Cytokine Expression Patterns and Single Nucleotide Polymorphisms (SNPs) in Patients with Chronic Borreliosis

Abstract
:1. Introduction
2. Results
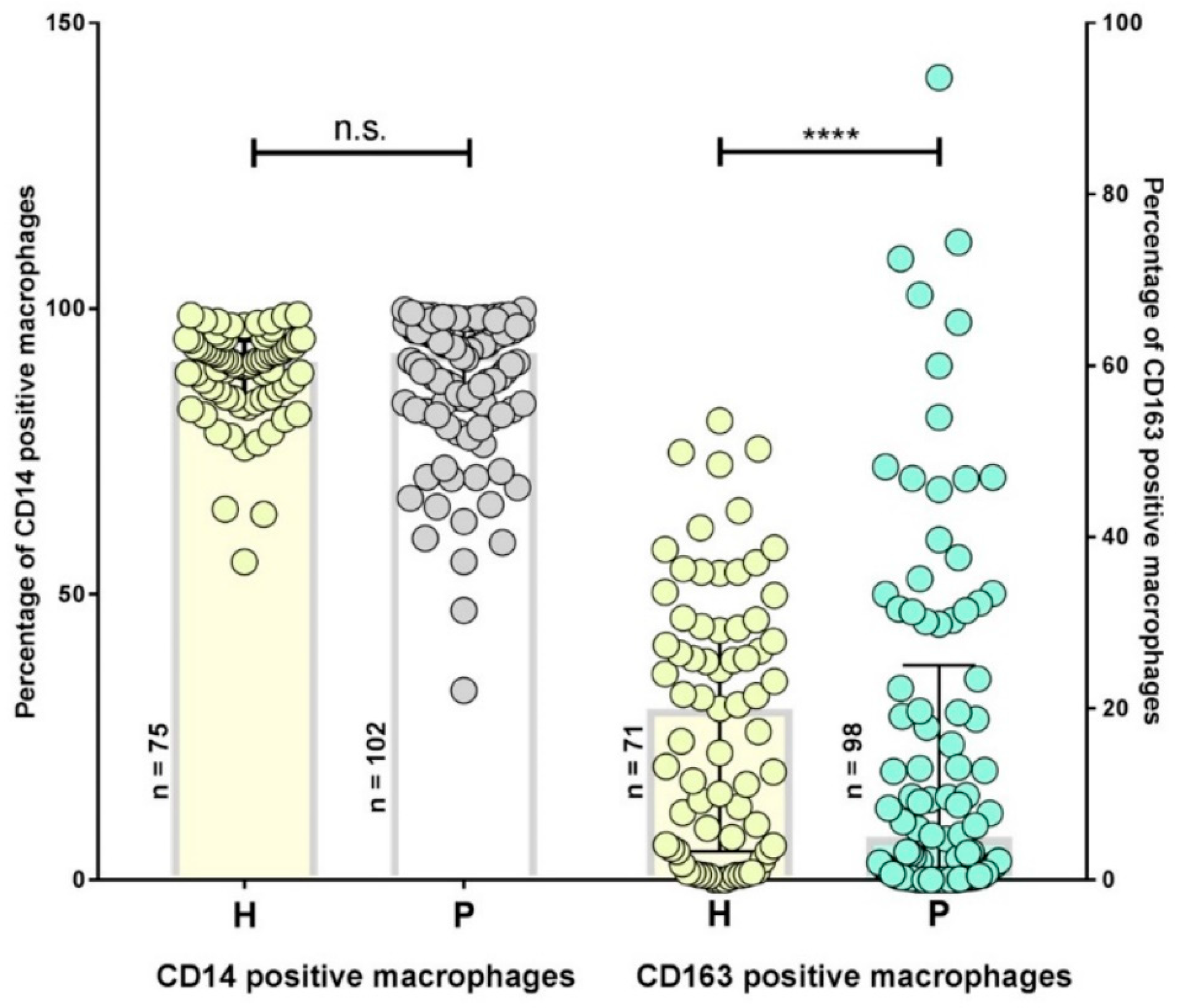
2.1. Inflammatory Phenotypes in Patients with Persistent Lyme Disease
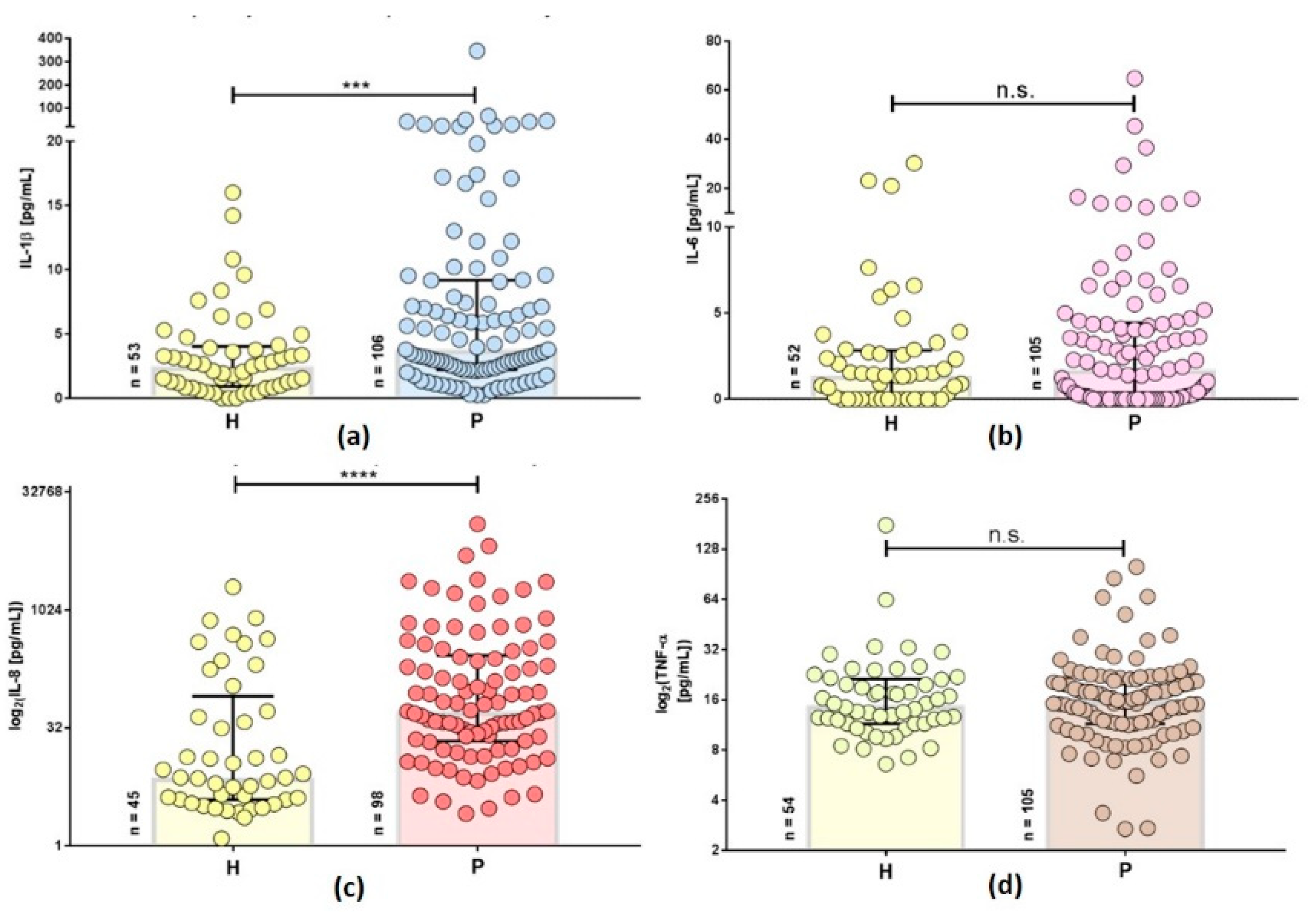
2.2. Inflammatory Cytokines are Increased in Persistent Lyme Borreliosis Patients
2.3. Inflammatory Cytokines Differ in Secondary Diseases of Patients with Persistent Lyme Disease
2.4. Allele Distribution of IL-6 Promotor rs1800795 in Patients with Persistent Lyme Disease
2.5. Different IL-6 rs1800795 Genotypes were Associated with a Decreased Number of CD163+ Macrophages and Elevated Cytokine Levels
3. Discussion
4. Materials and Methods
4.1. Ethics
4.2. Patients and Controls
4.3. Flow Cytometry for Phenotyping Patient PBMCs
4.4. Plasma Biomarkers
4.5. DNA Isolation
4.6. Pyrosequencing
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Steere, A.C.; Strle, F.; Wormser, G.P.; Hu, L.T.; Branda, J.A.; Hovius, J.W.; Li, X.; Mead, P.S. Lyme borreliosis. Nat. Rev. Dis. Primers 2016, 2, 16090. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.L.; Leydet, B.F.; Threlkeld, C. Geographical and genospecies distribution of Borrelia burgdorferi sensu lato DNA detected in humans in the USA. J. Med. Microbiol. 2014, 63, 674–684. [Google Scholar] [CrossRef] [PubMed]
- Peacock, B.N.; Gherezghiher, T.B.; Hilario, J.D.; Kellermann, G.H. New insights into Lyme disease. Redox Biol. 2015, 5, 66–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Girschick, H.J.; Morbach, H.; Tappe, D. Treatment of Lyme borreliosis. Arthritis Res. 2009, 11, 258. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, E.D. Lyme Disease. New Engl. J. Med. 2014, 370, 1724–1731. [Google Scholar] [CrossRef] [Green Version]
- Ścieszka, J.; Dąbek, J.; Cieślik, P. Post-Lyme disease syndrome. Reumatologia 2015, 53, 46–48. [Google Scholar] [CrossRef] [Green Version]
- Aucott, J.N.; Rebman, A.W.; Crowder, L.A.; Kortte, K.B. Post-treatment Lyme disease syndrome symptomatology and the impact on life functioning: Is there something here? Qual. Life Res. Int. J. Qual. Life Asp. Treat. Care Rehabil. 2013, 22, 75–84. [Google Scholar] [CrossRef]
- Embers, M.E.; Barthold, S.W.; Borda, J.T.; Bowers, L.; Doyle, L.; Hodzic, E.; Jacobs, M.B.; Hasenkampf, N.R.; Martin, D.S.; Narasimhan, S.; et al. Persistence of Borrelia burgdorferi in Rhesus Macaques following Antibiotic Treatment of Disseminated Infection. PLoS ONE 2012, 7, e29914. [Google Scholar] [CrossRef]
- Feng, J.; Shi, W.; Zhang, S.; Sullivan, D.; Auwaerter, P.G.; Zhang, Y. A Drug Combination Screen Identifies Drugs Active against Amoxicillin-Induced Round Bodies of In Vitro Borrelia burgdorferi Persisters from an FDA Drug Library. Front. Microbiol. 2016, 7, 743. [Google Scholar] [CrossRef] [Green Version]
- Sapi, E.; Balasubramanian, K.; Poruri, A.; Maghsoudlou, J.S.; Socarras, K.M.; Timmaraju, A.V.; Filush, K.R.; Gupta, K.; Shaikh, S.; Theophilus, P.A.; et al. Evidence of In Vivo Existence of Borrelia Biofilm in Borrelial Lymphocytomas. Eur. J. Microbiol. Immunol. 2016, 6, 9–24. [Google Scholar] [CrossRef]
- Glickstein, L.; Moore, B.; Bledsoe, T.; Damle, N.; Sikand, V.; Steere, A.C. Inflammatory cytokine production predominates in early Lyme disease in patients with erythema migrans. Infect. Immun. 2003, 71, 6051–6053. [Google Scholar] [CrossRef] [PubMed]
- Oosting, M.; Berende, A.; Sturm, P.; Ter Hofstede, H.J.; de Jong, D.J.; Kanneganti, T.D.; van der Meer, J.W.; Kullberg, B.J.; Netea, M.G.; Joosten, L.A. Recognition of Borrelia burgdorferi by NOD2 is central for the induction of an inflammatory reaction. J. Infect. Dis. 2010, 201, 1849–1858. [Google Scholar] [CrossRef] [PubMed]
- Oosting, M.; Ter Hofstede, H.; Sturm, P.; Adema, G.J.; Kullberg, B.J.; van der Meer, J.W.; Netea, M.G.; Joosten, L.A. TLR1/TLR2 heterodimers play an important role in the recognition of Borrelia spirochetes. PLoS ONE 2011, 6, e25998. [Google Scholar] [CrossRef] [PubMed]
- Hirschfeld, M.; Kirschning, C.J.; Schwandner, R.; Wesche, H.; Weis, J.H.; Wooten, R.M.; Weis, J.J. Cutting edge: Inflammatory signaling by Borrelia burgdorferi lipoproteins is mediated by toll-like receptor 2. J. Immunol. (Baltimore MD 1950) 1999, 163, 2382–2386. [Google Scholar]
- Parthasarathy, G.; Philipp, M.T. The MEK/ERK pathway is the primary conduit for Borrelia burgdorferi-induced inflammation and P53-mediated apoptosis in oligodendrocytes. Apoptosis 2014, 19, 76–89. [Google Scholar] [CrossRef] [PubMed]
- Giambartolomei, G.H.; Dennis, V.A.; Lasater, B.L.; Philipp, M.T. Induction of pro- and anti-inflammatory cytokines by Borrelia burgdorferi lipoproteins in monocytes is mediated by CD14. Infect. Immun. 1999, 67, 140–147. [Google Scholar] [PubMed]
- Jablonska, E.; Marcinczyk, M. TLR2 expression in relation to IL-6 and IL-1beta and their natural regulators production by PMN and PBMC in patients with Lyme disease. Mediat. Inflamm. 2006, 2006, 32071. [Google Scholar] [CrossRef] [PubMed]
- Hodzic, E.; Feng, S.; Barthold, S.W. Assessment of transcriptional activity of Borrelia burgdorferi and host cytokine genes during early and late infection in a mouse model. Vector Borne Zoonotic Dis. (Larchmt. N. Y.) 2013, 13, 694–711. [Google Scholar] [CrossRef] [PubMed]
- Meddeb, M.; Carpentier, W.; Cagnard, N.; Nadaud, S.; Grillon, A.; Barthel, C.; De Martino, S.J.; Jaulhac, B.; Boulanger, N.; Schramm, F. Homogeneous Inflammatory Gene Profiles Induced in Human Dermal Fibroblasts in Response to the Three Main Species of Borrelia burgdorferi sensu lato. PLoS ONE 2016, 11, e0164117. [Google Scholar] [CrossRef] [PubMed]
- Schindler, R.; Mancilla, J.; Endres, S.; Ghorbani, R.; Clark, S.C.; Dinarello, C.A. Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990, 75, 40–47. [Google Scholar] [PubMed]
- Bernardino, A.L.F.; Myers, T.A.; Alvarez, X.; Hasegawa, A.; Philipp, M.T. Toll-like receptors: Insights into their possible role in the pathogenesis of lyme neuroborreliosis. Infect. Immun. 2008, 76, 4385–4395. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, J.L.; Dunham-Ems, S.M.; La Vake, C.J.; Petzke, M.M.; Sahay, B.; Sellati, T.J.; Radolf, J.D.; Salazar, J.C. Phagosomal signaling by Borrelia burgdorferi in human monocytes involves Toll-like receptor (TLR) 2 and TLR8 cooperativity and TLR8-mediated induction of IFN-beta. Proc. Natl. Acad. Sci. USA 2011, 108, 3683–3688. [Google Scholar] [CrossRef] [PubMed]
- Cervantes, J.L.; La Vake, C.J.; Weinerman, B.; Luu, S.; O’Connell, C.; Verardi, P.H.; Salazar, J.C. Human TLR8 is activated upon recognition of Borrelia burgdorferi RNA in the phagosome of human monocytes. J. Leukoc. Biol. 2013, 94, 1231–1241. [Google Scholar] [CrossRef] [PubMed]
- Love, A.C.; Schwartz, I.; Petzke, M.M. Borrelia burgdorferi RNA induces type I and III interferons via Toll-like receptor 7 and contributes to production of NF-kappaB-dependent cytokines. Infect. Immun. 2014, 82, 2405–2416. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Seemanapalli, S.V.; Reif, K.E.; Brown, C.R.; Liang, F.T. Increasing the recruitment of neutrophils to the site of infection dramatically attenuates Borrelia burgdorferi infectivity. J. Immunol. (Baltimore MD 1950) 2007, 178, 5109–5115. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, R.R.; Nathanson, M.H.; Malawista, S.E. The fate of Borrelia burgdorferi, the agent for Lyme disease, in mouse macrophages. Destruction, survival, recovery. J. Immunol. (Baltimore MD 1950) 1993, 150, 909–915. [Google Scholar]
- Hodzic, E.; Feng, S.; Holden, K.; Freet, K.J.; Barthold, S.W. Persistence of Borrelia burgdorferi following antibiotic treatment in mice. Antimicrob. Agents Chemother. 2008, 52, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Brisson, D. Interactions between host immune response and antigenic variation that control Borrelia burgdorferi population dynamics. Microbiol. (Read. Engl.) 2017, 163, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Berndtson, K. Review of evidence for immune evasion and persistent infection in Lyme disease. Int. J. Gen. Med. 2013, 6, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Schoen, R.T.; Taylor, E. The clinical evolution of Lyme arthritis. Ann. Intern. Med. 1987, 107, 725–731. [Google Scholar] [CrossRef]
- Steere, A.C.; Glickstein, L. Elucidation of Lyme arthritis. Nat. Rev. Immunol. 2004, 4, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Mullegger, R.R.; Glatz, M. Skin manifestations of lyme borreliosis: Diagnosis and management. Am. J. Clin. Dermatol. 2008, 9, 355–368. [Google Scholar] [CrossRef] [PubMed]
- Biesiada, G.; Czepiel, J.; Lesniak, M.R.; Garlicki, A.; Mach, T. Lyme disease: Review. Arch. Med. Sci. AMS 2012, 8, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Halperin, J.J.; Little, B.W.; Coyle, P.K.; Dattwyler, R.J. Lyme disease: Cause of a treatable peripheral neuropathy. Neurology 1987, 37, 1700–1706. [Google Scholar] [CrossRef] [PubMed]
- Nau, R.; Christen, H.-J.; Eiffert, H. Lyme disease—Current state of knowledge. Dtsch. Arztebl. Int. 2009, 106, 72. [Google Scholar] [CrossRef]
- Piche-Renaud, P.P.; Branson, H.; Yeh, E.A.; Morris, S.K. Lyme disease presenting with multiple cranial neuropathies on MRI. IDCases 2018, 12, 117–118. [Google Scholar] [CrossRef]
- Terekhova, D.; Sartakova, M.L.; Wormser, G.P.; Schwartz, I.; Cabello, F.C. Erythromycin resistance in Borrelia burgdorferi. Antimicrob. Agents. Chemother. 2002, 46, 3637–3640. [Google Scholar] [CrossRef]
- Fishman, D.; Faulds, G.; Jeffery, R.; Mohamed-Ali, V.; Yudkin, J.S.; Humphries, S.; Woo, P. The effect of novel polymorphisms in the interleukin-6 (IL-6) gene on IL-6 transcription and plasma IL-6 levels, and an association with systemic-onset juvenile chronic arthritis. J. Clin. Investig. 1998, 102, 1369–1376. [Google Scholar] [CrossRef]
- Jenny Nancy, S.; Tracy Russell, P.; Ogg Malcolm, S.; Luong Le, A.; Kuller Lewis, H.; Arnold Alice, M.; Sharrett, A.R.; Humphries Steve, E. In the Elderly, Interleukin-6 Plasma Levels and the −174G>C Polymorphism Are Associated with the Development of Cardiovascular Disease. Arterioscler. Thromb. Vasc. Biol. 2002, 22, 2066–2071. [Google Scholar] [CrossRef]
- Antonicelli, R.; Olivieri, F.; Bonafe, M.; Cavallone, L.; Spazzafumo, L.; Marchegiani, F.; Cardelli, M.; Recanatini, A.; Testarmata, P.; Boemi, M.; et al. The interleukin-6 -174 G>C promoter polymorphism is associated with a higher risk of death after an acute coronary syndrome in male elderly patients. Int. J. Cardiol. 2005, 103, 266–271. [Google Scholar] [CrossRef]
- Illig, T.; Bongardt, F.; Schopfer, A.; Muller-Scholze, S.; Rathmann, W.; Koenig, W.; Thorand, B.; Vollmert, C.; Holle, R.; Kolb, H.; et al. Significant association of the interleukin-6 gene polymorphisms C-174G and A-598G with type 2 diabetes. J. Clin. Endocrinol. Metab. 2004, 89, 5053–5058. [Google Scholar] [CrossRef] [PubMed]
- Vaxillaire, M.; Veslot, J.; Dina, C.; Proenca, C.; Cauchi, S.; Charpentier, G.; Tichet, J.; Fumeron, F.; Marre, M.; Meyre, D.; et al. Impact of common type 2 diabetes risk polymorphisms in the DESIR prospective study. Diabetes 2008, 57, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Tan, D.; Wu, X.; Hou, M.; Lee, S.O.; Lou, W.; Wang, J.; Janarthan, B.; Nallapareddy, S.; Trump, D.L.; Gao, A.C. Interleukin-6 polymorphism is associated with more aggressive prostate cancer. J. Urol. 2005, 174, 753–756. [Google Scholar] [CrossRef] [PubMed]
- Gatti, L.L.; Burbano, R.R.; Zambaldi-Tunes, M.; de-Labio, R.W.; de Assumpcao, P.P.; de Arruda Cardoso-Smith, M.; Marques-Payao, S.L. Interleukin-6 polymorphisms, Helicobacter pylori infection in adult Brazilian patients with chronic gastritis and gastric adenocarcinoma. Arch. Med. Res. 2007, 38, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Rose-John, S. Interleukin-6 Family Cytokines. Cold Spring Harb. Perspect. Biol. 2018, 10. [Google Scholar] [CrossRef] [PubMed]
- Zimering, J.H.; Williams, M.R.; Eiras, M.E.; Fallon, B.A.; Logigian, E.L.; Dworkin, R.H. Acute and chronic pain associated with Lyme borreliosis: Clinical characteristics and pathophysiologic mechanisms. Pain 2014, 155, 1435–1438. [Google Scholar] [CrossRef] [PubMed]
- e!Ensembl. Available online: https://www.ensembl.org/Homo_sapiens/Variation/Population?db=core;r=7:22726526-22727526;v=rs1800795;vdb=variation;vf=415970384 (accessed on 25 July 2019).
- Landmann, R.; Knopf, H.P.; Link, S.; Sansano, S.; Schumann, R.; Zimmerli, W. Human monocyte CD14 is upregulated by lipopolysaccharide. Infect. Immun. 1996, 64, 1762–1769. [Google Scholar] [Green Version]
- Thomsen, J.H.; Etzerodt, A.; Svendsen, P.; Moestrup, S.K. The haptoglobin-CD163-heme oxygenase-1 pathway for hemoglobin scavenging. Oxid. Med. Cell. Longev. 2013, 2013, 523652. [Google Scholar] [CrossRef]
- Lasky, C.E.; Olson, R.M.; Brown, C.R. Macrophage Polarization during Murine Lyme Borreliosis. Infect. Immun. 2015, 83, 2627–2635. [Google Scholar] [CrossRef] [Green Version]
- Straubinger, R.K.; Straubinger, A.F.; Summers, B.A.; Jacobson, R.H. Status of Borrelia burgdorferi infection after antibiotic treatment and the effects of corticosteroids: An experimental study. J. Infect. Dis. 2000, 181, 1069–1081. [Google Scholar] [CrossRef]
- Khazali, A.S.; Clark, A.M.; Wells, A. Inflammatory cytokine IL-8/CXCL8 promotes tumour escape from hepatocyte-induced dormancy. Br. J. Cancer 2018, 118, 566–576. [Google Scholar] [CrossRef]
- Latz, E.; Xiao, T.S.; Stutz, A. Activation and regulation of the inflammasomes. Nat. Rev. Immunol. 2013, 13, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Oosting, M.; van de Veerdonk, F.L.; Kanneganti, T.D.; Sturm, P.; Verschueren, I.; Berende, A.; van der Meer, J.W.; Kullberg, B.J.; Netea, M.G.; Joosten, L.A. Borrelia species induce inflammasome activation and IL-17 production through a caspase-1-dependent mechanism. Eur. J. Immunol. 2011, 41, 172–181. [Google Scholar] [CrossRef] [PubMed]
- Yrjanainen, H.; Hytonen, J.; Song, X.Y.; Oksi, J.; Hartiala, K.; Viljanen, M.K. Anti-tumor necrosis factor-alpha treatment activates Borrelia burgdorferi spirochetes 4 weeks after ceftriaxone treatment in C3H/He mice. J. Infect. Dis. 2007, 195, 1489–1496. [Google Scholar] [CrossRef] [PubMed]
- McSweegan, E. TNF antibodies get new lease. Nat. Med. 1997, 3, 130. [Google Scholar] [CrossRef]
- Middelveen, M.J.; Sapi, E.; Burke, J.; Filush, K.R.; Franco, A.; Fesler, M.C.; Stricker, R.B. Persistent Borrelia Infection in Patients with Ongoing Symptoms of Lyme Disease. Healthcare 2018, 6, 33. [Google Scholar] [CrossRef]
- Noss, E.H.; Nguyen, H.N.; Chang, S.K.; Watts, G.F.; Brenner, M.B. Genetic polymorphism directs IL-6 expression in fibroblasts but not selected other cell types. Proc. Natl. Acad. Sci. USA 2015, 112, 14948–14953. [Google Scholar] [CrossRef] [Green Version]
- Muller, K.E. Damage of collagen and elastic fibres by borrelia burgdorferi—Known and new clinical and histopathological aspects. Open Neurol. J. 2012, 6, 179–186. [Google Scholar] [CrossRef]
- Ishida, K.; Kobayashi, T.; Ito, S.; Komatsu, Y.; Yokoyama, T.; Okada, M.; Abe, A.; Murasawa, A.; Yoshie, H. Interleukin-6 gene promoter methylation in rheumatoid arthritis and chronic periodontitis. J. Periodontol. 2012, 83, 917–925. [Google Scholar] [CrossRef]
- Nile, C.J.; Read, R.C.; Akil, M.; Duff, G.W.; Wilson, A.G. Methylation status of a single CpG site in the IL6 promoter is related to IL6 messenger RNA levels and rheumatoid arthritis. Arthritis Rheum. 2008, 58, 2686–2693. [Google Scholar] [CrossRef]
- Fok, E.T.; Davignon, L.; Fanucchi, S.; Mhlanga, M.M. The lncRNA Connection Between Cellular Metabolism and Epigenetics in Trained Immunity. Front. Immunol. 2018, 9, 3184. [Google Scholar] [CrossRef] [PubMed]
- Goc, A.; Rath, M. The anti-borreliae efficacy of phytochemicals and micronutrients: An update. Ther. Adv. Infect. Dis. 2016, 3, 75–82. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Group * | Secondary Disease | Patient Number | Inflammatory Phenotype |
|---|---|---|---|
| 1 | Malignancies | 10 | IL-1β↑, IL-6↑, IL-8↑ CD163+↓ TNF-α* |
| 2 | Autoimmunity | 27 | IL-8↑, CD163+↓ TNF-α* |
| 3 | Neurological dysfunctions (including PD, ALS, psychiatric diseases) | 30 | IL-1β↑, IL-8↑ TNF-α* |
| 4 | Multiple infections | 40 | IL-1β↑, IL-8↑, CD163+↓ TNF-α* |
| 1–4 | Total | 107 | IL-8↑, TNF-α * |
| Mediator | Group (Secondary Diseases) | Patient Groups | Median (±Range) | Significance (Groups vs. Control) |
|---|---|---|---|---|
| IL-1β | 1 | Malignancies | 6.03 (±64.8) | ** |
| 2 | Autoimmunity | 2.88 (±21) | n.s. | |
| 3 | Neurological Dysfunctions | 3.28 (±44.4) | ** | |
| 4 | None | 5.69 (±346.4) | ** | |
| Ctrl | Control | 2.48 (±16) | ||
| IL-6 | 1 | Malignancies | 4.11 (±36.5) | * |
| 2 | Autoimmunity | 1.4 (±13.9) | n.s. | |
| 3 | Neurological Dysfunctions | 0.76 (±45.3) | n.s. | |
| 4 | None | 2.08 (±64.7) | n.s. | |
| Ctrl | Control | 1.38 (±30.2) | ||
| IL-8 | 1 | Malignancies | 406 (±5013.2) | *** |
| 2 | Autoimmunity | 37.2 (±2370.8) | ** | |
| 3 | Neurological Dysfunctions | 60 (±12726) | ** | |
| 4 | None | 49 (±6656.4) | ** | |
| Ctrl | Control | 7.45 (±2011.7) | ||
| TNF-α | 1 | Malignancies | 19.7 (±35.4) | n.s. |
| 2 | Autoimmunity | 15.2 (±25.1) | n.s. | |
| 3 | Neurological Dysfunctions | 15.4 (±58.8) | n.s. | |
| 4 | None | 17.7 (±97.3) | n.s. | |
| Ctrl | Control | 14.8 (±171.4) | ||
| CD163+ Macrophages | 1 | Malignancies | 1.71 (±93.6) | * |
| 2 | Autoimmunity | 2.98 (±65) | *** | |
| 3 | Neurological Dysfunctions | 9.7 (±74.4) | n.s. | |
| 4 | None | 5.63 (±68.3) | ** | |
| Ctrl | Control | 29.96 (±84.4) |
| SNP Genotype | Lyme Borreliosis Patients n (%) | E!Ensembl Reference Population n (%) |
|---|---|---|
| IL-6 rs1800795 Genotype | ||
| G/G | 36 (33.6%) | 181 (36%) |
| G/C | 59 (55.1%) | 226 (44.9%) |
| C/C | 12 (11.2%) | 96 (19.1%) |
| Total | 107 (100%) | 503 (100%) |
| Secondary Disease Group | Malignancies n (%) 1 | Autoimmunity n (%) 2 | Neurological Dysfunction n (%) 3 | None * n (%) 4 |
|---|---|---|---|---|
| IL-6 rs1800795 | ||||
| G/G | 0 (0%) | 7 (25.9%) | 13 (43.3%) | 16 (40%) |
| G/C | 7 (70%) | 18 (66.7%) | 13 (43.3%) | 21 (52.5%) |
| C/C | 3 (30%) | 2 (7.4%) | 4 (13.3%) | 3 (7.5%) |
| Total | 10 (100%) | 27 (100%) | 30 (100%) | 40 (100%) |
| p = | 0.0620 | 0.089 | 0.6264 | 0.1864 |
| χ2 = | 5.56 | 5.548 | 0.9355 | 3.359 |
| Mediator | Genotype IL-6 Promotor rs1800795 | Median (±Range) | Significance (Genotype vs. Control) |
|---|---|---|---|
| IL-1β | G/G | 3.27 (±346.4) | * |
| G/C | 3.52 (±65.6) | ** | |
| C/C | 5.81 (±18.7) | ** | |
| Control | 2.48 (±16) | ||
| IL-6 | G/G | 0.3 (±7.6) | n.s. |
| G/C | 2.88 (±64.7) | ** | |
| C/C | 0.12 (±29.3) | n.s. | |
| Control | 1.38 (±30.2) | ||
| IL-8 | G/G | 39.3 (±12723.4) | * |
| G/C | 66 (±5016.3) | **** | |
| C/C | 168 (±1220.8) | ** | |
| Control | 7.45 (±2011.7) | ||
| CD163+ Macrophages | G/G | 3.28 (±72.49) | ** |
| G/C | 3.93 (±68.26) | **** | |
| C/C | 12.66 (±93.64) | n.s. | |
| Control | 29.96 (±84.4) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hein, T.M.; Sander, P.; Giryes, A.; Reinhardt, J.-O.; Hoegel, J.; Schneider, E.M. Cytokine Expression Patterns and Single Nucleotide Polymorphisms (SNPs) in Patients with Chronic Borreliosis. Antibiotics 2019, 8, 107. https://doi.org/10.3390/antibiotics8030107
Hein TM, Sander P, Giryes A, Reinhardt J-O, Hoegel J, Schneider EM. Cytokine Expression Patterns and Single Nucleotide Polymorphisms (SNPs) in Patients with Chronic Borreliosis. Antibiotics. 2019; 8(3):107. https://doi.org/10.3390/antibiotics8030107
Chicago/Turabian StyleHein, Tabea M., Philip Sander, Anwar Giryes, Jan-Olaf Reinhardt, Josef Hoegel, and E. Marion Schneider. 2019. "Cytokine Expression Patterns and Single Nucleotide Polymorphisms (SNPs) in Patients with Chronic Borreliosis" Antibiotics 8, no. 3: 107. https://doi.org/10.3390/antibiotics8030107
APA StyleHein, T. M., Sander, P., Giryes, A., Reinhardt, J. -O., Hoegel, J., & Schneider, E. M. (2019). Cytokine Expression Patterns and Single Nucleotide Polymorphisms (SNPs) in Patients with Chronic Borreliosis. Antibiotics, 8(3), 107. https://doi.org/10.3390/antibiotics8030107