Proteomic and Metabolomic Analyses of a Tea-Tree Oil-Selected Staphylococcus aureus Small Colony Variant



Abstract
:1. Introduction
2. Results
2.1. General Proteomics Comparison
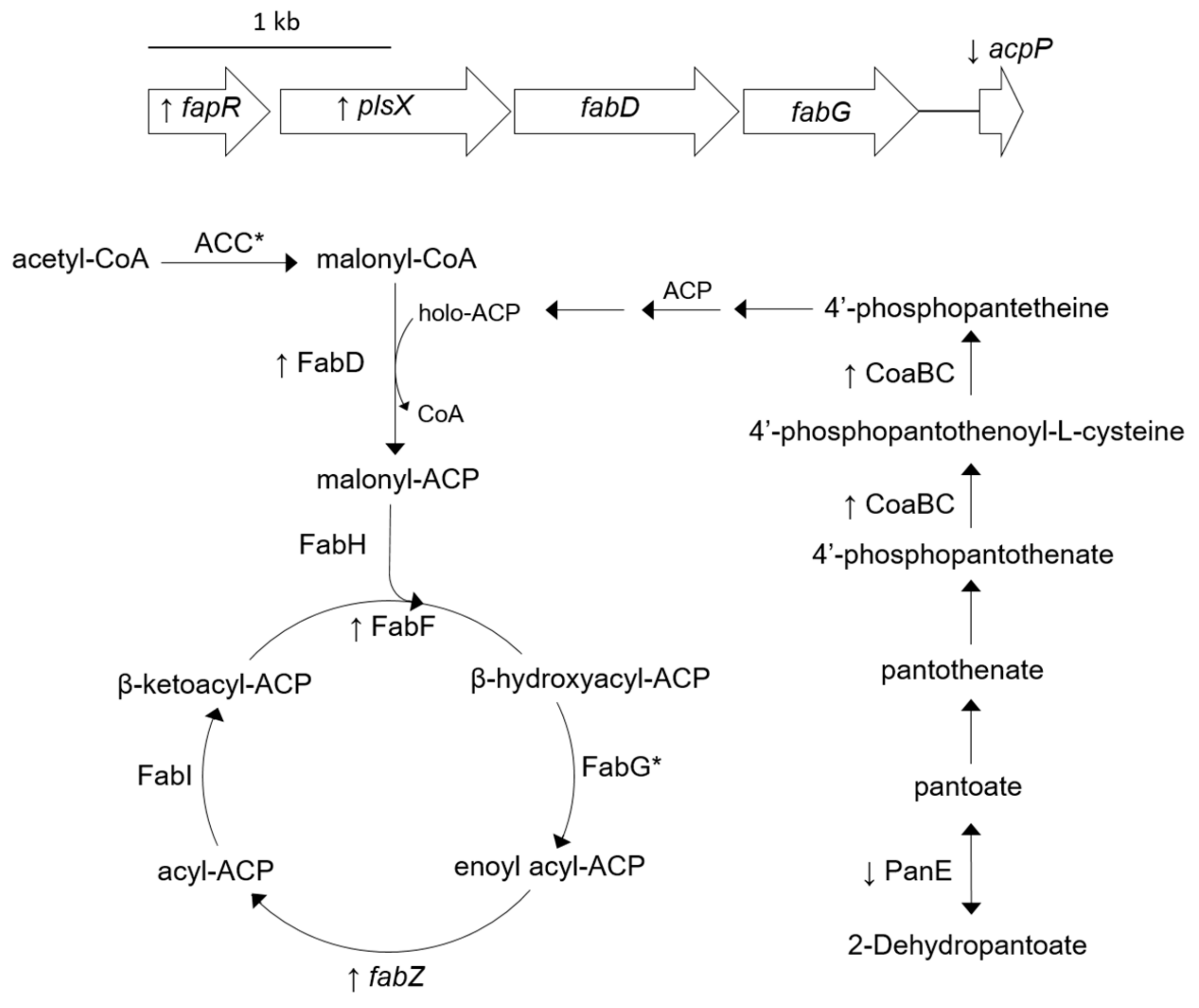
2.2. Alterations in Fatty Acid Biosynthesis in SH1000-TTORS-1
2.3. Proteins Involved with Central Metabolism Are Altered in SH1000-TTORS-1
2.4. Proteins Associated with Stress and Stringent Response Are Altered in SH1000-TTORS-1
2.5. Additional Proteins Altered in SH1000-TTORS-1
2.6. Metabolomics Analysis
3. Discussion
4. Materials and Methods
4.1. Strains, Antibiotic Gradient Plate Analysis, Colony Size, and Chemicals
4.2. Protein Extraction
4.3. Mass Spectrometry Analysis
4.4. Metabolite Extraction and Analysis
4.5. RNA Extraction and RT-PCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- King, M.D.; Humphrey, B.J.; Wang, Y.F.; Kourbatova, E.V.; Ray, S.M.; Blumberg, H.M. Emergence of community-acquired methicillin-resistant Staphylococcus aureus USA 300 clone as the predominant cause of skin and soft-tissue infections. Ann. Intern. Med. 2006, 144, 309–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Proctor, R.A.; van Langevelde, P.; Kristjansson, M.; Maslow, J.N.; Arbeit, R.D. Persistent and relapsing infections associated with small-colony variants of Staphylococcus aureus. Clin. Infect. Dis. 1995, 20, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Proctor, R.A.; Balwit, J.M.; Vesga, O. Variant subpopulations of Staphylococcus aureus as cause of persistent and recurrent infections. Infect. Agents Dis. 1994, 3, 302–312. [Google Scholar] [PubMed]
- Proctor, R.A.; von Eiff, C.; Kahl, B.C.; Becker, K.; McNamara, P.; Herrmann, M.; Peters, G. Small colony variants: A pathogenic form of bacteria that facilitates persistent and recurrent infections. Nat. Rev. Microbiol. 2006, 4, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Kohler, C.; von Eiff, C.; Liebeke, M.; McNamara, P.J.; Lalk, M.; Proctor, R.A.; Hecker, M.; Engelmann, S. A defect in menadione biosynthesis induces global changes in gene expression in Staphylococcus aureus. J. Bacteriol. 2008, 190, 6351–6364. [Google Scholar] [CrossRef] [Green Version]
- Kohler, C.; von Eiff, C.; Peters, G.; Proctor, R.A.; Hecker, M.; Engelmann, S. Physiological characterization of a heme-deficient mutant of Staphylococcus aureus by a proteomic approach. J. Bacteriol. 2003, 185, 6928–6937. [Google Scholar] [CrossRef] [Green Version]
- Seggewiss, J.; Becker, K.; Kotte, O.; Eisenacher, M.; Yazdi, M.R.; Fischer, A.; McNamara, P.; Al Laham, N.; Proctor, R.; Peters, G.; et al. Reporter metabolite analysis of transcriptional profiles of a Staphylococcus aureus strain with normal phenotype and its isogenic hemB mutant displaying the small-colony-variant phenotype. J. Bacteriol. 2006, 188, 7765–7777. [Google Scholar] [CrossRef] [Green Version]
- Kriegeskorte, A.; Konig, S.; Sander, G.; Pirkl, A.; Mahabir, E.; Proctor, R.A.; von Eiff, C.; Peters, G.; Becker, K. Small colony variants of Staphylococcus aureus reveal distinct protein profiles. Proteomics 2011, 11, 2476–2490. [Google Scholar] [CrossRef]
- Kriegeskorte, A.; Grubmuller, S.; Huber, C.; Kahl, B.C.; von Eiff, C.; Proctor, R.A.; Peters, G.; Eisenreich, W.; Becker, K. Staphylococcus aureus small colony variants show common metabolic features in central metabolism irrespective of the underlying auxotrophism. Front. Cell Infect. Microbiol. 2014, 4, 141. [Google Scholar] [CrossRef]
- Von Eiff, C.; Heilmann, C.; Proctor, R.A.; Woltz, C.; Peters, G.; Gotz, F. A site-directed Staphylococcus aureus hemB mutant is a small-colony variant which persists intracellularly. J. Bacteriol. 1997, 179, 4706–4712. [Google Scholar] [CrossRef] [Green Version]
- Schaaff, F.; Bierbaum, G.; Baumert, N.; Bartmann, P.; Sahl, H.G. Mutations are involved in emergence of aminoglycoside-induced small colony variants of Staphylococcus aureus. Int. J. Med. Microbiol. 2003, 293, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Bates, D.M.; von Eiff, C.; McNamara, P.J.; Peters, G.; Yeaman, M.R.; Bayer, A.S.; Proctor, R.A. Staphylococcus aureus menD and hemB mutants are as infective as the parent strains, but the menadione biosynthetic mutant persists within the kidney. J. Infect. Dis. 2003, 187, 1654–1661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, M.O.; Watson, S.P.; Poole, R.K.; Foster, S.J. CtaA of Staphylococcus aureus is required for starvation survival, recovery, and cytochrome biosynthesis. J. Bacteriol. 1999, 181, 501–507. [Google Scholar] [PubMed]
- Chatterjee, I.; Kriegeskorte, A.; Fischer, A.; Deiwick, S.; Theimann, N.; Proctor, R.A.; Peters, G.; Herrmann, M.; Kahl, B.C. In vivo mutations of thymidylate synthase (encoded by thyA) are responsible for thymidine dependency in clinical small-colony variants of Staphylococcus aureus. J. Bacteriol. 2008, 190, 834–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duval, B.D.; Mathew, A.; Satola, S.W.; Shafer, W.M. Altered growth, pigmentation, and antimicrobial susceptibility properties of Staphylococcus aureus due to loss of the major cold shock gene cspB. Antimicrob. Agents Chemother. 2010, 54, 2283–2290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Chua, K.; Davies, J.K.; Newton, H.J.; Seemann, T.; Harrison, P.F.; Holmes, N.E.; Rhee, H.W.; Hong, J.I.; Hartland, E.L.; et al. Two novel point mutations in clinical Staphylococcus aureus reduce linezolid susceptibility and switch on the stringent response to promote persistent infection. PLoS Pathog. 2010, 6, e1000944. [Google Scholar] [CrossRef] [Green Version]
- Oun, S.; Redder, P.; Didier, J.P.; Francois, P.; Corvaglia, A.R.; Buttazzoni, E.; Giraud, C.; Girard, M.; Schrenzel, J.; Linder, P. The CshA DEAD-box RNA helicase is important for quorum sensing control in Staphylococcus aureus. RNA Biol. 2013, 10, 157–165. [Google Scholar] [CrossRef] [Green Version]
- Gaupp, R.; Schlag, S.; Liebeke, M.; Lalk, M.; Gotz, F. Advantage of upregulation of succinate dehydrogenase in Staphylococcus aureus biofilms. J. Bacteriol. 2010, 192, 2385–2394. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Wright, J.A.; Osman, A.A.; Nair, S.P. An aroD Ochre Mutation Results in a Staphylococcus aureus Small Colony Variant That Can Undergo Phenotypic Switching via Two Alternative Mechanisms. Front. Microbiol. 2017, 8, 1001. [Google Scholar] [CrossRef]
- Norstrom, T.; Lannergard, J.; Hughes, D. Genetic and phenotypic identification of fusidic acid-resistant mutants with the small-colony-variant phenotype in Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 4438–4446. [Google Scholar] [CrossRef] [Green Version]
- Painter, K.L.; Strange, E.; Parkhill, J.; Bamford, K.B.; Armstrong-James, D.; Edwards, A.M. Staphylococcus aureus adapts to oxidative stress by producing H2O2-resistant small-colony variants via the SOS response. Infect. Immun. 2015, 83, 1830–1844. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Huseby, D.L.; Brandis, G.; Hughes, D. Alternative Evolutionary Pathways for Drug-Resistant Small Colony Variant Mutants in Staphylococcus aureus. MBio 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (Tea Tree) oil: A review of antimicrobial and other medicinal properties. Clin. Microbiol. Rev. 2006, 19, 50–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carson, C.F.; Cookson, B.D.; Farrelly, H.D.; Riley, T.V. Susceptibility of methicillin-resistant Staphylococcus aureus to the essential oil of Melaleuca alternifolia. J. Antimicrob. Chemother. 1995, 35, 421–424. [Google Scholar] [CrossRef]
- Cox, S.D.; Gustafson, J.E.; Mann, C.M.; Markham, J.L.; Liew, Y.C.; Hartland, R.P.; Bell, H.C.; Warmington, J.R.; Wyllie, S.G. Tea tree oil causes K+ leakage and inhibits respiration in Escherichia coli. Lett. Appl. Microbiol. 1998, 26, 355–358. [Google Scholar] [CrossRef]
- Gustafson, J.E.; Liew, Y.C.; Chew, S.; Markham, J.; Bell, H.C.; Wyllie, S.G.; Warmington, J.R. Effects of tea tree oil on Escherichia coli. Lett. Appl. Microbiol. 1998, 26, 194–198. [Google Scholar] [CrossRef]
- Cox, S.D.; Mann, C.M.; Markham, J.L.; Bell, H.C.; Gustafson, J.E.; Warmington, J.R.; Wyllie, S.G. The mode of antimicrobial action of the essential oil of Melaleuca alternifolia (tea tree oil). J. Appl. Microbiol. 2000, 88, 170–175. [Google Scholar] [CrossRef]
- Carson, C.F.; Mee, B.J.; Riley, T.V. Mechanism of action of Melaleuca alternifolia (tea tree) oil on Staphylococcus aureus determined by time-kill, lysis, leakage, and salt tolerance assays and electron microscopy. Antimicrob. Agents Chemother. 2002, 46, 1914–1920. [Google Scholar] [CrossRef] [Green Version]
- Cuaron, J.A.; Dulal, S.; Song, Y.; Singh, A.K.; Montelongo, C.E.; Yu, W.; Nagarajan, V.; Jayaswal, R.K.; Wilkinson, B.J.; Gustafson, J.E. Tea tree oil-induced transcriptional alterations in Staphylococcus aureus. Phytother. Res. PTR 2013, 27, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Cuaron, J.A.; Dulal, S.; Cooke, P.H.; Torres, N.J.; Gustafson, J.E. The isolation of Staphylococcus aureus tea tree oil-reduced susceptibility mutants. Phytother. Res. PTR 2014, 28, 1240–1245. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, M.; Cronan, J.E., Jr. The gene encoding Escherichia coli acyl carrier protein lies within a cluster of fatty acid biosynthetic genes. J. Biol. Chem. 1992, 267, 5751–5754. [Google Scholar] [PubMed]
- Majerus, P.W.; Alberts, A.W.; Vagelos, P.R. The Acyl Carrier Protein of Fatty Acid Synthesis: Purification, Physical Properties, and Substrate Binding Site. Proc. Natl. Acad. Sci. USA 1964, 51, 1231–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, D.I.; Vogel, H.J. Current understanding of fatty acid biosynthesis and the acyl carrier protein. Biochem. J. 2010, 430, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rock, C.O.; Cronan, J.E. Escherichia coli as a model for the regulation of dissociable (type II) fatty acid biosynthesis. Biochim. Biophys. Acta 1996, 1302, 1–16. [Google Scholar] [CrossRef]
- Paoletti, L.; Lu, Y.J.; Schujman, G.E.; de Mendoza, D.; Rock, C.O. Coupling of fatty acid and phospholipid synthesis in Bacillus subtilis. J. Bacteriol. 2007, 189, 5816–5824. [Google Scholar] [CrossRef] [Green Version]
- Schujman, G.E.; Guerin, M.; Buschiazzo, A.; Schaeffer, F.; Llarrull, L.I.; Reh, G.; Vila, A.J.; Alzari, P.M.; de Mendoza, D. Structural basis of lipid biosynthesis regulation in Gram-positive bacteria. EMBO J. 2006, 25, 4074–4083. [Google Scholar] [CrossRef]
- Schujman, G.E.; Paoletti, L.; Grossman, A.D.; de Mendoza, D. FapR, a bacterial transcription factor involved in global regulation of membrane lipid biosynthesis. Dev. Cell 2003, 4, 663–672. [Google Scholar] [CrossRef] [Green Version]
- Lannergard, J.; von Eiff, C.; Sander, G.; Cordes, T.; Seggewiss, J.; Peters, G.; Proctor, R.A.; Becker, K.; Hughes, D. Identification of the genetic basis for clinical menadione-auxotrophic small-colony variant isolates of Staphylococcus aureus. Antimicrob. Agents Chemother. 2008, 52, 4017–4022. [Google Scholar] [CrossRef] [Green Version]
- Strauss, E.; Kinsland, C.; Ge, Y.; McLafferty, F.W.; Begley, T.P. Phosphopantothenoylcysteine synthetase from Escherichia coli. Identification and characterization of the last unidentified coenzyme A biosynthetic enzyme in bacteria. J. Biol. Chem. 2001, 276, 13513–13516. [Google Scholar] [CrossRef] [Green Version]
- Kupke, T.; Uebele, M.; Schmid, D.; Jung, G.; Blaesse, M.; Steinbacher, S. Molecular characterization of lantibiotic-synthesizing enzyme EpiD reveals a function for bacterial Dfp proteins in coenzyme A biosynthesis. J. Biol. Chem. 2000, 275, 31838–31846. [Google Scholar] [CrossRef] [Green Version]
- Zheng, R.; Blanchard, J.S. Kinetic and mechanistic analysis of the E. coli panE-encoded ketopantoate reductase. Biochemistry 2000, 39, 3708–3717. [Google Scholar] [CrossRef]
- Bazaid, A.S.; Forbes, S.; Humphreys, G.J.; Ledder, R.G.; O’Cualain, R.; McBain, A.J. Fatty Acid Supplementation Reverses the Small Colony Variant Phenotype in Triclosan-Adapted Staphylococcus aureus: Genetic, Proteomic and Phenotypic Analyses. Sci. Rep. 2018, 8, 3876. [Google Scholar] [CrossRef]
- Nie, L.; Ren, Y.; Schulz, H. Identification and characterization of Escherichia coli thioesterase III that functions in fatty acid beta-oxidation. Biochemistry 2008, 47, 7744–7751. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.B.; Frank, M.W.; Subramanian, C.; Saenkham, P.; Rock, C.O. Metabolic basis for the differential susceptibility of Gram-positive pathogens to fatty acid synthesis inhibitors. Proc. Natl. Acad. Sci. USA 2011, 108, 15378–15383. [Google Scholar] [CrossRef] [Green Version]
- Heath, R.J.; Rock, C.O. Enoyl-acyl carrier protein reductase (fabI) plays a determinant role in completing cycles of fatty acid elongation in Escherichia coli. J. Biol. Chem. 1995, 270, 26538–26542. [Google Scholar] [CrossRef] [Green Version]
- Sadykov, M.R.; Thomas, V.C.; Marshall, D.D.; Wenstrom, C.J.; Moormeier, D.E.; Widhelm, T.J.; Nuxoll, A.S.; Powers, R.; Bayles, K.W. Inactivation of the Pta-AckA pathway causes cell death in Staphylococcus aureus. J. Bacteriol. 2013, 195, 3035–3044. [Google Scholar] [CrossRef] [Green Version]
- Tuchscherr, L.; Bischoff, M.; Lattar, S.M.; Noto Llana, M.; Pfortner, H.; Niemann, S.; Geraci, J.; Van de Vyver, H.; Fraunholz, M.J.; Cheung, A.L.; et al. Sigma Factor SigB Is Crucial to Mediate Staphylococcus aureus Adaptation during Chronic Infections. PLoS Pathog. 2015, 11, e1004870. [Google Scholar] [CrossRef]
- Tuchscherr, L.; Geraci, J.; Loffler, B. Staphylococcus aureus Regulator Sigma B is Important to Develop Chronic Infections in Hematogenous Murine Osteomyelitis Model. Pathogens 2017, 6, 31. [Google Scholar] [CrossRef] [Green Version]
- Vijay, K.; Brody, M.S.; Fredlund, E.; Price, C.W. A PP2C phosphatase containing a PAS domain is required to convey signals of energy stress to the sigmaB transcription factor of Bacillus subtilis. Mol. Microbiol. 2000, 35, 180–188. [Google Scholar] [CrossRef]
- Bischoff, M.; Dunman, P.; Kormanec, J.; Macapagal, D.; Murphy, E.; Mounts, W.; Berger-Bachi, B.; Projan, S. Microarray-based analysis of the Staphylococcus aureus sigmaB regulon. J. Bacteriol. 2004, 186, 4085–4099. [Google Scholar] [CrossRef] [Green Version]
- Dalebroux, Z.D.; Svensson, S.L.; Gaynor, E.C.; Swanson, M.S. ppGpp conjures bacterial virulence. Microbiol. Mol. Biol. Rev. 2010, 74, 171–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, T.; Kastle, B.; Gratani, F.L.; Goerke, C.; Wolz, C. Two small (p)ppGpp synthases in Staphylococcus aureus mediate tolerance against cell envelope stress conditions. J. Bacteriol. 2014, 196, 894–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiss, S.; Pane-Farre, J.; Fuchs, S.; Francois, P.; Liebeke, M.; Schrenzel, J.; Lindequist, U.; Lalk, M.; Wolz, C.; Hecker, M.; et al. Global analysis of the Staphylococcus aureus response to mupirocin. Antimicrob. Agents Chemother. 2012, 56, 787–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meinnel, T.; Mechulam, Y.; Blanquet, S. Methionine as translation start signal: A review of the enzymes of the pathway in Escherichia coli. Biochimie 1993, 75, 1061–1075. [Google Scholar] [CrossRef]
- Blake, K.L.; O’Neill, A.J.; Mengin-Lecreulx, D.; Henderson, P.J.; Bostock, J.M.; Dunsmore, C.J.; Simmons, K.J.; Fishwick, C.W.; Leeds, J.A.; Chopra, I. The nature of Staphylococcus aureus MurA and MurZ and approaches for detection of peptidoglycan biosynthesis inhibitors. Mol. Microbiol. 2009, 72, 335–343. [Google Scholar] [CrossRef]
- Halsey, C.R.; Lei, S.; Wax, J.K.; Lehman, M.K.; Nuxoll, A.S.; Steinke, L.; Sadykov, M.; Powers, R.; Fey, P.D. Amino Acid Catabolism in Staphylococcus aureus and the Function of Carbon Catabolite Repression. mBio 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Townsend, D.E.; Kaenjak, A.; Jayaswal, R.K.; Wilkinson, B.J. Proline is biosynthesized from arginine in Staphylococcus aureus. Microbiology 1996, 142 Pt 6, 1491–1497. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, K.J.; Bjorge, D.; Schwan, W.R. Mutational and transcriptional analyses of the Staphylococcus aureus low-affinity proline transporter OpuD during in vitro growth and infection of murine tissues. FEMS Immunol. Med. Microbiol. 2011, 61, 346–355. [Google Scholar] [CrossRef]
- Stockland, A.E.; San Clemente, C.L. Lactate dehydrogenase activity in certain strains of Staphylococcus aureus. J. Bacteriol. 1968, 95, 74–80. [Google Scholar]
- Fuller, J.R.; Vitko, N.P.; Perkowski, E.F.; Scott, E.; Khatri, D.; Spontak, J.S.; Thurlow, L.R.; Richardson, A.R. Identification of a lactate-quinone oxidoreductase in Staphylococcus aureus that is essential for virulence. Front. Cell Infect. Microbiol. 2011, 1, 19. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.J.; Chang, J.; Singh, M. Peptidoglycan architecture of Gram-positive bacteria by solid-state NMR. Biochim. Biophys. Acta 2015, 1848, 350–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stulke, J.; Hillen, W. Carbon catabolite repression in bacteria. Curr. Opin. Microbiol. 1999, 2, 195–201. [Google Scholar] [CrossRef]
- Horsburgh, M.J.; Aish, J.L.; White, I.J.; Shaw, L.; Lithgow, J.K.; Foster, S.J. sigmaB modulates virulence determinant expression and stress resistance: Characterization of a functional rsbU strain derived from Staphylococcus aureus 8325-4. J. Bacteriol. 2002, 184, 5457–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Leary, J.O.; Langevin, M.J.; Price, C.T.; Blevins, J.S.; Smeltzer, M.S.; Gustafson, J.E. Effects of sarA inactivation on the intrinsic multidrug resistance mechanism of Staphylococcus aureus. FEMS Microbiol. Lett. 2004, 237, 297–302. [Google Scholar] [CrossRef]
- Wolff, S.; Hahne, H.; Hecker, M.; Becher, D. Complementary analysis of the vegetative membrane proteome of the human pathogen Staphylococcus aureus. Mol. Cell Proteom. 2008, 7, 1460–1468. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Voruganti, S.; Lacroix, J.C.; Rogers, C.N.; Rogers, J.; Matts, R.L.; Hartson, S.D. The anticancer drug AUY922 generates a proteomics fingerprint that is highly conserved among structurally diverse Hsp90 inhibitors. J. Proteome Res. 2013, 12, 3697–3706. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Neuhauser, N.; Michalski, A.; Scheltema, R.A.; Olsen, J.V.; Mann, M. Andromeda: A peptide search engine integrated into the MaxQuant environment. J. Proteome Res. 2011, 10, 1794–1805. [Google Scholar] [CrossRef]
- Zhang, W.; Zhang, J.; Xu, C.; Li, N.; Liu, H.; Ma, J.; Zhu, Y.; Xie, H. LFQuant: A label-free fast quantitative analysis tool for high-resolution LC-MS/MS proteomics data. Proteomics 2012, 12, 3475–3484. [Google Scholar] [CrossRef]
- Tyanova, S.; Cox, J. Perseus: A Bioinformatics Platform for Integrative Analysis of Proteomics Data in Cancer Research. Methods Mol. Biol. 2018, 1711, 133–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Hattangady, D.S.; Singh, A.K.; Muthaiyan, A.; Jayaswal, R.K.; Gustafson, J.E.; Ulanov, A.V.; Li, Z.; Wilkinson, B.J.; Pfeltz, R.F. Genomic, Transcriptomic and Metabolomic Studies of Two Well-Characterized, Laboratory-Derived Vancomycin-Intermediate Staphylococcus aureus Strains Derived from the Same Parent Strain. Antibiotics 2015, 4, 76–112. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| a Locus ID | Protein Symbol | Description | Functional Category | Log2-Fold Change |
|---|---|---|---|---|
| Increased in SH1000-TTORS-1 compared to SH1000 | ||||
| SAOUHSC_00356 | YkoI | membrane protein | uncharacterized | 2.1 |
| SAOUHSC_00452 | YaaQ | cyclic-di-AMP receptor | uncharacterized | 1.4 |
| SAOUHSC_00468 | reactive intermediate/imine deaminase | uncharacterized | 1.5 | |
| SAOUHSC_00604 | WrbA | multimeric flavodoxin | uncharacterized | 2.5 |
| SAOUHSC_00833 | nitroreductase | uncharacterized | 1.9 | |
| SAOUHSC_00850 | SUF system FeS assembly protein | uncharacterized | 1.6 | |
| SAOUHSC_00894 | RocD | ornithine-oxo-acid transaminase | amino acid metabolism | 1.8 |
| SAOUHSC_00921 | FabF | 3-Oxoacyl-synthase | lipid metabolism | 1.8 |
| SAOUHSC_00942 | RelQ | GTP pyrophosphokinase | nucleotide metabolism | 1.4 |
| SAOUHSC_01178 | CoaBC | bifunctional phosphopantothenoylcysteine decarboxylase/phosphopantothenate-cysteine ligase | cofactors and secondary metabolites | 1.3 |
| SAOUHSC_01186 | PrpC | serine/threonine protein phosphatase | uncharacterized | 1.1 |
| SAOUHSC_01198 | FabD | malonyl CoA-acyl carrier protein transacylase | lipid metabolism | 3.0 |
| SAOUHSC_01218 | SucD | succinyl-CoA synthetase subunit alpha | carbohydrate metabolism | 4.0 |
| SAOUHSC_01261 | CinA | competence-damage inducible protein | stress response | 2.6 |
| SAOUHSC_01264 | 71 aa protein | uncharacterized | 18.4 | |
| SAOUHSC_01346 | OpuD | proline/choline/glycine betaine transporter | transporter | 3.0 |
| SAOUHSC_01348 | YneP | acyl-CoA thioesterase | uncharacterized | 6.1 |
| SAOUHSC_01401 | LysA | diaminopimelate decarboxylase | amino acid metabolism | 1.2 |
| SAOUHSC_01490 | Hup | DNA-binding protein HU | DNA replication, recombination and repair | 1.3 |
| SAOUHSC_01594 | predicted oxidoreductase | uncharacterized | 1.9 | |
| SAOUHSC_01768 | Tag | 3-methyladenine DNA glycosylase | DNA replication, recombination and repair | 1.7 |
| SAOUHSC_01814 | universal stress protein | uncharacterized | 1.4 | |
| SAOUHSC_02029 | phi ETA orf 56-like protein | uncharacterized | 1.3 | |
| SAOUHSC_02150 | thioredoxin | uncharacterized | 2.5 | |
| SAOUHSC_02298 | SigB | RNA polymerase sigma factor | stress response | 1.3 |
| SAOUHSC_02301 | RsbU | SigmaB regulation protein | stress response | 1.7 |
| SAOUHSC_02365 | MurZ | UDP-N-acetylglucosamine 1-carboxyvinyltransferase | cell wall metabolism | 1.2 |
| SAOUHSC_02370 | acetyltranferase GNAT family | uncharacterized | 2.1 | |
| SAOUHSC_02390 | lytic regulatory protein | uncharacterized | 1.7 | |
| SAOUHSC_02447 | CurA | NADPH-dependent curcumin reductase | uncharacterized | 5.7 |
| SAOUHSC_02604 | oxidoreductase | uncharacterized | 9.2 | |
| SAOUHSC_02626 | 139 aa protein | uncharacterized | 1.3 | |
| SAOUHSC_02665 | general stress protein 26 | uncharacterized | 2.1 | |
| SAOUHSC_02689 | YndB | conserved protein | uncharacterized | 1.8 |
| SAOUHSC_02737 | epimerase/dehydratase | uncharacterized | 1.7 | |
| SAOUHSC_02774 | alkylhydroperoxidase family enzyme | uncharacterized | 3.5 | |
| SAOUHSC_02827 | GNAT superfamily predicted acetyltransferase | uncharacterized | 2.6 | |
| SAOUHSC_02849 | CidC | pyruvate oxidase | carbohydrate metabolism | 1.8 |
| SAOUHSC_02980 | cysteine hydrolase | uncharacterized | 1.4 | |
| Decreased in SH1000-TTORS-1 compared to SH1000 | ||||
| SAOUHSC_00002 | DnaN | DNA polymerase III subunit beta | DNA replication, recombination and repair | −1.4 |
| SAOUHSC_00019 | PurA | adenylosuccinate synthetase | nucleotide metabolism | −1.4 |
| SAOUHSC_00074 | periplasmic binding protein | transporter | −2.6 | |
| SAOUHSC_00200 | PrsW | membrane proteinase | uncharacterized | −1.7 |
| SAOUHSC_00204 | Hmp | globin domain-containing protein | uncharacterized | −1.4 |
| SAOUHSC_00284 | Hel | 5′-Nucleotidase | nucleotide metabolism | −3.4 |
| SAOUHSC_00362 | 208 aa protein | uncharacterized | −1.9 | |
| SAOUHSC_00369 | prolipoprotein diacylglyceryl transferase | uncharacterized | −1.7 | |
| SAOUHSC_00417 | esterase/lipase | uncharacterized | −2.2 | |
| SAOUHSC_00442 | DnaX | DNA polymerase III subunits gamma and tau | DNA replication, recombination and repair | −1.6 |
| SAOUHSC_00461 | MetG | methionyl-tRNA synthetase | protein synthesis | −1.3 |
| SAOUHSC_00474 | RplY | 50S ribosomal protein L25/general stress protein Ctc | protein synthesis | −1.5 |
| SAOUHSC_00483 | 133 aa protein | uncharacterized | −1.5 | |
| SAOUHSC_00513 | 23S rRNA (guanosine(2251)-2′-O)-methyltransferase RlmB | protein synthesis | −1.6 | |
| SAOUHSC_00542 | phosphatase | uncharacterized | −1.5 | |
| SAOUHSC_00578 | MvaD | mevalonate diphosphate decarboxylase | cofactors and secondary metabolites | −1.2 |
| SAOUHSC_00634 | MntC | ABC transporter substrate binding protein | transporter | −2.5 |
| SAOUHSC_00637 | MntA | uncharacterized ABC-type Mn2+/Zn2+ transport system, ATPase component | transporter | −2.3 |
| SAOUHSC_00669 | YkaA | conserved protein | uncharacterized | −1.5 |
| SAOUHSC_00717 | electron transfer DM13 | uncharacterized | −3.0 | |
| SAOUHSC_00785 | TrxB | thioredoxin reductase | amino acid metabolism | −1.8 |
| SAOUHSC_00795 | GapA1 | glyceraldehyde-3-phosphate dehydrogenase | carbohydrate metabolism | −1.6 |
| SAOUHSC_00798 | GpmI | 2,3-Bisphosphogylcerate-independent phosphoglycerate mutase | carbohydrate metabolism | −1.6 |
| SAOUHSC_00819 | CspC | cold shock protein | stress response | −4.6 |
| SAOUHSC_00838 | YwqG | 292 aa protein | uncharacterized | −1.9 |
| SAOUHSC_00933 | TrpS | tryptophanyl-tRNA synthetase | protein synthesis | −1.2 |
| SAOUHSC_00985 | MenB | 1,4-dihydroxy-2-naphthoyl-CoA synthase | cofactors and secondary metabolites | −1.4 |
| SAOUHSC_01040 | PdhA | pyruvate dehydrogenase complex, E1 subunit alpha | carbohydrate metabolism | −1.5 |
| SAOUHSC_01041 | PdhB | pyruvate dehydrogenase complex, E1 subunit beta | carbohydrate metabolism | −1.5 |
| SAOUHSC_01043 | PdhD | dihydrolipoamide dehydrogenase | carbohydrate metabolism | −1.3 |
| SAOUHSC_01058 | TypA | GTP-binding protein | unclassified | −1.7 |
| SAOUHSC_01064 | PycA | pyruvate carboxylase | carbohydrate metabolism | −1.7 |
| SAOUHSC_01091 | SpoU | tRNA G18 (ribose-2′-O)-methylase | uncharacterized | −1.4 |
| SAOUHSC_01093 | PheT | phenylalanyl-tRNA synthetase subunit beta | protein synthesis | −1.4 |
| SAOUHSC_01110 | Ecb | extracellular complement-binding protein | virulence factor | −4.3 |
| SAOUHSC_01163 | RluA family pseudouridine synthase | uncharacterized | −1.7 | |
| SAOUHSC_01488 | heptaprenyl diphosphate synthase subunit 1 | uncharacterized | −1.3 | |
| SAOUHSC_01525 | phage tail tape measure protein | uncharacterized | −2.2 | |
| SAOUHSC_01587 | RluB | ribosomal large subunit pseudouridine synthase B | protein synthesis | −1.3 |
| SAOUHSC_01661 | tRNA A22 N-methylase | uncharacterized | −1.6 | |
| SAOUHSC_01679 | MiaB | tRNA A37 methylthiotransferase | uncharacterized | −1.8 |
| SAOUHSC_01698 | YbhY | RNA-binding protein | uncharacterized | −1.3 |
| SAOUHSC_01716 | PrtC family collagenase-like protease | uncharacterized | −2.7 | |
| SAOUHSC_01735 | tRNA A37 threonylcarbamoyladenosine dehydratase | uncharacterized | −1.6 | |
| SAOUHSC_01810 | MaeB | malate dehydrogenase | carbohydrate metabolism | −2.0 |
| SAOUHSC_01858 | phenylalanyl-tRNA synthetase subunit beta | uncharacterized | −1.5 | |
| SAOUHSC_01901 | Tal | transaldolase | carbohydrate metabolism | −1.4 |
| SAOUHSC_01979 | DNA-binding transcriptional regulator | uncharacterized | −2.0 | |
| SAOUHSC_02096 | 91 aa protein | uncharacterized | −1.6 | |
| SAOUHSC_02102 | Map | methionine aminopeptidase | protein fate | −1.8 |
| SAOUHSC_02133 | PncB | nicotinate phosphoribosyltransferase | cofactors and secondary metabolites | −1.3 |
| SAOUHSC_02139 | pyrazinamidase/nicotinamidase | cofactors and secondary metabolites | −1.5 | |
| SAOUHSC_02258 | fatty acid repression mutant protein/oxidoreductase | uncharacterized | −2.1 | |
| SAOUHSC_02268 | ScrB | sucrose-6-phosphate hydrolase | carbohydrate metabolism | −1.3 |
| SAOUHSC_02343 | AtpG | F0F1 ATP synthase subunit gamma | energy metabolism | −1.3 |
| SAOUHSC_02360 | Tdk | thymidine kinase | nucleotide metabolism | −2.0 |
| SAOUHSC_02525 | AcrB | multidrug efflux pump subunit | uncharacterized | −1.4 |
| SAOUHSC_02544 | MoaB | molybdopterin precursor biosynthesis | cofactors and secondary metabolites | −1.6 |
| SAOUHSC_02549 | ModA | molybdenum ABC transporter substrate-binding protein | transporter | −1.7 |
| SAOUHSC_02574 | NAD/NADP octopine/nopaline Dehydrogenase, alpha-helical domain | uncharacterized | −1.6 | |
| SAOUHSC_02627 | Acetyl esterase/lipase | Uncharacterized | −1.9 | |
| SAOUHSC_02652 | YhdH/YhfP family putative quinone oxidoreductase | Uncharacterized | −1.3 | |
| SAOUHSC_02697 | YecC | amino acid ABC transporter ATP-binding protein | transporter | −1.4 |
| SAOUHSC_02699 | FliY | substrate binding domain of ABC transporters involved in cystine import | transporter | −1.5 |
| SAOUHSC_02739 | PanE | 2-dehydropantoate 2-reductase | cofactors and secondary metabolites | −1.9 |
| SAOUHSC_02767 | NikA | peptide ABC transporter substrate binding protein | transporter | −2.2 |
| SAOUHSC_02791 | pyrophosphohydrolase | DNA replication, recombination and repair | −1.5 | |
| SAOUHSC_02820 | ATP-binding cassette domain of the bacitracin-resistance transporter | uncharacterized | −14.1 | |
| SAOUHSC_02834 | SrtA | sortase | protein fate | −1.7 |
| SAOUHSC_02911 | adenine nucleotide alpha hydrolase superfamily predicted ATPase | uncharacterized | −4.5 | |
| SAOUHSC_02927 | Lqo | lacatate:quinone oxidoreductase | carbohydrate metabolism | −1.7 |
| SAOUHSC_02976 | ManA | mannose-6-phosphate isomerase | carbohydrate metabolism | −2.0 |
| SAOUHSC_03045 | CspB | cold shock protein | stress response | −4.8 |
| Proteins only detected in SH1000-TTORS-1 | ||||
| SAOUHSC_00077 | diderophore synthetase component | uncharacterized | ||
| SAOUHSC_00128 | Cap5O | cap5O protein/UDP-N-acetyl-D- mannosaminuronic acid dehydrogenase | virulence factor | |
| SAOUHSC_00668 | VraG | ABC transporter permease | transporter | |
| SAOUHSC_01279 | hydrolase alpha/beta fold domain-containing protein | uncharacterized | ||
| SAOUHSC_01258 | protein of unknown function | uncharacterized | ||
| SAOUHSC_02460 | aldo/keto reductase | uncharacterized | ||
| SAOUHSC_02866 | YdfJ | membrane protein | uncharacterized | |
| SAOUHSC_02908 | fructosamine-3-kinase | uncharacterized | ||
| SAOUHSC_02935 | GbsR | DNA binding transcriptional regulator | stress response | |
| Proteins only detected in SH1000 | ||||
| SAOUHSC_00197 | glutaryl-CoA dehydrogenase | lipid metabolism | ||
| SAOUHSC_01128 | ArgF | ornithine carbamoyltransferase | amino acid metabolism | |
| SAOUHSC_02648 | L-lactate permease | transporter | ||
| Locus ID | Gene Symbol | Function | Fold-Change in Gene Expression in SH1000-TTORS-1 |
|---|---|---|---|
| SAOUHSC_00574 | pta | Phosphate acetyltransferase | 2.7 |
| SAOUHSC_01196 | fapR | Fatty acid biosynthesis transcriptional regulator | 6.3 |
| SAOUHSC_01197 | plsX | Glycerol-3-phosphate acyltransferase | 2.3 |
| SAOUHSC_01201 | acpP | Acyl carrier protein | −2.0 |
| SAOUHSC_01820 | ackA | Acetate kinase | 3.2 |
| SAOUHSC_02336 | fabZ | 3-Hydroxyacyl-(acyl carrier protein) dehydratase | 9.1 |
| Metabolite Relative Concentration Per Gram Dry Weight (Mean ± SE) | ||||
|---|---|---|---|---|
| Metabolite Class | Metabolite | SH1000 | SH1000-TTORS-1 | Fold Increase SH1000-TTORS-1/SH1000 |
| Amino acids | 3-Hydroxyproline | *N.D. | 18.9 ± 1.0 | |
| Asparagine | 2698.8 ± 211.5 | 38,678.1 ± 4325.2 | 14.3 | |
| Aspartate | 120,734.5 ± 3382.8 | 193,201.1 ± 19,069.7 | 1.6 | |
| Homoserine | 12.1 ± 3.6 | 40.9 ± 4.3 | 3.3 | |
| Lysine | 531.8 ± 236.9 | 1928.6 ± 79.5 | 3.6 | |
| Methyl-lysine | 1163.5 ± 140.8 | 1830 ± 94.7 | 1.6 | |
| N-acetyltryptophan | 343.9 ± 18.5 | 945.1 ± 157.5 | 2.7 | |
| ornithine | 46.1 ± 3.5 | 1058.9 ± 269.4 | 23.0 | |
| Phenylalanine | 3017.4 ± 501.1 | 6954.5 ± 1149.9 | 2.3 | |
| Proline | 7579.6 ± 515.8 | 20,362 ± 2232.6 | 2.7 | |
| Tyrosine | 578 ± 39.3 | 1073.2 ± 78.1 | 1.9 | |
| Polar organic acids | 2-Hydroxyglutaric acid | 35.2 ± 7.6 | 75.4 ± 9.3 | 2.1 |
| 3-Hydroxypropanoic acid | N.D. | 78.05 ± 15.7 | ||
| Fumarate | 44.1 ± 4.2 | 63.8 ± 0.8 | 1.4 | |
| Malonic acid | N.D. | 7.8 ± 2.3 | ||
| Succinate | 431.7 ± 43.7 | 735.6 ± 72.9 | 1.7 | |
| Sugars | N-acetylglucosamine | N.D. | 124.1 ± 15.5 | |
| Metabolite Relative Concentration Per Gram Dry Weight (Mean ± SE) | ||||
|---|---|---|---|---|
| Metabolite Class | Metabolite | SH1000 | SH1000-TTORS-1 | Fold Decrease SH1000/SH1000-TTORS-1 |
| Amines and polyamines | Adenosine | 2358 ± 110.1 | 1251.9 ± 102 | −1.9 |
| Guanosine | 1051.8 ± 127.9 | *N.D. | ||
| Lactamide | 46.2 ± 5.3 | 6.8 ± 2.3 | −6.8 | |
| Amino acids | Cysteine | 130.4 ± 17.1 | 61.2 ± 3.1 | −2.1 |
| Glycine | 8735.5 ± 164.5 | 5519.1 ± 613.2 | −1.6 | |
| N-acetyl-L-serine | 3464.6 ± 323.1 | 482.4 ± 49.4 | −7.2 | |
| Polar organic acids | 2,4-Dihydroxybutaonic acid | 116.0 ± 3.2 | 37.1 ± 3.4 | −3.1 |
| 2-Methyl-2,3-dihydroxypropanoic acid | 1588.6 ± 90.0 | 389.4 ± 50.1 | −4.1 | |
| Aminomalonic acid | 240.0 ± 5.7 | 118.4 ± 18.2 | −2.0 | |
| Gluconic acid | 195.4 ± 31.7 | 27.3 ± 3.7 | −7.2 | |
| Glucuronic acid-6-phosphate | 191.7 ± 73.3 | N.D. | ||
| Glycolic acid | 1430.8 ± 61.4 | 374.2 ± 52.7 | −3.8 | |
| Lactic acid | 15,550.8 ± 896.5 | 9663.5 ± 1296.1 | −1.6 | |
| N-acetylneuraminic acid | 1295.1 ± 113.2 | 307.9 ± 95.8 | −4.2 | |
| Oxamic acid | 93.7 ± 19.6 | 9 ± 7.1 | −10.4 | |
| Sugars | Fructose-1,6-bisphosphate | 657.3 ± 61.7 | 262.4 ± 53.8 | −2.5 |
| Fructose-6-phosphate | 1422.4 ± 271.7 | 523.5 ± 44.6 | −2.7 | |
| Glucose-6-phosphate | 3539.3 ± 770.5 | 485.5 ± 63.2 | −7.3 | |
| Glyceraldehyde | 3.3 ± 1.1 | N.D. | ||
| Mannitol | 17,749.6 ± 749.9 | 8271.7 ± 941.2 | −2.1 | |
| Mannitol-6-phosphate | 36,039.4 ± 347.6 | 16,171 ± 3376.65 | −2.2 | |
| Mannose | 831.2 ± 70.4 | 74.7 ± 14.2 | −11.1 | |
| Sedoheptulose | 138.9 ± 11.8 | 67.8 ± 14.2 | −2.0 | |
| Sedoheptulose-7-phosphate | 722.7 ± 122 | 186 ± 25.3 | −3.9 | |
| Trehalose | 3243.3 ± 892.6 | 127.6 ± 13.2 | −25.4 | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torres, N.J.; Hartson, S.D.; Rogers, J.; Gustafson, J.E. Proteomic and Metabolomic Analyses of a Tea-Tree Oil-Selected Staphylococcus aureus Small Colony Variant. Antibiotics 2019, 8, 248. https://doi.org/10.3390/antibiotics8040248
Torres NJ, Hartson SD, Rogers J, Gustafson JE. Proteomic and Metabolomic Analyses of a Tea-Tree Oil-Selected Staphylococcus aureus Small Colony Variant. Antibiotics. 2019; 8(4):248. https://doi.org/10.3390/antibiotics8040248
Chicago/Turabian StyleTorres, Nathanial J., Steven D. Hartson, Janet Rogers, and John E. Gustafson. 2019. "Proteomic and Metabolomic Analyses of a Tea-Tree Oil-Selected Staphylococcus aureus Small Colony Variant" Antibiotics 8, no. 4: 248. https://doi.org/10.3390/antibiotics8040248
APA StyleTorres, N. J., Hartson, S. D., Rogers, J., & Gustafson, J. E. (2019). Proteomic and Metabolomic Analyses of a Tea-Tree Oil-Selected Staphylococcus aureus Small Colony Variant. Antibiotics, 8(4), 248. https://doi.org/10.3390/antibiotics8040248