Evaluation by Flow Cytometry of Escherichia coli Viability in Lettuce after Disinfection



 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
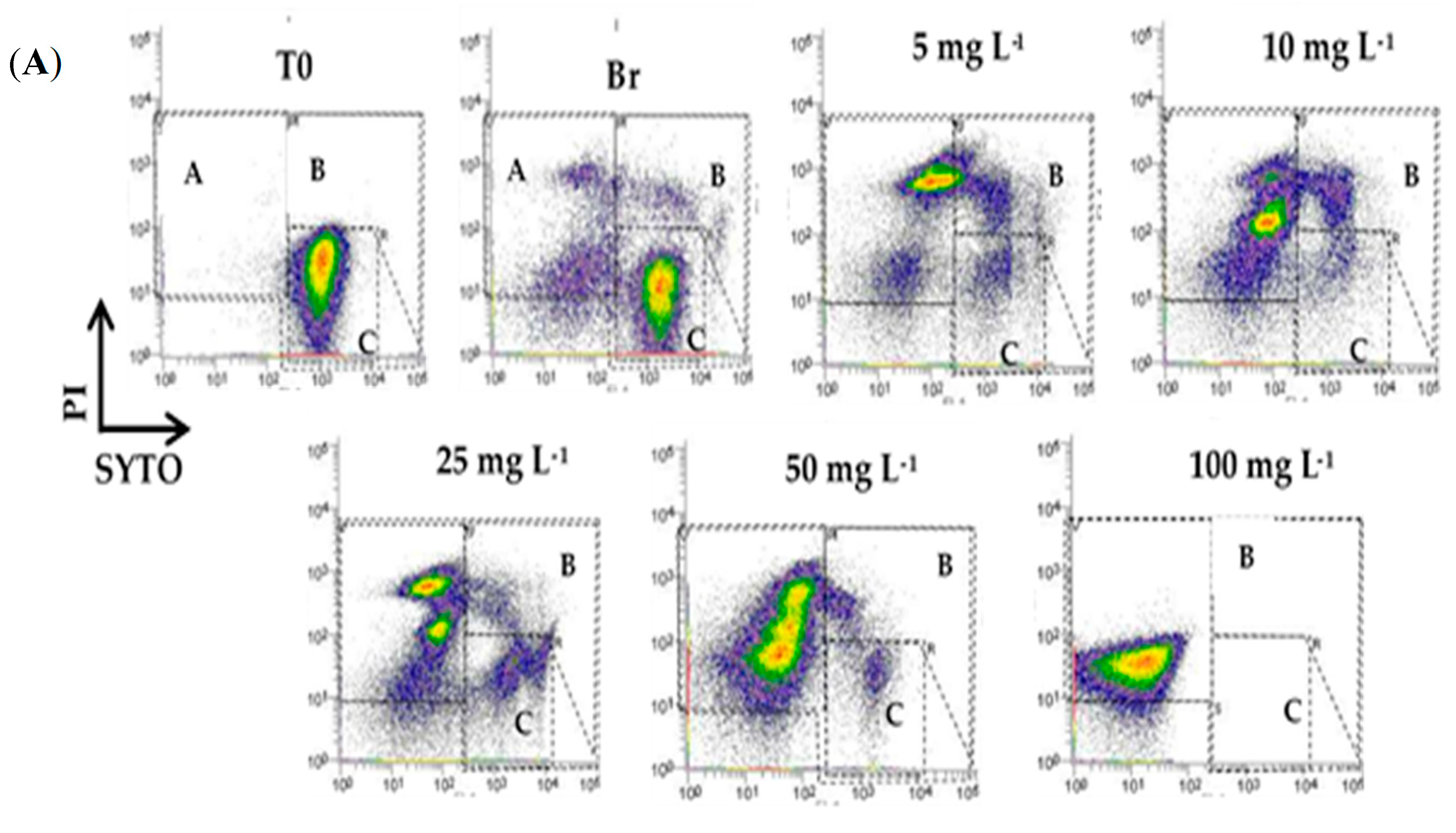
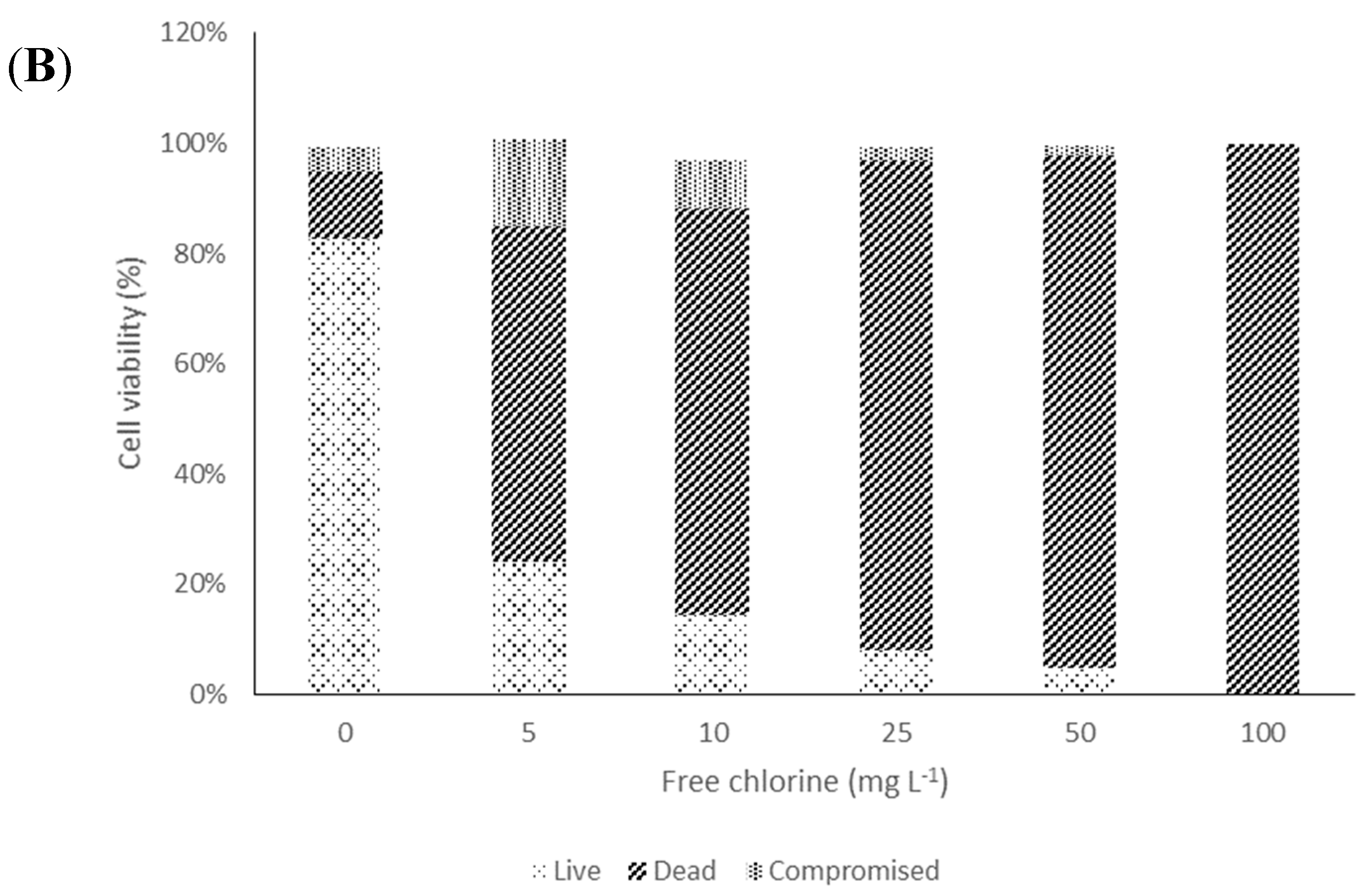
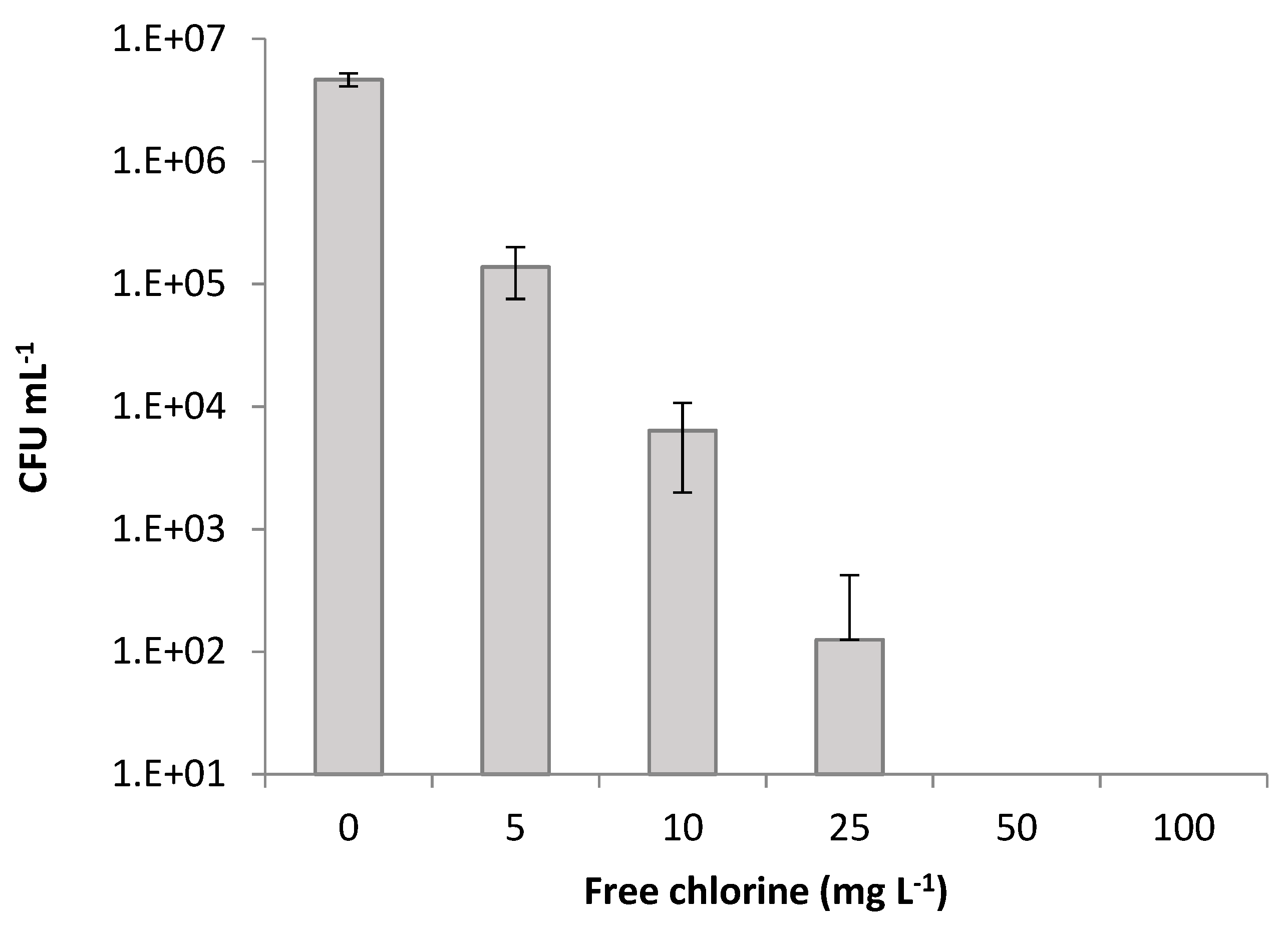
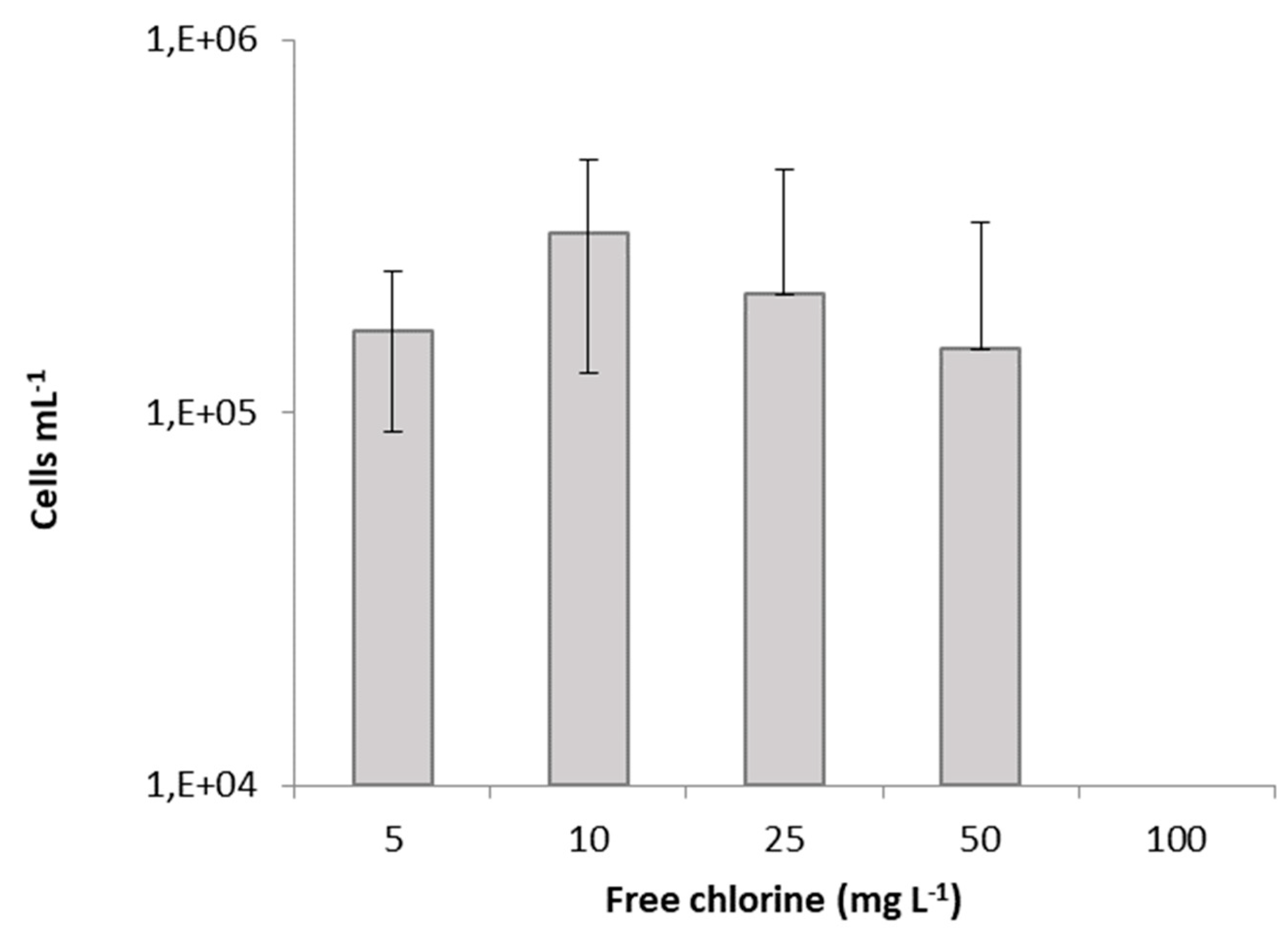
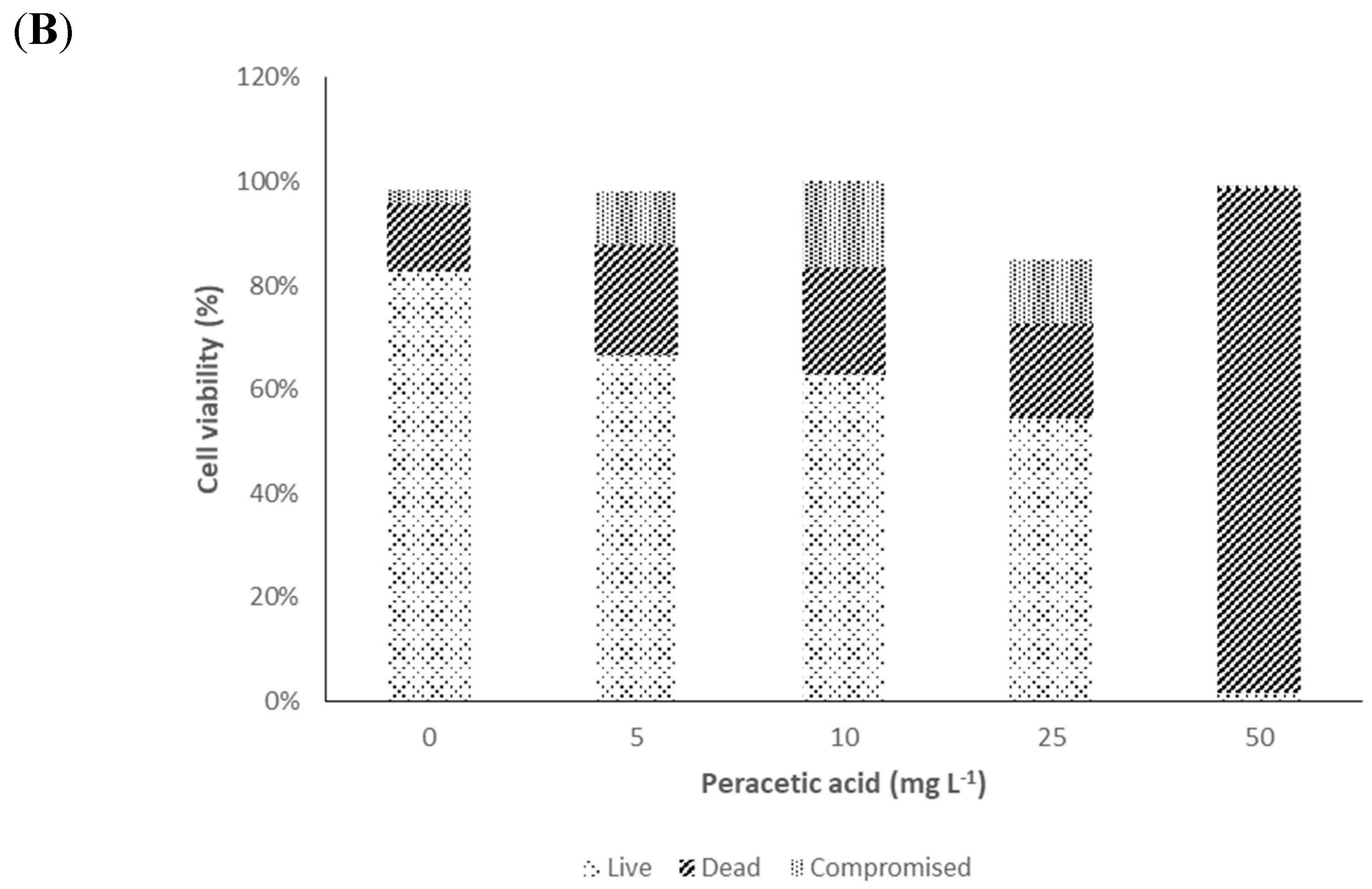
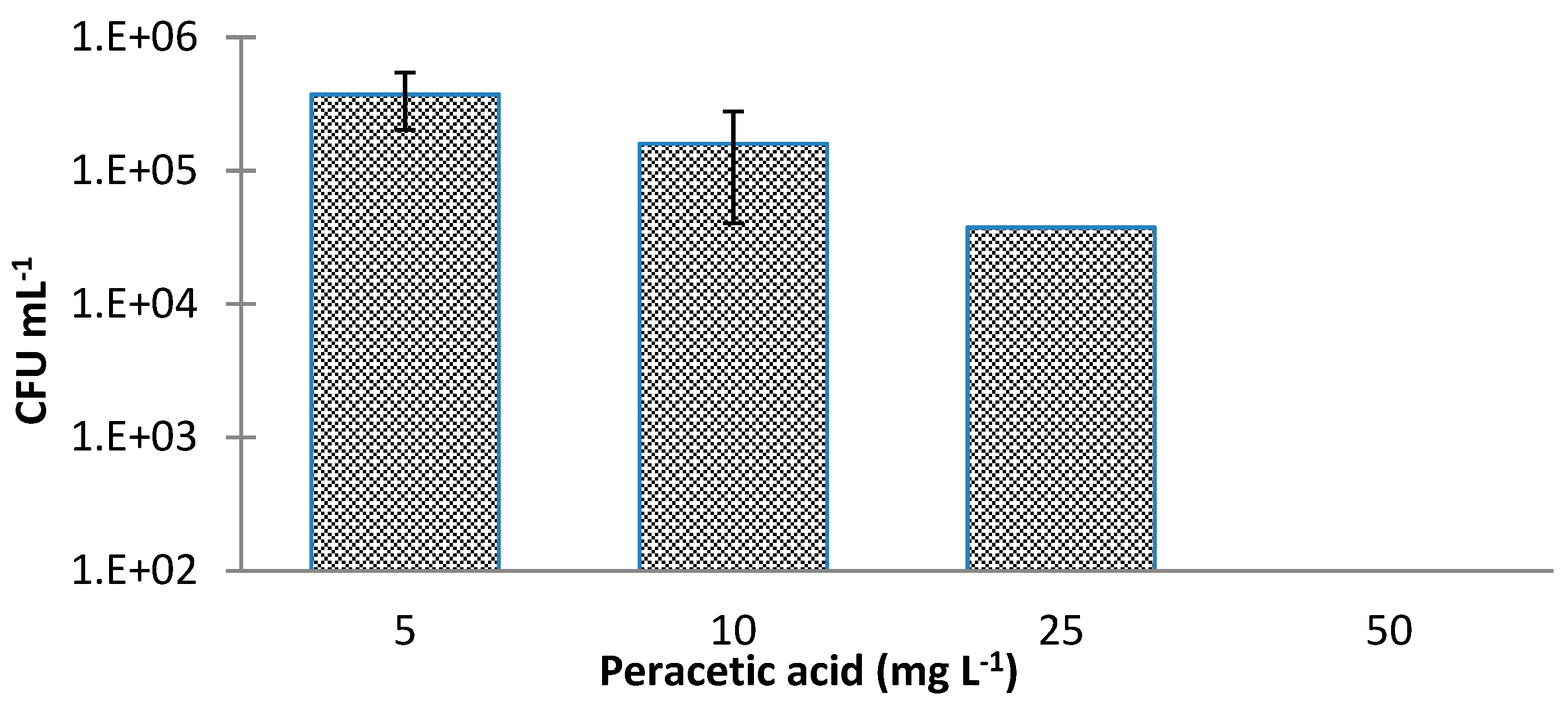
2. Results and Discussion
3. Materials and Methods
3.1. Bacterial Strain and Inoculum Preparation
3.2. Lettuce Preparation
3.3. Inoculation of Lettuce
3.4. Washing Solutions
3.5. Bacterial Viability Assays
3.5.1. Flow Cytometry Analysis
3.5.2. Standard Plate Count Method
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- European Food Safety Authority (EFSA). Panel on Biological Hazards (BIOHAZ). Scientific Opinion on the risk posed by pathogens in food of non-animal origin. Part 1 (outbreak data analysis and risk ranking of food/pathogen combinations). EFSA J. 2013, 11, 3025. [Google Scholar] [CrossRef] [Green Version]
- Herman, K.M.; Hall, A.J.; Gould, L.H. Outbreaks attributed to fresh leafy vegetables, United States, 1973–2012. Epidemiol. Infect. 2015, 143, 3011–3021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franz, E.; Visser, A.A.; Van Diepeningen, A.D.; Klerks, M.M.; Termorshuizen, A.J.; van Bruggen, A.H. Quantification of contamination of lettuce by GFP-expressing Escherichia coli O157:H7 and Salmonella enterica serovar Typhimurium. Food Microbiol. 2007, 24, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.; Jang, H.; Matthews, K.R. Effect of the food production chain from farm practices to vegetable processing on outbreak incidence. Microb. Biotechnol. 2014, 7, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control (CDC). Multistate Outbreak of E. coli O157:H7 Infections Linked to Romaine Lettuce (Final Update). 2018. Available online: https://www.cdc.gov/ecoli/2018/o157h7-04-18/index.html (accessed on 24 January 2019).
- Back, K.-H.; Ha, J.-W.; Kang, D.-H. Effect of hydrogen peroxide vapor treatment for inactivating Salmonella Typhimurium, Escherichia coli O157: H7 and Listeria monocytogenes on organic fresh lettuce. Food Control 2014, 44, 78–85. [Google Scholar] [CrossRef]
- World Health Organization & Food and Agriculture Organization of the United Nations. Benefits and risks of the use of chlorine-containing disinfectants in food production and food processing. In Proceedings of the Benefits and risks of the use of chlorine-containing disinfectants in food production and food processing: Report of a joint FAO/WHO expert meeting, Ann Arbor, MI, USA, 27–30 May 2008. [Google Scholar]
- Richardson, S.D.; Thruston, A.; Caughran, T.; Collette, T.; Patterson, K.; Lykins, B. Chemical by-products of chlorine and alternative disinfectants. Food Technol. 1998, 52, 58–61. [Google Scholar]
- Nou, X.; Lu, Y. Whole-Leaf Wash Improves Chlorine Efficacy for Microbial Reduction and Prevents Pathogen Cross-Contamination during Fresh-Cut Lettuce Processing. J. Food Sci. 2010, 75, M283–M290. [Google Scholar] [CrossRef]
- Anonymous. Commodity specific food safety guidelines for the lettuce and leafy greens supply chain. 2006. Available online: http://www.fda.gov/downloads/Food/GuidanceRegulation/UCM169008.pdf (accessed on 30 September 2019).
- Fu, T.L.; Li, Y.; Awad, D.; Zhou, T.-Y.; Liu, L. Factors affecting the performance and monitoring of a chlorine wash in preventing Escherichia coli O157:H7 cross-contamination during postharvest washing of cut lettuce. Food Control 2018, 94, 212–221. [Google Scholar] [CrossRef]
- Ölmez, H.; Kretzschamr, U. Potential alternative disinfection methods for organic fresh-cut industry for minimizing water consumption and environmental impact. LWT-Food Sci. Technol. 2009, 42, 686–693. [Google Scholar] [CrossRef]
- Carrasco, G.; Moggia, C.; Osses, I.J.; Álvaro, J.E.; Urrestarazu, M. Use of peroxyacetic acid as green chemical on yield and sensorial quality in watercress (Nasturtium officinale R. Br.) under soilless culture. Int. J. Mol. Sci. 2011, 12, 9463–9470. [Google Scholar] [CrossRef]
- Santoro, D.; Gehr, R.; Bartrand, T.A.; Liberti, L.; Notarnicola, M.; Dell’Erba, A.; Falsanisi, D.; Haas, C.N. Wastewater disinfection by peracetic acid: Assessment of models for tracking residual measurements and inactivation. Water Environ. Res. 2007, 79, 775–787. [Google Scholar] [CrossRef] [PubMed]
- Ding, T.; Suo, Y.; Xiang, Q.; Zhao, X.; Chen, S.; Ye, X.; Liu, D. Significance of viable but nonculturable Escherichia coli: Induction, detection, and control. J. Microbiol. Biotechnol. 2017, 27, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Ayari, S.; Dussault, D.; Hayouni, E.A.; Hamdi, M.; Lacroix, M. Radiation tolerance of Bacillus cereus pre-treated with carvacrol alone or in combination with nisin after exposure to single and Multiple sub-lethal radiation treatment. Food Control 2013, 32, 693–701. [Google Scholar] [CrossRef]
- Schenk, M.; Raffellini, S.; Guerrero, S.; Blanco, G.A.; Alzamora, S.M. Inactivation of Escherichia coli, Listeria innocua and Saccharomyces cerevisiae by UV-C light: Study of cell injury by flow cytometry. LWT- Food Sci. Technol. 2011, 44, 191–198. [Google Scholar] [CrossRef]
- Oliver, J.D. The viable but nonculturable state in bacteria. J. Microbiol. 2005, 43, 93–100. [Google Scholar]
- Hewitt, C.J.; Nebe-Von-Caron, G. The application of multiparameter flow cytometry to monitor individual microbial cell physiological state. Adv. Biochem. Eng/Biot. 2004, 89, 197–223. [Google Scholar]
- Saegeman, V.S.M.; De Vos, R.; Tebaldi, N.D.; van der Wolf, J.M.; Bergervoet, J.H.W.; Verhaegen, J.; Lismont, D.; Verduyckt, B.; Ectors, N.L. Flow cytometric viability assessment and transmission electron microscopic morphological study of bacteria in glycerol. Microsc. Microanal. 2007, 13, 18–29. [Google Scholar] [CrossRef] [Green Version]
- Barbesti, S.; Citterio, S.; Labra, M.; Baroni, M.D.; Neri, M.G.; Sgorbati, S. Two and three-color fluorescence flow cytometric analysis of immunoidentified viable bacteria. Cytometry 2000, 40, 214–218. [Google Scholar] [CrossRef]
- Ben-Amor, K.; Heilig, H.; Smidt, H.; Vaughan, E.E.; Abee, T.; de Vos, W.M. Genetic diversity of viable, injured, and dead fecal bacteria assessed by fluorescence-activated cell sorting and 16S rRNA gene analysis. Appl. Environ. Microbiol. 2005, 71, 4679–4689. [Google Scholar] [CrossRef] [Green Version]
- Berney, M.; Hammes, F.; Bosshard, F.; Weilenmann, H.U.; Egli, T. Assessment and interpretation of bacterial viability by using the LIVE/DEAD BacLight Kit in combination with flow cytometry. Appl. Environ. Microbiol. 2007, 73, 3283–3290. [Google Scholar] [CrossRef] [Green Version]
- Booyens, J.; Thantsha, M.S. Fourier transform infra-red spectroscopy and flow cytometric assessment of the antibacterial mechanism of action of aqueous extract of garlic (Allium sativum) against selected probiotic Bifidobacterium strains. Complement. Altern. Med. 2014, 14, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayouni, E.A.; Bouix, M.; Abedrabba, M.; Leveau, J.-Y.; Hamdi, M. Mechanism of action of Melaleuca armillaris (Sol. Ex Gaertu) Sm. essential oil on six LAB strains as assessed by multiparametric flow Cytometry and automated microtiter-based assay. Food Chem. 2008, 111, 707–718. [Google Scholar] [CrossRef]
- Massicotte, R.; Mafu, A.A.; Deshaies, F.; Pichette, G.; Pierre, B. Comparison between Flow Cytometry and Traditional Culture Methods for Efficacy Assessment of Six Disinfectant Agents against Nosocomial Bacterial Species. Front. Microbiol. 2017, 8, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hawkins, C.L.; Davies, M.J. Hypochlorite-induced damage to DNA, RNA, and polynucleotides: Formation of chloramines and nitrogen-centered radicals. Chem. Res. Toxicol. 2002, 15, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Phe, M.H.; Dossot, M.; Guilloteau, H.; Block, J.C. Highly chlorinated Escherichia coli cannot be stained by propidium iodide. Can. J. Microbiol. 2007, 53, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xi, L.; Yahong, W.; Jie, Z.; Chengsong, Y.; Xianping, L.; Lizheng, G.; Shenghua, Z.; Xin, Y. Induction of E. coli into a VBNC state through chlorination/chloramination and differences in characteristics of the bacterium between states. Water Res. 2018, 142, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Ayala-Zavala, J.F.; González- Aguilar, G.A. Use of Additives to Preserve the Quality of Fresh-Cut Fruits and Vegetable. In Advances in Fresh-Cut Fruits and Vegetables Processing; Martin-Belloso, O., Fortuny, R.S., Eds.; CRC Press: Boca Raton, FL, USA, 2011; Chapter 9; pp. 232–249. [Google Scholar]
- Van Haute, S.; Sampers, I.; Holvoet, K.; Uyttendaele, M. Physicochemical quality and chemical safety of chlorine as a reconditioning agent and wash water disinfectant for fresh-cut lettuce washing. Appl. Environ. Microbiol. 2013, 79, 2850–2861. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Brackett, R.E.; Chen, J.; Beuchat, L.R. Survival and growth of Escherichia coli 0157:H7 inoculated onto cut lettuce before or after heating in chlorinated water, followed by storage at 5 °C or 15 °C. J. Food Prot. 2001, 64, 305–309. [Google Scholar]
- Beuchat, L.R. Survival of Enterohemorrhagic Escherichia coli O157:H7 in bovine feces applied to lettuce and the effectiveness of chlorinated water as a disinfectant. J. Food Prot. 1999, 62, 845–849. [Google Scholar] [CrossRef]
- Behrsing, J.; Winkler, S.; Franz, P.; Premier, R. Efficacy of chlorine for inactivation of Escherichia coli on vegetables. Postharvest Biol. Technol. 2000, 19, 187–192. [Google Scholar] [CrossRef]
- Pan, X.; Nakano, H. Effects of Chlorine-Based Antimicrobial Treatments on the Microbiological Qualities of Selected Leafy Vegetables and Wash Water. Food Sci. Technol. Res. 2014, 20, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Seo, K.H.; Frank, J.F. Attachment of Escherichia coli O157:H7 to Lettuce Leaf Surface and Bacterial Viability in Response to Chlorine Treatment as Demonstrated by Using Confocal Scanning Laser Microscopy. J. Food Prot. 1999, 62, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Niemira, B.A. Irradiation compared with chlorination for elimination of Escherichia coli O157:H7 internalized in lettuce leaves: Influence of lettuce variety. J. Food Sci. 2008, 73, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Vandekinderen, I.; Devlieghere, F.; De Meulenaer, B.; Veramme, K.; Ragaert, P.; Van Camp, J. Impact of decontamination agents and a packaging delay on the respiration rate of fresh-cut produce. Postharvest Biol. Technol. 2008, 49, 277–282. [Google Scholar] [CrossRef]
- Zoellner, C.; Aguayo-Acosta, A.; Siddiqui, M.W.; Dávila-Aviña, J.E. Peracetic Acid in Disinfection of Fruits and Vegetables. In Postharvest Disinfection of Fruits and Vegetables; Wasim Siddiqui, M., Ed.; Academic Press: London, UK, 2018; Chapter 2; pp. 53–66. [Google Scholar]
- Park, C.M.; Beuchat, L.R. Evaluation of sanitizers for killing Escherichia coli O157:H7, Salmonella and naturally occurring microorganisms on cantaloupes, honeydew melons, and asparagus. Dairy Food Environ. Sanit. 1999, 19, 842–847. [Google Scholar]
- Keeratipibul, S.; Phewpan, A.; Lursinsap, C. Prediction of coliforms and Escherichia coli on tomato fruit and lettuce leaves after sanitizing by using artificial neural networks. LWT-Food Sci. Technol. 2011, 44, 130–138. [Google Scholar] [CrossRef]
- Kitis, M. Disinfection of wastewater with peracetic acid: A review. Environ. Int. 2004, 30, 47–55. [Google Scholar] [CrossRef]
- Viola, K.; Rodrigues, E.; Tanomaru-Filho, M.; Carlos, I.; Ramos, S.; Guerreiro-Tanomaru, J.; Faria, G. Cytotoxicity of peracetic acid: Evaluation of effects on metabolism, structure and cell death. Int. Endod. J. 2018, 51, e264–e277. [Google Scholar] [CrossRef]
- Pechacek, N.; Osorio, M.; Caudill, J.; Peterson, B. Evaluation of the toxicity data for peracetic acid in deriving occupational exposure limits: A minireview. Toxicol. Lett. 2015, 233, 45–57. [Google Scholar] [CrossRef]
- Ölmez, H.; Temur, S.D. Effects of different sanitizing treatments on biofilms and attachment of Escherichia coli and Listeria monocytogenes on green leaf lettuce. LWT—Food Sci. Technol. 2010, 43, 964–970. [Google Scholar] [CrossRef]
- Kroupitski, Y.; Pinto, R.; Belausov, E.; Sela, S. Distribution of Salmonella Typhimurium in romaine lettuce leaves. Food Microbiol. 2011, 28, 990–997. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teixeira, P.; Fernandes, B.; Silva, A.M.; Dias, N.; Azeredo, J. Evaluation by Flow Cytometry of Escherichia coli Viability in Lettuce after Disinfection. Antibiotics 2020, 9, 14. https://doi.org/10.3390/antibiotics9010014
Teixeira P, Fernandes B, Silva AM, Dias N, Azeredo J. Evaluation by Flow Cytometry of Escherichia coli Viability in Lettuce after Disinfection. Antibiotics. 2020; 9(1):14. https://doi.org/10.3390/antibiotics9010014
Chicago/Turabian StyleTeixeira, Pilar, Bruna Fernandes, Ana Margarida Silva, Nicolina Dias, and Joana Azeredo. 2020. "Evaluation by Flow Cytometry of Escherichia coli Viability in Lettuce after Disinfection" Antibiotics 9, no. 1: 14. https://doi.org/10.3390/antibiotics9010014
APA StyleTeixeira, P., Fernandes, B., Silva, A. M., Dias, N., & Azeredo, J. (2020). Evaluation by Flow Cytometry of Escherichia coli Viability in Lettuce after Disinfection. Antibiotics, 9(1), 14. https://doi.org/10.3390/antibiotics9010014