Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin

 ,
,  ,
,  , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
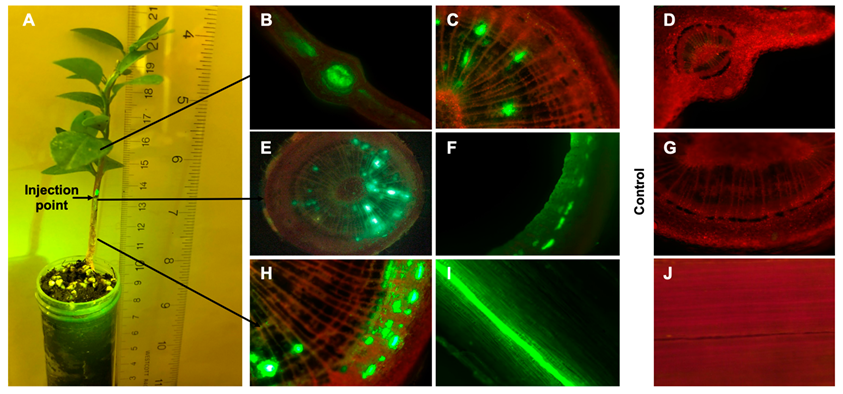
2.1. Translocation of FL-Penicillin in Intact Plants
2.2. Translocation of FL-Penicillin in Girdled Plants
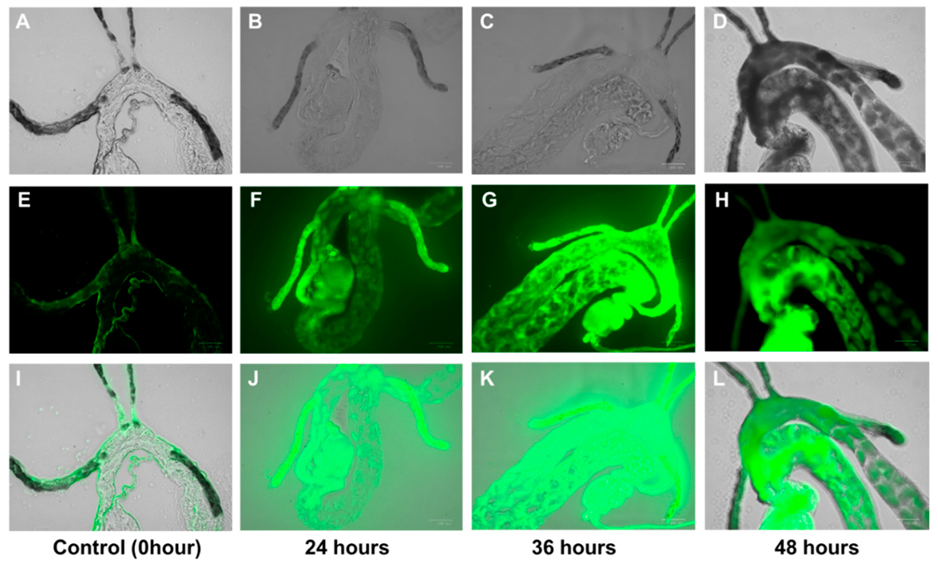
2.3. Tracing of Penicillin in D. citri
3. Discussion
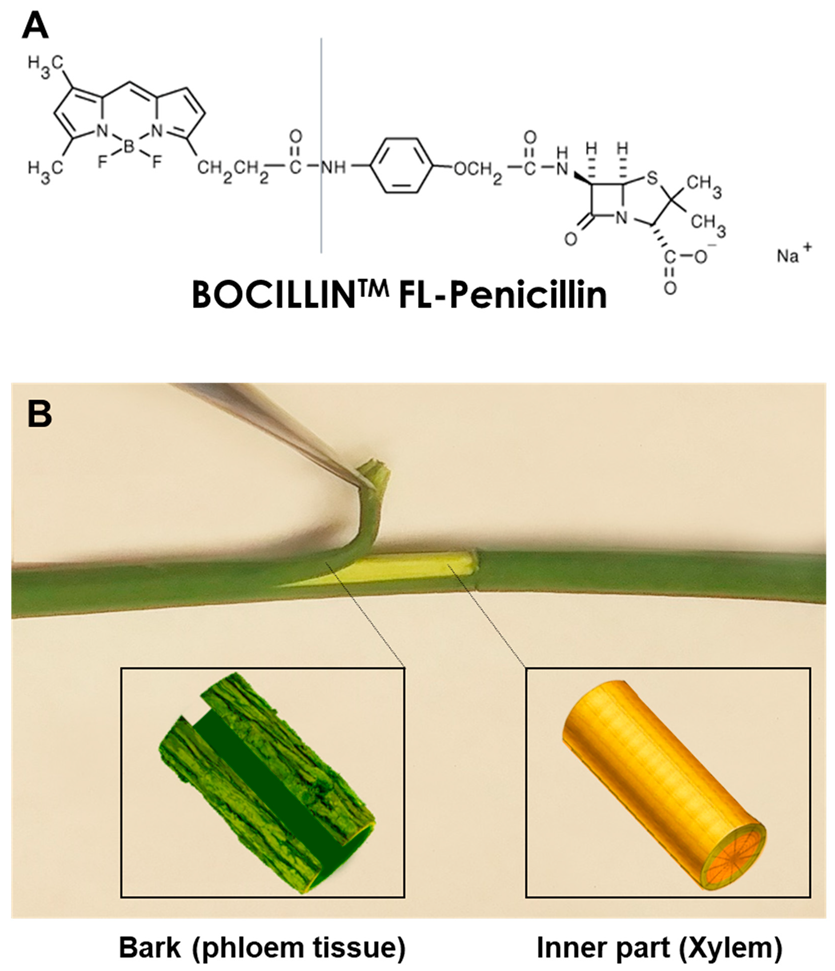
4. Materials and Methods
4.1. Plant Materials
4.2. Insect Culture
4.3. Application of Labeled Penicillin
4.4. Feeding of D. citri on Penicillin-Treated Citrus Cuttings
4.5. Microscopy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Grafton-Cardwell, E.E.; Stelinski, L.L.; Stansly, P.A. Biology and management of Asian citrus psyllid, vector of the Huanglongbing pathogens. Annu. Rev. Entomol. 2013, 58, 413–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinking, O.A. Diseases of economic plants in southern China. Philipp. Agric. 1919, 8, 109–135. [Google Scholar]
- Stansly, P.A.; Arevalo, H.A.; Qureshi, J.A.; Jones, M.M.; Hendricks, K.; Roberts, P.D.; Roka, F.M. Vector control and foliar nutrition to maintain economic sustainability of bearing citrus in Florida groves affected by huanglongbing. Pest Manag. Sci. 2014, 70, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Hao, G.; Stover, E.; Gupta, G. Overexpression of a Modified Plant Thionin Enhances Disease Resistance to Citrus Canker and Huanglongbing (HLB). Front. Plant. Sci. 2016, 7, 1078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaustein, R.A.; Lorca, G.L.; Teplitski, M. Challenges for managing Candidatus Liberibacter spp. (Huanglongbing Disease Pathogen): Current control measures and future directions. Phytopathology 2017, 108, 424–435. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, R.E.; van Vuuren, S.P. Decreases in fruit greening of sweet orange by trunk injections with tetracyclines. Plant Dis. Rep. 1970, 55, 747–750. [Google Scholar]
- Capoor, S.P.; Thirumal, M.J. Cure of greening affected citrus plants by chemotherapeutic agents. Plant Dis. Rep. 1973, 57, 160–163. [Google Scholar]
- Aubert, B.; Bove, J.M. Effect of Penicillin or tetracycline injections of citrus trees affected by greening disease under field conditions in reunion island. Proc. Eighth Conf. Int. Organ. Citrus Virol 1980, 8, 103–108. [Google Scholar]
- Zhao, X.Y. Citrus yellow shoot disease (Huanglongbing) in China—A review. In Proceedings of the International Society of Citriculture International Citrus/[International Citrus Congr, Tokyo, Japan, 9–12 November 1981. [Google Scholar]
- Zhang, M.; Yang, C.; Powell, C.A. Application of antibiotics for control of citrus Huanglongbing. Adv. Antibiot Antibodies 2015, 1, e101. [Google Scholar]
- Zhang, M.; Powell, C.A.; Guo, Y.; Doud, M.S.; Duan, Y.A. Graft-based chemotherapy method for screening effective molecules and rescuing Huanglongbing-affected citrus plants. Phytopathology 2012, 102, 567–574. [Google Scholar] [CrossRef] [Green Version]
- Shin, K.; Ascunce, M.S.; Narouei-Khandan, H.A.; Sun, X.; Jones, D.; Kolawole, O.O.; Goss, E.M.; van Bruggen, A.H.C. Effects and side effects of penicillin injection in Huanglongbing affected grapefruit trees. Crop. Prot. 2016, 90, 106–116. [Google Scholar] [CrossRef]
- Zhang, M.; Guo, Y.; Powell, C.A.; Doud, M.S.; Yang, C.; Duan, Y. Effective antibiotics against ‘Candidatus Liberibacter asiaticus’ in HLB-affected citrus plants identified via the graft-based evaluation. PLoS ONE 2014, 9, e111032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daniels, M.J. Editorial: Possible adverse effects of antibiotic therapy in plants. Clin. Infect. Dis. 1982, 4, S167–S170. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Jiang, J.; Wang, N. Control of citrus Huanglongbing via trunk injection of plant defense activators and antibiotics. Phytopathology 2018, 108, 186–195. [Google Scholar] [CrossRef]
- DeBoer, G.J.; Satchivi, N. Comparison of translocation properties of insecticides versus herbicides that leads to efficacious control of pests as specifically illustrated by isoclast active, a new insecticide and arylex active, a new herbicides. In Retention, Uptake, and Translocation of Agrochemicals in Plants; ACS Symposium Series; Myung, K., Satchivi, N.M., Kingston, C.K., Eds.; American Chemical Society: Washington, DC, USA, 2014; pp. 75–93. [Google Scholar]
- Zamora, S.M.A.; Escobar, F.R. Injector-size and the time of application affects uptake of tree trunk-injected solutions. Sci. Hortic. 2000, 84, 163–177. [Google Scholar] [CrossRef]
- Chaney, W.R. Anatomy and physiology related to chemical movement in trees. J. Arboric. 1986, 12, 85–91. [Google Scholar]
- Stone, M.R.L.; Butler, M.S.; Phetsang, W.; Cooper, M.A.; Blaskovich, M.A.T. Fluorescent antibiotics: new research tools to fight antibiotic resistance. Trends Biotechnol. 2018, 36, 523–536. [Google Scholar] [CrossRef]
- Zhao, G.; Meier, T.I.; Kahl, S.D.; Gee, K.R.; Blaszczak, L.C. BOCILLIN FL, a sensitive and commercially available reagent for detection of penicillin-binding proteins. Antimicrob. Agents Chemother. 1999, 43, 1124–1128. [Google Scholar] [CrossRef] [Green Version]
- Gessler, A.; Weber, P.; Schneider, S.; Rennenberg, H. Bidirectional exchange of amino compounds between phloem and xylem during long-distance transport in Norway spruce trees (Picea abies [L.] Karst). J. Exp. Bot. 2003, 54, 1389–1397. [Google Scholar] [CrossRef] [Green Version]
- Ebert, T.A.; Backus, E.A.; Shugart, H.J.; Rogers, M.E. Behavioral plasticity in probing by Diaphorina citri (Hemiptera, Liviidae): ingestion from phloem versus xylem is influenced by leaf age and surface. J. Insect Behav. 2018, 31, 119–137. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. Phloem-sap feeding by animals: Problems and solutions. J. Exp. Bot. 2006, 57, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Ascunce, M.S.; Shin, K.; Huguet-Tapia, J.C.; Poudel, R.; Garrett, K.A.; van Bruggen, A.H.C.; Goss, E.M. Penicillin trunk injection affects bacterial community structure in citrus trees. Microb. Ecol. 2019, 78, 457–469. [Google Scholar] [CrossRef] [PubMed]




© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Killiny, N.; Gonzalez-Blanco, P.; Santos-Ortega, Y.; Al-Rimawi, F.; Levy, A.; Hijaz, F.; Albrecht, U.; Batuman, O. Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin. Antibiotics 2019, 8, 262. https://doi.org/10.3390/antibiotics8040262
Killiny N, Gonzalez-Blanco P, Santos-Ortega Y, Al-Rimawi F, Levy A, Hijaz F, Albrecht U, Batuman O. Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin. Antibiotics. 2019; 8(4):262. https://doi.org/10.3390/antibiotics8040262
Chicago/Turabian StyleKilliny, Nabil, Pedro Gonzalez-Blanco, Yulica Santos-Ortega, Fuad Al-Rimawi, Amit Levy, Faraj Hijaz, Ute Albrecht, and Ozgur Batuman. 2019. "Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin" Antibiotics 8, no. 4: 262. https://doi.org/10.3390/antibiotics8040262
APA StyleKilliny, N., Gonzalez-Blanco, P., Santos-Ortega, Y., Al-Rimawi, F., Levy, A., Hijaz, F., Albrecht, U., & Batuman, O. (2019). Tracing Penicillin Movement in Citrus Plants Using Fluorescence-Labeled Penicillin. Antibiotics, 8(4), 262. https://doi.org/10.3390/antibiotics8040262