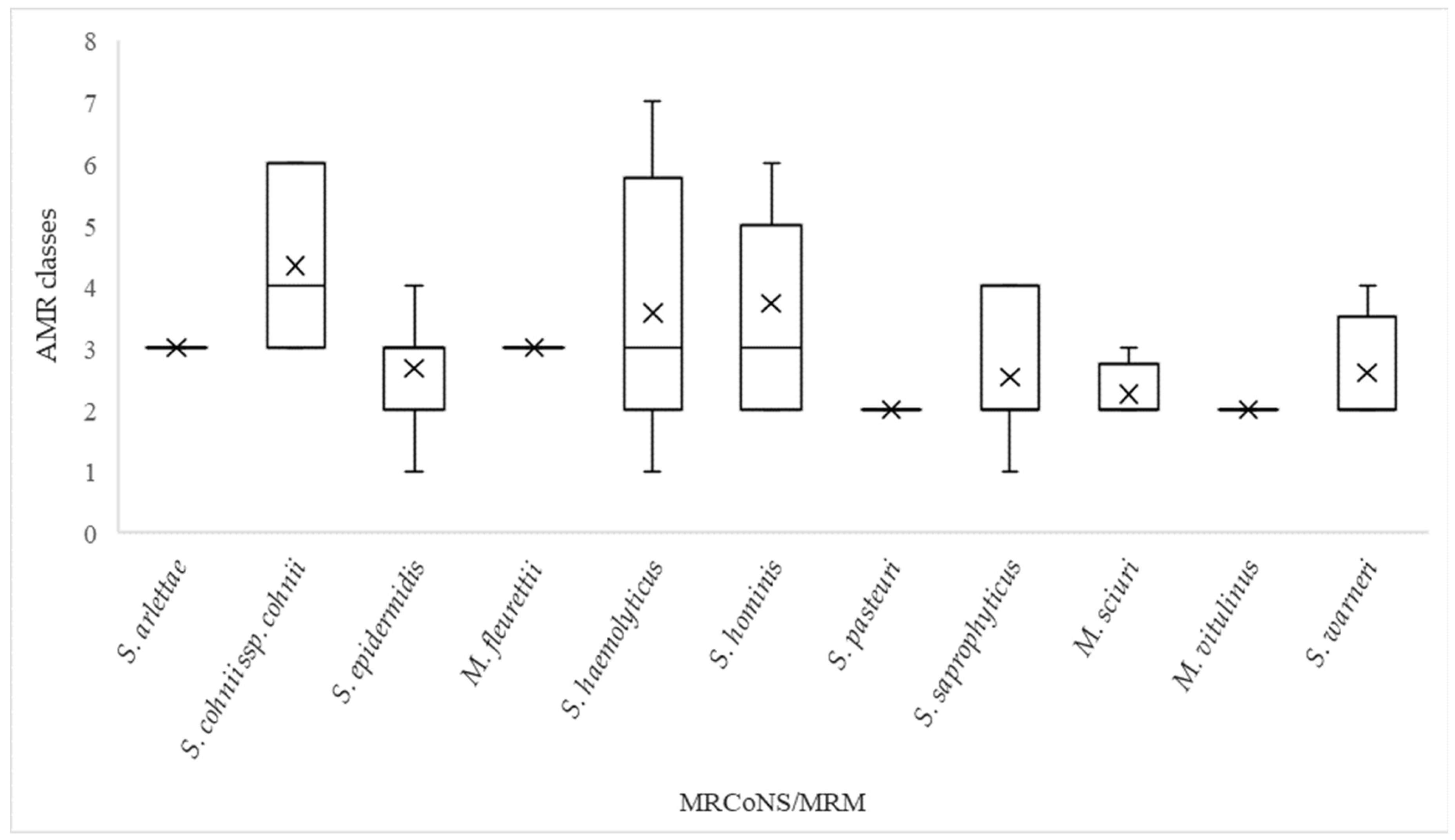
2.3. Antimicrobial Resistance
2.4. SCCmec Cassettes and Sequence Types
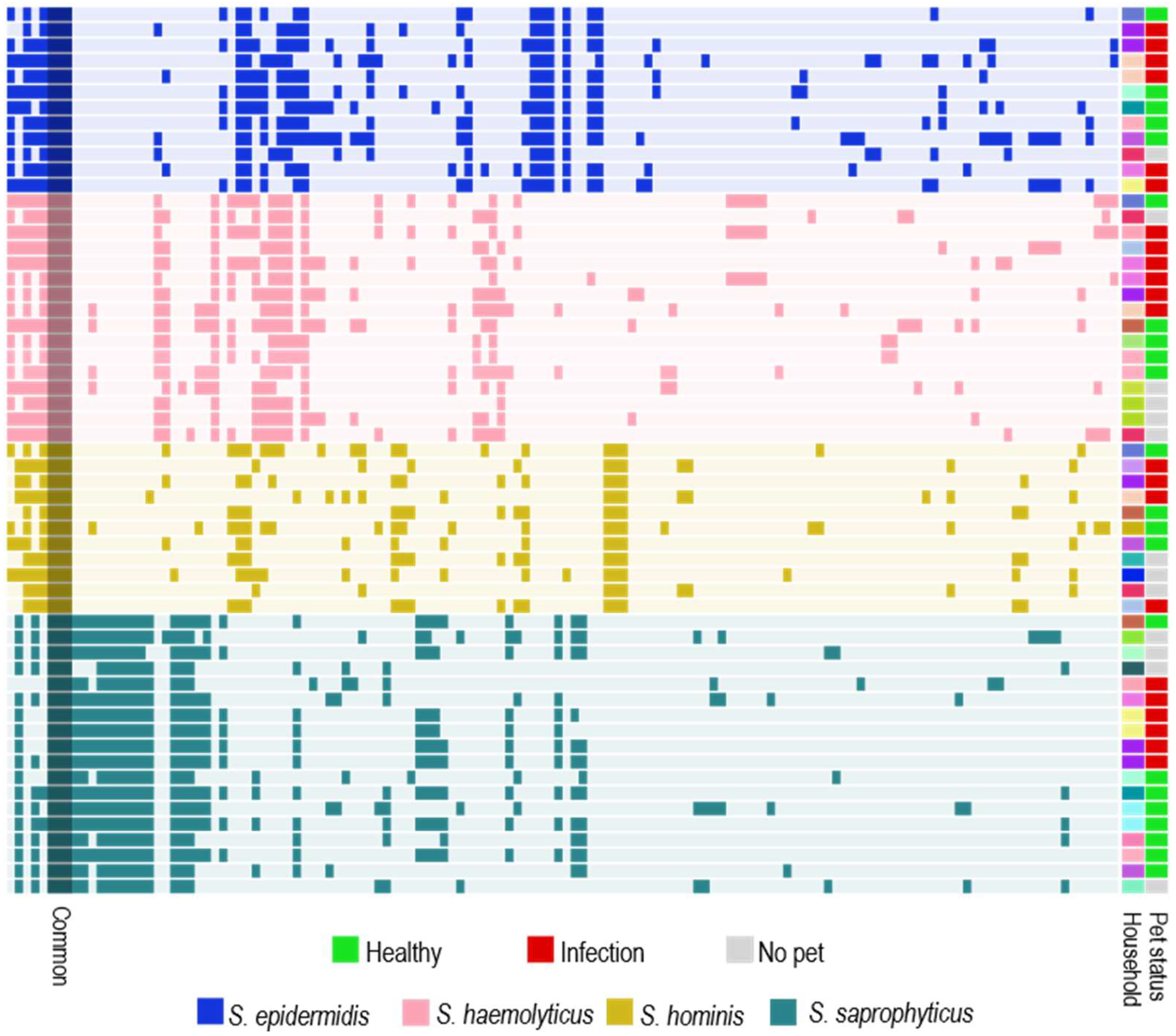
2.5. Household Analysis of Human and Pet Isolates
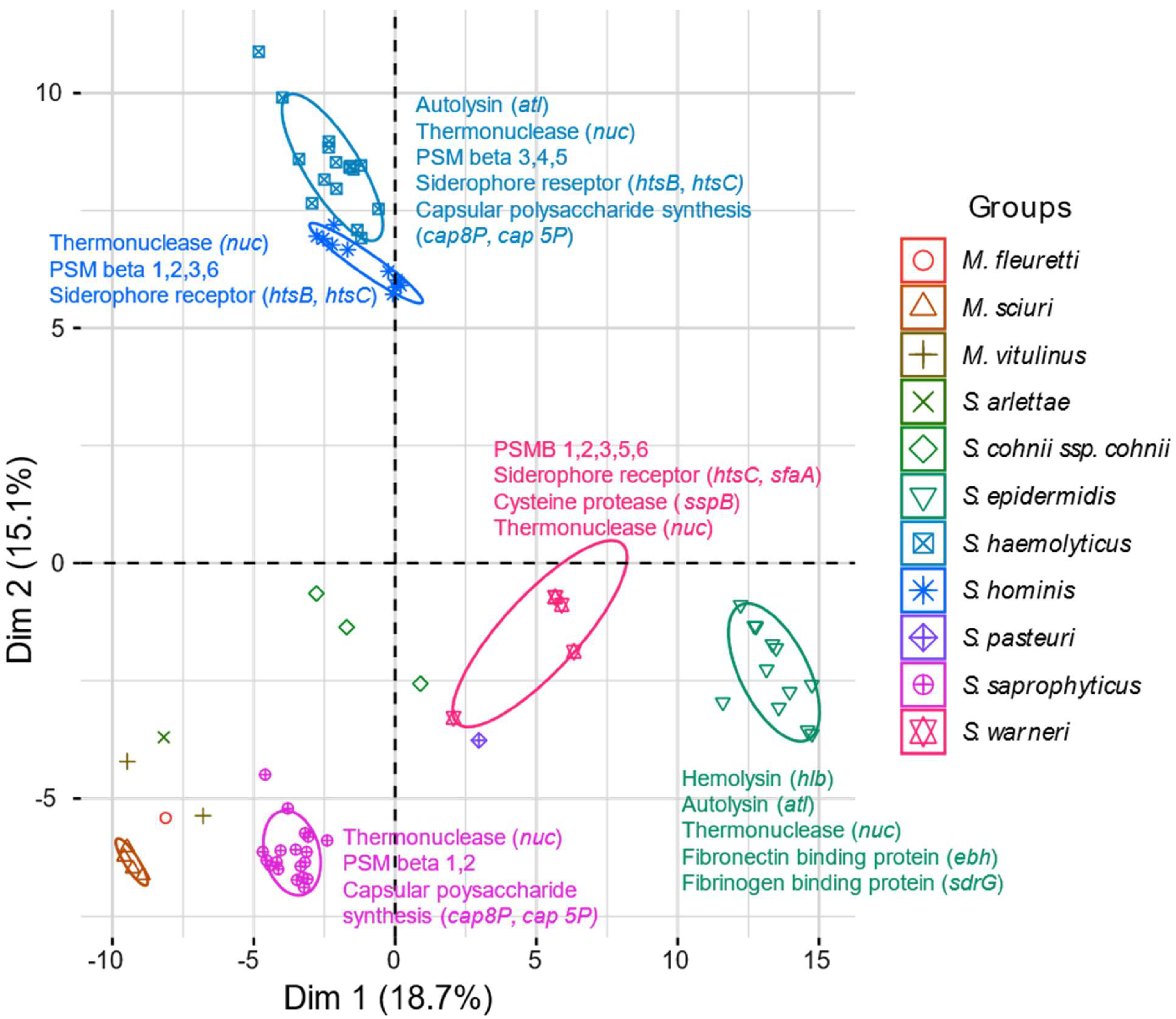
2.6. Virulence Genes
2.7. Mobilome Analysis
3. Discussion
4. Materials and Methods
4.1. Participants
4.2. Sampling
4.3. Culturing and Identification
4.4. Susceptibility Testing
4.5. Selection of Isolates
4.6. DNA Extraction, Library Preparation, Sequencing and Assembly
4.7. Bioinformatics
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Becker, K.; Heilmann, C.; Peters, G. Coagulase-Negative Staphylococci. Clin. Microbiol. Rev. 2014, 27, 870. [Google Scholar] [CrossRef]
- Piette, A.; Verschraegen, G. Role of coagulase-negative staphylococci in human disease. Vet. Microbiol. 2009, 134, 45–54. [Google Scholar] [CrossRef]
- Becker, K.; Both, A.; Weißelberg, S.; Heilmann, C.; Rohde, H. Emergence of coagulase-negative staphylococci. Expert Rev. Anti-Infect. Ther. 2020, 18, 349–366. [Google Scholar] [CrossRef]
- Hovelius, B.; Mårdh, P.-A. Staphylococcus saprophyticus as a Common Cause of Urinary Tract Infections. Rev. Infect. Dis. 1984, 6, 328–337. [Google Scholar] [CrossRef]
- Kranz, J.; Schmidt, S.; Lebert, C.; Schneidewind, L.; Mandraka, F.; Kunze, M.; Helbig, S.; Vahlensieck, W.; Naber, K.; Schmiemann, G.; et al. The 2017 Update of the German Clinical Guideline on Epidemiology, Diagnostics, Therapy, Prevention, and Management of Uncomplicated Urinary Tract Infections in Adult Patients. Part II: Therapy and Prevention. Urol. Int. 2018, 100, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.; Aarag, E.; Rønnestad, A.; Sollid, J.E.; Abrahamsen, T.G.; Kjeldsen, G.; Flægstad, T. Coagulase-Negative Staphylococcal Sepsis in Neonates: Association Between Antibiotic Resistance, Biofilm Formation and the Host Inflammatory Response. Pediatr. Infect. Dis. J. 2005, 24, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Rolo, J.; Worning, P.; Nielsen Jesper, B.; Bowden, R.; Bouchami, O.; Damborg, P.; Guardabassi, L.; Perreten, V.; Tomasz, A.; Westh, H.; et al. Evolutionary Origin of the Staphylococcal Cassette Chromosome mec (SCCmec). Antimicrob. Agents Chemother. 2017, 61, e02302-16. [Google Scholar] [CrossRef] [PubMed]
- Klingenberg, C.; Glad, T.; Olsvik, Ø.; Flægstad, T. Rapid PCR Detection of the Methicillin Resistance Gene, mecA, on the Hands of Medical and Non-medical Personnel and Healthy Children and on Surfaces in a Neonatal Intensive Care Unit. Scand. J. Infect. Dis. 2001, 33, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Mkrtchyan, H.; Cutler, R. Antibiotic resistance and mecA characterization of coagulase-negative staphylococci isolated from three hotels in London, UK. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Ćirković, I.; Djukić, S.; Vuković, D.; Švabić-Vlahović, M. Public transport as a reservoir of methicillin-resistant staphylococci. Lett. Appl. Microbiol. 2008, 47, 339–341. [Google Scholar] [CrossRef] [PubMed]
- Barbier, F.; Ruppé, E.; Hernandez, D.; Lebeaux, D.; Francois, P.; Felix, B.; Desprez, A.; Maiga, A.; Woerther, P.-L.; Gaillard, K.; et al. Methicillin-Resistant Coagulase-Negative Staphylococci in the Community: High Homology of SCCmec IVa between Staphylococcus epidermidis and Major Clones of Methicillin-Resistant Staphylococcus aureus. J. Infect. Dis. 2010, 202, 270–281. [Google Scholar] [CrossRef]
- Røken, M.; Iakhno, S.; Haaland, A.H.; Wasteson, Y.; Bjelland, A.M. Transmission of Methicillin-Resistant Staphylococcus spp. from Infected Dogs to the Home Environment and Owners. Antibiotics 2022, 11, 637. [Google Scholar] [CrossRef]
- Weiß, S.; Kadlec, K.; Feßler, A.T.; Schwarz, S. Identification and characterization of methicillin-resistant Staphylococcus aureus, Staphylococcus epidermidis, Staphylococcus haemolyticus and Staphylococcus pettenkoferi from a small animal clinic. Vet. Microbiol. 2013, 167, 680–685. [Google Scholar] [CrossRef]
- Nemeghaire, S.; Argudín, M.A.; Feßler, A.T.; Hauschild, T.; Schwarz, S.; Butaye, P. The ecological importance of the Staphylococcus sciuri species group as a reservoir for resistance and virulence genes. Vet. Microbiol. 2014, 171, 342–356. [Google Scholar] [CrossRef]
- De Vos, P.; Garrity, G.M.; Bergey, D.H. Bergey’s Manual of Systematic Bacteriology: Volume 3: The Firmicutes, 2nd ed.; Springer: New York, NY, USA, 2009; Volume 3. [Google Scholar]
- Han, J.-I.; Yang, C.-H.; Park, H.-M. Prevalence and risk factors of Staphylococcus spp. carriage among dogs and their owners: A cross-sectional study. Vet. J. 2016, 212, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Bierowiec, K.; Korzeniowska-Kowal, A.; Wzorek, A.; Rypuła, K.; Gamian, A. Prevalence of Staphylococcus Species Colonization in Healthy and Sick Cats. Biomed. Res. Int. 2019, 2019, 4360525. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shah, H.N.; Misra, R.; Chen, J.; Zhang, W.; Liu, Y.; Cutler, R.R.; Mkrtchyan, H.V. The prevalence, antibiotic resistance and mecA characterization of coagulase negative staphylococci recovered from non-healthcare settings in London, UK. Antimicrob. Resist. Infect. Control 2018, 7, 73. [Google Scholar] [CrossRef]
- Malachowa, N.; DeLeo, F.R. Mobile genetic elements of Staphylococcus aureus. Cell. Mol. Life Sci. 2010, 67, 3057–3071. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. Breakpoint Tables for Interpretation of MICs and Zone Diameters Version 12.0. Available online: https://www.eucast.org/clinical_breakpoints/ (accessed on 10 May 2022).
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 32nd ed.; CLSI supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2022; pp. 66–77. [Google Scholar]
- Cosimi, R.A.; Beik, N.; Kubiak, D.W.; Johnson, J.A. Ceftaroline for Severe Methicillin-Resistant Staphylococcus aureus Infections: A Systematic Review. Open Forum Infect. Dis. 2017, 4, ofx084. [Google Scholar] [CrossRef]
- Stein, G.E.; Johnson, L.B. Ceftaroline: A Novel Cephalosporin with Activity against Methicillin-resistant Staphylococcus aureus. Clin. Infect. Dis. 2011, 52, 1156–1163. [Google Scholar] [CrossRef]
- Chen, X.-P.; Li, W.-G.; Zheng, H.; Du, H.-Y.; Zhang, L.; Zhang, L.; Che, J.; Wu, Y.; Liu, S.-M.; Lu, J.-X. Extreme diversity and multiple SCCmec elements in coagulase-negative Staphylococcus found in the Clinic and Community in Beijing, China. Ann. Clin. Microbiol. Antimicrob. 2017, 16, 57. [Google Scholar] [CrossRef]
- Gómez-Sanz, E.; Ceballos, S.; Ruiz-Ripa, L.; Zarazaga, M.; Torres, C. Clonally Diverse Methicillin and Multidrug Resistant Coagulase Negative Staphylococci Are Ubiquitous and Pose Transfer Ability between Pets and Their Owners. Front. Microbiol. 2019, 10. [Google Scholar] [CrossRef] [PubMed]
- Naushad, S.; Naqvi, S.A.; Nobrega, D.; Luby, C.; Kastelic John, P.; Barkema Herman, W.; De Buck, J.; Kent Angela, D. Comprehensive Virulence Gene Profiling of Bovine Non-aureus Staphylococci Based on Whole-Genome Sequencing Data. mSystems 2019, 4, e00098-18. [Google Scholar] [CrossRef]
- Stegger, M.; Andersen, P.S.; Kearns, A.; Pichon, B.; Holmes, M.A.; Edwards, G.; Laurent, F.; Teale, C.; Skov, R.; Larsen, A.R. Rapid detection, differentiation and typing of methicillin-resistant Staphylococcus aureus harbouring either mecA or the new mecA homologue mecALGA251. Clin. Microbiol. Infect. 2012, 18, 395–400. [Google Scholar] [CrossRef] [PubMed]
- EUCAST. EUCAST Disk Diffusion Method for Antimicrobial Susceptibility Testing; EUCAST: Basel, Switzerland, 2019. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Rodriguez-R, L.M.; Harvey, W.T.; Rosselló-Mora, R.; Tiedje, J.M.; Cole, J.R.; Konstantinidis, K.T. Classifying prokaryotic genomes using the Microbial Genomes Atlas (MiGA) webserver. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley and Sons: Hoboken, NJ, USA, 2018; pp. 1–11. [Google Scholar]
- Krueger, F.; Galore, T. A Wrapper Tool around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Filesl; Babraham Institute: Cambridge, UK, 2015; Volume 516. [Google Scholar]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. Abricate: Mass Screening of Contigs for Antimicrobial and Virulence Genes; Department of Microbiology and Immunology, The University of Melbourne: Melbourne, Australia, 2018; Available online: https://github.com/tseemann/abricate (accessed on 28 February 2019).
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Hasman, H.; Larsen, J.; Stegger, M.; Johannesen, T.B.; Allesøe, R.L.; Lemvigh, C.K.; Aarestrup, F.M.; Lund, O.; Larsen, A.R. SCCmecFinder, a Web-Based Tool for Typing of Staphylococcal Cassette Chromosome mec in Staphylococcus aureus Using Whole-Genome Sequence Data. mSphere 2018, 3, e00612-17. [Google Scholar] [CrossRef] [PubMed]
- Seemann, T. MLST. Available online: https://github.com/tseemann/mlst (accessed on 9 February 2022).
- Eren, A.M.; Esen, Ö.C.; Quince, C.; Vineis, J.H.; Morrison, H.G.; Sogin, M.L.; Delmont, T.O. Anvi’o: An advanced analysis and visualization platform for ‘omics data. PeerJ 2015, 3, e1319. [Google Scholar] [CrossRef]
- Hyatt, D.; Chen, G.-L.; LoCascio, P.F.; Land, M.L.; Larimer, F.W.; Hauser, L.J. Prodigal: Prokaryotic gene recognition and translation initiation site identification. BMC Bioinform. 2010, 11, 119. [Google Scholar] [CrossRef]
- Tatusov, R.L.; Galperin, M.Y.; Natale, D.A.; Koonin, E.V. The COG database: A tool for genome-scale analysis of protein functions and evolution. Nucleic Acids Res. 2000, 28, 33–36. [Google Scholar] [CrossRef] [PubMed]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using DIAMOND. Nat. Methods 2015, 12, 59–60. [Google Scholar] [CrossRef] [PubMed]
- van Dongen, S.; Abreu-Goodger, C. Using MCL to extract clusters from networks. Methods Mol. Biol. 2012, 804, 281–295. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Status | Household | Pet AM Treatment (within Months) | Human AM Treatment (within 12 Months) | Hospital Admission (within 12 Months) | Work in Health Care | Home Environment | Human Carriage | Pet Carriage | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. arlettae | S. cohnii ssp. cohnii | S. epidermidis | M. fleurettii | S. haemolyticus | S. hominis | S. pasteuri | S. saprophyticus | M. sciuri | M. vitulinus | S. warneri | ||||||||
| Household with dogs with infection | A | Yes | B, FB, K | FB, K | B, FL, K | |||||||||||||
| B | Pet | B, FL K | ||||||||||||||||
| C | Cefalexin (0–3) Polymyxin B (3–6) | Unknown agent | B, K | FB, FL | B | FB, S | S. epidermidis | S. epidermidis | ||||||||||
| D | Cefalexin (0–3) | Pet | S | B, FL, K, S | B | FL, S | S. haemolyticus | |||||||||||
| E | Amoxicillin (0–3) Trim/sulfa (0–3) | Pet | B | K | FB | FL | ||||||||||||
| F | Amoxicillin 0–3) Cefalexin (3–6) Enrofloxacin (3–6) | Pet. Human | Yes | K, S | B | B | FB | FL, K | FL, S | S. epidermidis | S. epidermidis | |||||||
| G | Pet | FL | ||||||||||||||||
| H | Amoxicillin (0–3) Fusidic acid (6–12) | K | FB, K | S. epidermidis | S. epidermidis | |||||||||||||
| Household with healthy dogs | 1 | FL, S | FB | FL, K | ||||||||||||||
| 2 | ||||||||||||||||||
| 3 | Clindamycin | Human | S | K | S. warneri | |||||||||||||
| 4 | Amoxicillin–clav (0–3) | Penicillin | S | |||||||||||||||
| 5 | Fusidic acid (0–3) | Chloramphenicol | Pet | Yes | B, FL, S | |||||||||||||
| 6 | ||||||||||||||||||
| 7 | Human | B | FB, FL | |||||||||||||||
| 8 | Erythromycin | Human | B, FB, FL, S, K | B | FL | FB | ||||||||||||
| Household with heathy cats | 9 | Pivemecillinam | B, FL | K | B, FB, S | |||||||||||||
| 10 | Amoxicillin (0–3) | FB | FL | |||||||||||||||
| 11 | FB | FB, S | ||||||||||||||||
| 12 | FL | |||||||||||||||||
| 13 | FL, S | |||||||||||||||||
| 14 | B | FL | FL | S. haemolyticus | ||||||||||||||
| Household without pets | I | Yes | B | |||||||||||||||
| II | Yes | FL, K | ||||||||||||||||
| III | B | K | ||||||||||||||||
| IV | Penicillin | FL | B, K | |||||||||||||||
| V | FL | |||||||||||||||||
| VI | Yes | B | ||||||||||||||||
| VII | Penicillin | |||||||||||||||||
| VIII | Human | B, FL | FL | |||||||||||||||
| IX | Unknown agent | Human | K | |||||||||||||||
| X | FL, K | |||||||||||||||||
| XI | Tetracycline | K | FL, K | FL | K | B | ||||||||||||
| Antimicrobial Agent | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species | n | Gen | Chl | Oxa | Cfox | AmCl | Clex | Enr | T/S | Fus | Cli | Ery | Tet |
| All isolates | 75 | 20 | 5 | 99 | 88 | 73 | 95 | 12 | 21 | 51 | 21 | 51 | 20 |
| S. arlettae | 1 | 100 | 100 | 100 | 100 | 100 | 100 | ||||||
| S. cohnii ssp. cohnii | 3 | 33 | 100 | 100 | 33 | 100 | 33 | 33 | 67 | 100 | 67 | ||
| S. epidermidis | 12 | 8 | 100 | 100 | 33 | 83 | 8 | 17 | 50 | 17 | 50 | 17 | |
| M. fleuretti | 1 | 100 | 100 | 100 | 100 | 100 | 100 | ||||||
| S. haemolyticus | 16 | 50 | 6 | 100 | 88 | 88 | 100 | 38 | 25 | 25 | 31 | 50 | 31 |
| S. hominis | 11 | 46 | 91 | 55 | 46 | 82 | 18 | 46 | 55 | 27 | 64 | 27 | |
| S. pasteuri | 1 | 100 | 100 | 100 | 100 | 100 | |||||||
| S. saprophyticus | 19 | 11 | 100 | 95 | 100 | 100 | 16 | 53 | 5 | 58 | 11 | ||
| M. sciuri | 4 | 100 | 100 | 100 | 100 | 100 | 25 | ||||||
| M. vitulinus | 2 | 100 | 50 | 50 | 100 | 100 | |||||||
| S. warneri | 5 | 20 | 100 | 100 | 80 | 80 | 20 | 80 | 20 | 20 | |||
| Antimicrobial Class | ID | S. arlettae | S. cohnii ssp. cohnii | S. epidermidis | M. fleurettii | S. haemolyticus | S. hominis | S. pasteuri | S. saprophyticus | M. sciuri | M. vitulinus | S. warneri | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ARG | All Isolates n = 75 | 1 | 3 | 12 | 1 | 16 | 11 | 1 | 19 | 4 | 2 | 5 | |
| Aminoglycoside | aac(6′)-le/aph(2″)-la | 20 | 8 | 56 | 46 | ||||||||
| ant(4′)-lb | 13 | 67 | 25 | 31 | |||||||||
| aph(3′)-IIIa | 4 | 19 | |||||||||||
| sat4 | 5 | 8 | 19 | ||||||||||
| Amphenicol | cat | 4 | 33 | 11 | |||||||||
| catA | 1 | 6 | |||||||||||
| Beta-lactam | arl | 1 | 100 | ||||||||||
| blaZ | 63 | 33 | 100 | 94 | 82 | 32 | 80 | ||||||
| mecA | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | |
| Folate pathway antagonist | dfrC | 27 | 92 | 9 | 21 | 80 | |||||||
| dfrG | 8 | 8 | 31 | ||||||||||
| Fosfomycin | fosB6 | 4 | 27 | ||||||||||
| fosD | 1 | 25 | |||||||||||
| Fusidane | fusB | 29 | 50 | 25 | 36 | 21 | 80 | ||||||
| fusC | 7 | 8 | 6 | 27 | |||||||||
| fusD | 25 | 100 | |||||||||||
| fusF | 4 | 100 | |||||||||||
| Macrolide, lincosamide, streptogramin | ermC | 13 | 8 | 31 | 18 | 5 | 20 | ||||||
| lnuA | 15 | 67 | 27 | 32 | |||||||||
| mphC | 29 | 100 | 17 | 38 | 9 | 100 | 47 | ||||||
| msrA | 43 | 100 | 100 | 42 | 38 | 42 | 100 | 46 | 20 | ||||
| salA | 5 | 100 | |||||||||||
| vgaA | 4 | 8 | 9 | ||||||||||
| vgaALC | 5 | 19 | |||||||||||
| Multidrug | mgrA | 61 | 33 | 17 | 100 | 92 | 100 | 100 | |||||
| norA | 24 | 100 | 8 | 100 | 80 | ||||||||
| Pseudomonic acid | mupA | 5 | 13 | 17 | |||||||||
| Quaternary ammonium compounds | qacA | 41 | 33 | 42 | 69 | 91 | 11 | 40 | |||||
| qacB | 3 | 33 | 100 | ||||||||||
| Tetracycline | tetK | 20 | 100 | 67 | 17 | 38 | 18 | 11 | |||||
| tetL | 3 | 13 |
| Species ID | II (2A) | III (3A) | IV (2B) | IVa (2B) | IVd (2B) | IVc (2B) | V (5C2 and 5) | VIII (4A) | Non-Typeable |
|---|---|---|---|---|---|---|---|---|---|
| S. arlettae | 1 | ||||||||
| S. cohnii ssp. cohnii | 3 | ||||||||
| S. epidermidis | 1 | 1 | 2 | 6 | 1 | 1 | |||
| M. fleurettii | 1 | ||||||||
| S. haemolyticus | 1 | 1 | 14 | ||||||
| S. hominis | 4 | 7 | |||||||
| S. pasteuri | 1 | ||||||||
| S. saprophyticus | 10 | 9 | |||||||
| M. sciuri | 4 | ||||||||
| M. vitulinus | 2 | ||||||||
| S. warneri | 3 | 1 | 1 |
| Analysis | Isolates Included in the Analysis | Isolates Presented in the Results Section |
|---|---|---|
| mecA PCR | All cultured isolates | |
| Species identification (MALDI-TOF MS) | All cultured isolates | |
| Susceptibility testing | All cultured isolates | Non-redundant WGS isolates |
| Whole-genome sequencing (WGS) | Non-redundant isolates based on phenotypical resistance profiles, species and household | |
| Additional species identification (in silico, TypeMat) | All WGS isolates | |
| Resistome analysis | All WGS isolates | Non-redundant WGS isolates |
| Virulence gene analysis | All WGS isolates | Non-redundant WGS isolates |
| SCCmec typing | All WGS isolates | Non-redundant WGS isolates |
| Sequence typing (in silico MLST) | All WGS S. epidermidis, S. hominis and S. haemolyticus isolates | Non-redundant WGS S. epidermidis, S. hominis and S. haemolyticus isolates |
| Mobilome analysis | All whole genome-sequenced S. epidermidis, S. haemolyticus, S. hominis and S. saprophyticus isolates | Non-redundant S. epidermidis, S. haemolyticus, S. hominis and S. saprophyticus isolates |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Røken, M.; Iakhno, S.; Haaland, A.H.; Bjelland, A.M.; Wasteson, Y. The Home Environment Is a Reservoir for Methicillin-Resistant Coagulase-Negative Staphylococci and Mammaliicocci. Antibiotics 2024, 13, 279. https://doi.org/10.3390/antibiotics13030279
Røken M, Iakhno S, Haaland AH, Bjelland AM, Wasteson Y. The Home Environment Is a Reservoir for Methicillin-Resistant Coagulase-Negative Staphylococci and Mammaliicocci. Antibiotics. 2024; 13(3):279. https://doi.org/10.3390/antibiotics13030279
Chicago/Turabian StyleRøken, Mari, Stanislav Iakhno, Anita Haug Haaland, Ane Mohn Bjelland, and Yngvild Wasteson. 2024. "The Home Environment Is a Reservoir for Methicillin-Resistant Coagulase-Negative Staphylococci and Mammaliicocci" Antibiotics 13, no. 3: 279. https://doi.org/10.3390/antibiotics13030279
APA StyleRøken, M., Iakhno, S., Haaland, A. H., Bjelland, A. M., & Wasteson, Y. (2024). The Home Environment Is a Reservoir for Methicillin-Resistant Coagulase-Negative Staphylococci and Mammaliicocci. Antibiotics, 13(3), 279. https://doi.org/10.3390/antibiotics13030279