Clonal Diversity, Biofilm Formation, and Antimicrobial Resistance among Stenotrophomonas maltophilia Strains from Cystic Fibrosis and Non-Cystic Fibrosis Patients

 ,
, 


Abstract
:1. Introduction
2. Results
2.1. The S. maltophilia Population Shows a High Genetic Heterogeneity
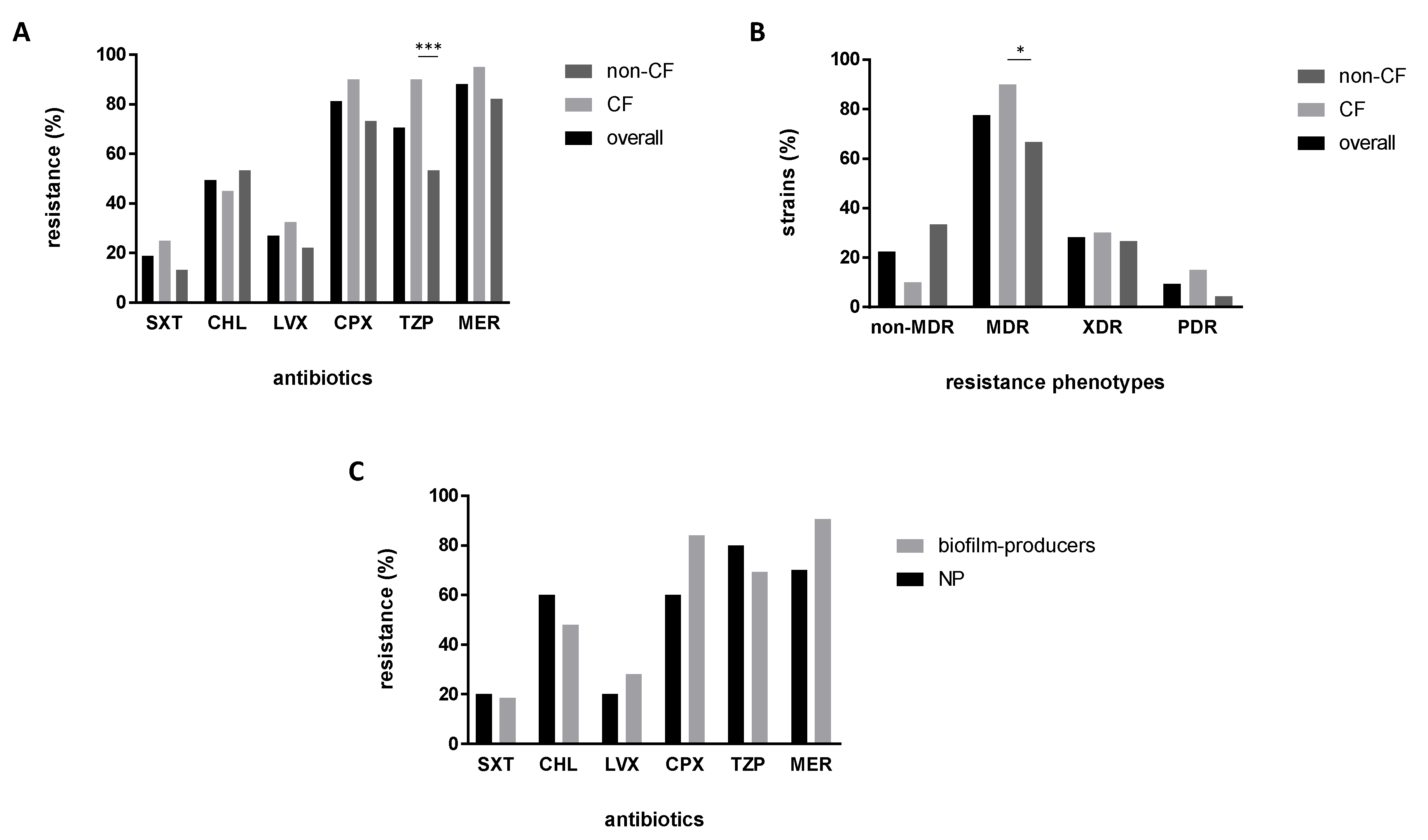
2.2. Antibiotic Resistance Is More Prevalent in CF Strains
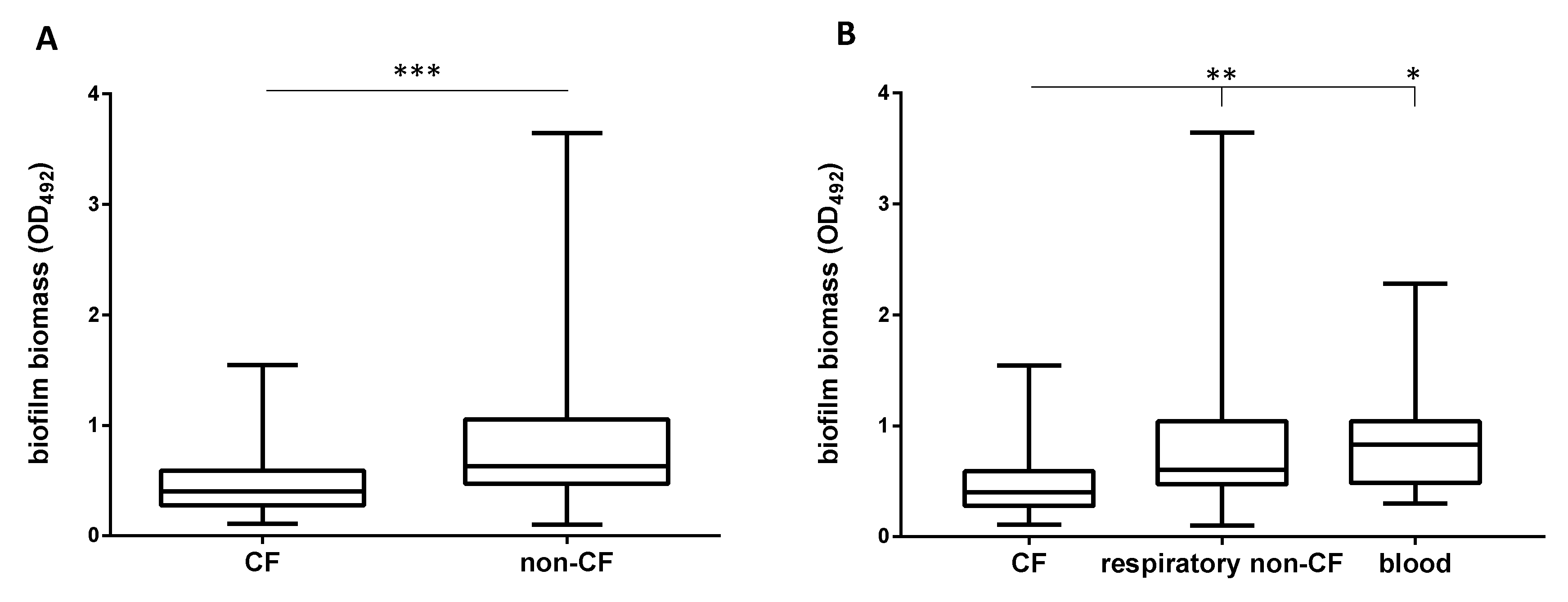
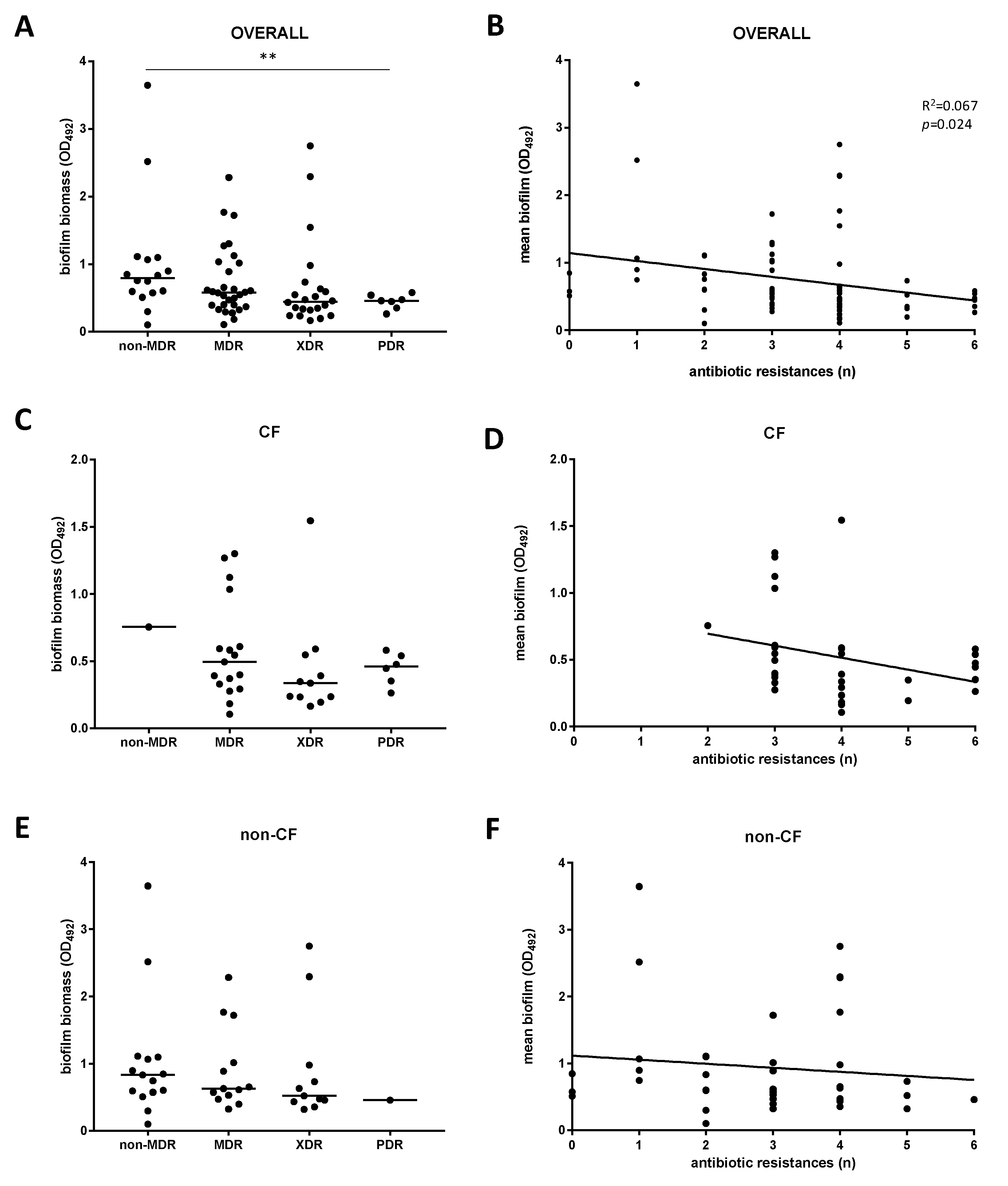
2.3. Non-CF Strains Are Significantly More Efficient Than CF Strains in Forming Biofilm
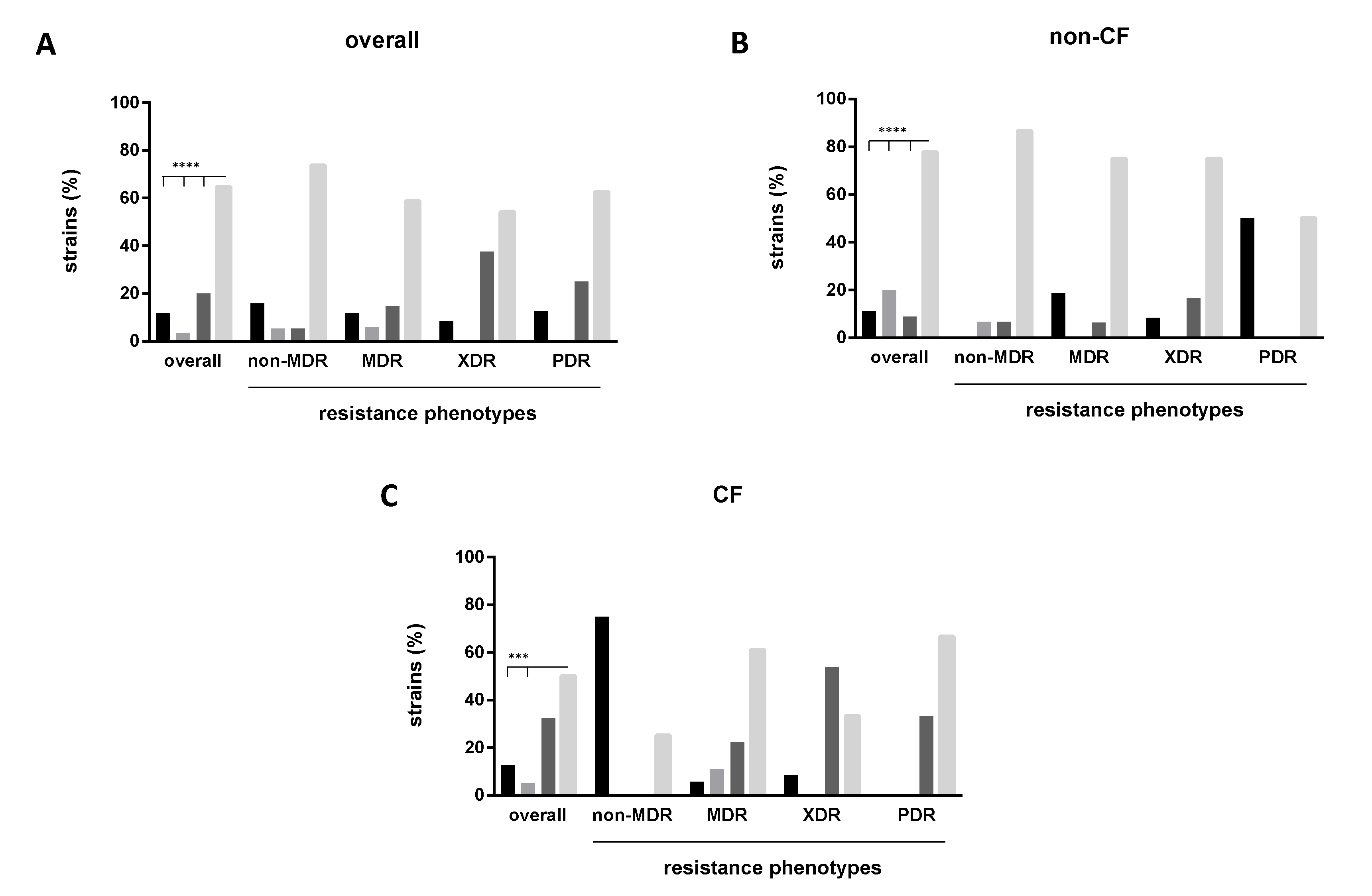
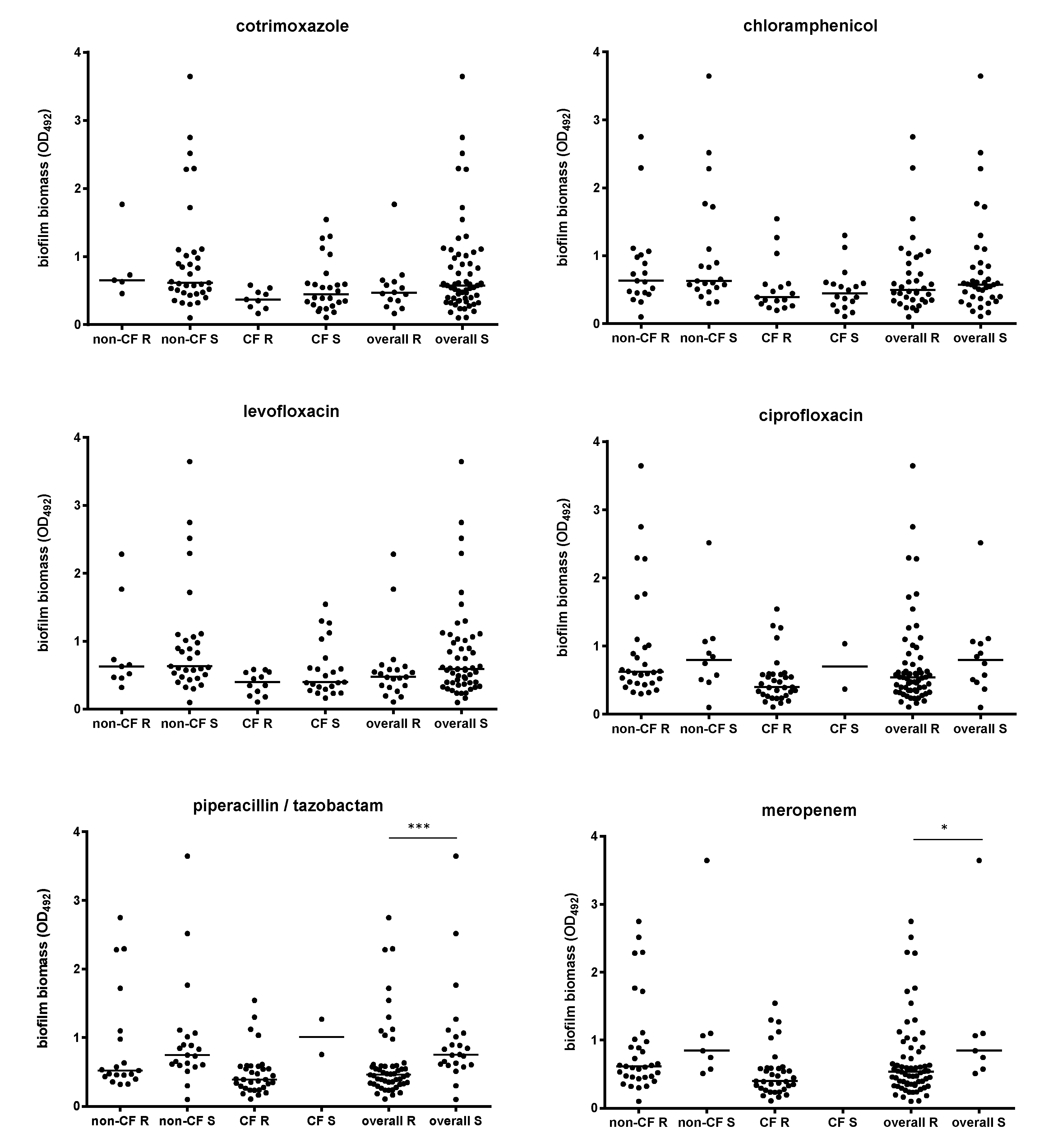
2.4. Biofilm Formation and Antibiotic Resistance Are Inversely Related
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Genetic Relatedness and Cluster Analysis
4.3. Antibiotic Susceptibility Testing
4.4. Biofilm Formation Assay
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Looney, W.J.; Narita, M.; Mühlemann, K. Stenotrophomonas maltophilia: An emerging opportunist human pathogen. Lancet Infect. Dis. 2009, 9, 312–323. [Google Scholar] [CrossRef]
- Brooke, J.S. Stenotrophomonas maltophilia: An emerging global opportunistic pathogen. Clin. Microbiol. Rev. 2012, 25, 2–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, V.; Yau, Y.; Prasad, S.; Lu, A.; Atenafu, E.; Crandall, I.; Tom, S.; Tullis, E.; Ratjen, F. Stenotrophomonas maltophilia in cystic fibrosis: Serologic response and effect on lung disease. Am. J. Respir. Crit. Care Med. 2011, 183, 635–640. [Google Scholar] [CrossRef] [PubMed]
- Goss, C.H.; Mayer-Hamblett, N.; Aitken, M.L.; Rubenfeld, G.D.; Ramsey, B.W. Association between Stenotrophomonas maltophilia and lung function in cystic fibrosis. Thorax 2004, 59, 955–959. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, V.; Atenafu, E.G.; Lu, A.; Yau, Y.; Tullis, E.; Ratjen, F. Chronic Stenotrophomonas maltophilia infection and mortality or lung transplantation in cystic fibrosis patients. J. Cyst. Fibros. 2013, 12, 482–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raidt, L.; Idelevich, E.A.; Dübbers, A.; Küster, P.; Drevinek, P.; Peters, G.; Kahl, B.C. Increased prevalence and resistance of important pathogens recovered from respiratory specimens of cystic fibrosis patients during a decade. Pediatr. Infect. Dis. J. 2015, 34, 700–705. [Google Scholar] [CrossRef]
- Di Bonaventura, G.; Prosseda, G.; Del Chierico, F.; Cannavacciuolo, S.; Cipriani, P.; Petrucca, A.; Superti, F.; Ammendolia, M.G.; Concato, C.; Fiscarelli, E.; et al. Molecular characterization of virulence determinants of Stenotrophomonas maltophilia strains isolated from patients affected by cystic fibrosis. Int. J. Immunopathol. Pharmacol. 2007, 20, 529. [Google Scholar] [CrossRef] [Green Version]
- Di Bonaventura, G.; Spedicato, I.; D’Antonio, D.; Robuffo, I.; Piccolomini, R. Biofilm formation by Stenotrophomonas maltophilia: Modulation by quinolones, trimethoprim-sulfamethoxazole, and ceftazidime. Antimicrob. Agents Chemother. 2004, 48, 151–160. [Google Scholar] [CrossRef] [Green Version]
- Sun, E.; Liang, G.; Wang, L.; Wei, W.; Lei, M.; Song, S.; Han, R.; Wang, Y.; Qi, W. Antimicrobial susceptibility of hospital acquired Stenotrophomonas maltophilia isolate biofilms. Braz. J. Infect. Dis. 2016, 20, 365–373. [Google Scholar] [CrossRef] [Green Version]
- Pompilio, A.; Crocetta, V.; Confalone, P.; Nicoletti, M.; Petrucca, A.; Guarnieri, S.; Fiscarelli, E.; Savini, V.; Piccolomini, R.; Di Bonaventura, G. Adhesion to and biofilm formation on IB3-1 bronchial cells by Stenotrophomonas maltophilia isolates from cystic fibrosis patients. BMC Microbiol. 2010, 10, 102. [Google Scholar] [CrossRef] [Green Version]
- Pompilio, A.; Crocetta, V.; Ghosh, D.; Chakrabarti, M.; Gherardi, G.; Vitali, L.A.; Fiscarelli, E.; Di Bonaventura, G. Stenotrophomonas maltophilia phenotypic and genotypic diversity during a 10-year colonization in the lungs of a cystic fibrosis patient. Front. Microbiol. 2016, 7, 1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Abreu, P.M.; Farias, P.G.; Paiva, G.S.; Almeida, A.M.; Morais, P.V. Persistence of microbial communities including Pseudomonas aeruginosa in a hospital environment: A potential health hazard. BMC Microbiol. 2014, 14, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, A.; Pompilio, A.; Bettua, C.; Crocetta, V.; Giacobazzi, E.; Fiscarelli, E.; Jousson, O.; Di Bonaventura, G. Evolution of Stenotrophomonas maltophilia in cystic fibrosis lung over chronic infection: A genomic and phenotypic population study. Front. Microbiol. 2017, 8, 1590. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, M.B. Antibiotic resistance in the opportunistic pathogen Stenotrophomonas maltophilia. Front. Microbiol. 2015, 6, 658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, Y.D.; Jeong, W.Y.; Kim, M.H.; Jung, I.Y.; Ahn, M.Y.; Ann, H.W.; Ahn, J.Y.; Han, S.H.; Choi, J.Y.; Song, Y.G.; et al. Risk factors for mortality in patients with Stenotrophomonas maltophilia bacteremia. Medicine 2016, 95, e4375. [Google Scholar] [CrossRef]
- Kwon, A.S.; Park, G.C.; Ryu, S.Y.; Lim, D.H.; Lim, D.Y.; Choi, C.H.; Park, Y.; Lim, Y. Higher biofilm formation in multidrug-resistant clinical isolates of Staphylococcus aureus. Int. J. Antimicrob. Agents 2008, 32, 68–72. [Google Scholar] [CrossRef]
- Shrestha, L.B.; Bhattarai, N.R.; Khanal, B. Comparative evaluation of methods for the detection of biofilm formation in coagulase-negative staphylococci and correlation with antibiogram. Infect. Drug Resist. 2018, 11, 607–613. [Google Scholar] [CrossRef] [Green Version]
- Badave, G.K.; Kulkarni, D. Biofilm producing multidrug resistant Acinetobacter baumannii: An emerging challenge. J. Clin. Diagn. Res. 2015, 9, DC08. [Google Scholar] [CrossRef]
- Sahal, G.; Bilkay, I.S. Multi drug resistance in strong biofilm forming clinical isolates of Staphylococcus epidermidis. Braz. J. Microbiol. 2014, 45, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Krzyściak, P.; Chmielarczyk, A.; Pobiega, M.; Romaniszyn, D.; Wójkowska-Mach, J. Acinetobacter baumannii isolated from hospital-acquired infection: Biofilm production and drug susceptibility. APMIS 2017, 125, 1017–1026. [Google Scholar] [CrossRef]
- Qi, L.; Li, H.; Zhang, C.; Liang, B.; Li, J.; Wang, L.; Du, X.; Liu, X.; Qiu, S.; Song, H. Relationship between antibiotic resistance, biofilm formation, and biofilm-specific resistance in Acinetobacter baumannii. Front. Microbiol. 2016, 7, 483. [Google Scholar] [CrossRef] [Green Version]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 29th ed.; CLSI Supplement M100; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2019. [Google Scholar]
- Stepanovic, S.; Vukovic, D.; Hola, V.; Di Bonaventura, G.; Djukic, S.; Cirkovic, I.; Ruzicka, F. Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by staphylococci. APMIS 2007, 115, 891–899. [Google Scholar] [CrossRef]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Harms, A.; Maisonneuve, E.; Gerdes, K. Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 2016, 354, aaf4268. [Google Scholar] [CrossRef]
- Ciofu, O.; Rojo-Molinero, E.; Macià, M.D.; Oliver, A. Antibiotic treatment of biofilm infections. APMIS 2017, 125, 304–319. [Google Scholar] [CrossRef]
- Azeredo, J.; Azevedo, N.F.; Briandet, R.; Cerca, N.; Coenye, T.; Costa, A.R.; Desvaux, M.; Di Bonaventura, G.; Hébraud, M.; Jaglic, Z.; et al. Critical review on biofilm methods. Crit. Rev. Microbiol. 2017, 43, 313–351. [Google Scholar] [CrossRef] [Green Version]
- Tanimoto, K. Stenotrophomonas maltophilia strains isolated from a university hospital in Japan: Genomic variability and antibiotic resistance. J. Med. Microbiol. 2013, 62, 565–570. [Google Scholar] [CrossRef] [Green Version]
- Neela, V.; Rankouhi, S.Z.; van Belkum, A.; Goering, R.V.; Awang, R. Stenotrophomonas maltophilia in Malaysia: Molecular epidemiology and trimethoprim-sulfamethoxazole resistance. Int. J. Infect. Dis. 2012, 16, e603–e607. [Google Scholar] [CrossRef] [Green Version]
- Prunier, A.L.; Malbruny, B.; Laurans, M.; Brouard, J.F.; Leclercq, R. High rate of macrolide resistance in Staphylococcus aureus strains from patients with cystic fibrosis reveals high proportions of hypermutable strains. J. Infect. Dis. 2003, 187, 1709–1716. [Google Scholar] [CrossRef] [Green Version]
- Molin, S.; Tolker-Nielsen, T. Gene transfer occurs with enhanced efficiency in biofilms and induces enhanced stabilisation of the biofilm structure. Curr. Opin. Biotechnol. 2003, 14, 255–261. [Google Scholar] [CrossRef]
- Lee, H.W.; Koh, Y.M.; Kim, J.; Lee, J.C.; Lee, Y.C.; Seol, S.Y.; Cho, D.T.; Kim, J. Capacity of multidrug-resistant clinical isolates of Acinetobacter baumannii to form biofilm and adhere to epithelial cell surfaces. Clin. Microbiol. Infect. 2008, 14, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Safdar, A.; Rodriguez, G.; Balakrishnan, M.; Tarrand, J.; Rolston, K. Changing trends in etiology of bacteremia in patients with cancer. Eur. J. Clin. Microbiol. Infect. Dis. 2006, 25, 522–526. [Google Scholar] [CrossRef]
- Barbolla, R.; Catalano, M.; Orman, B.E.; Famiglietti, A.; Vay, C.; Smayevsky, J.; Centrón, D.; Piñeiro, S.A. Class 1 integrons increase trimethoprim-sulfamethoxazole MICs against epidemiologically unrelated Stenotrophomonas maltophilia isolates. Antimicrob. Agents Chemother. 2004, 48, 666–669. [Google Scholar] [CrossRef] [Green Version]
- De Dios Caballero, J.; del Campo, R.; Royuela, A.; Solé, A.; Máiz, L.; Olveira, C.; Quintana-Gallego, E.; de Gracia, J.; Cobo, M.; de la Pedrosa, E.G.; et al. Bronchopulmonary infection–colonization patterns in Spanish cystic fibrosis patients: Results from a national multicenter study. J. Cyst. Fibros. 2016, 15, 357–365. [Google Scholar] [CrossRef] [Green Version]
- Valenza, G.; Tappe, D.; Turnwald, D.; Frosch, M.; König, C.; Hebestreit, H.; Abele-Horn, M. Prevalence and antimicrobial susceptibility of microorganisms isolated from sputa of patients with cystic fibrosis. J. Cyst. Fibros. 2008, 7, 123–127. [Google Scholar] [CrossRef]
- Chang, Y.T.; Lin, C.Y.; Chen, Y.H.; Hsueh, P.R. Update on infections caused by Stenotrophomonas maltophilia with particular attention to resistance mechanisms and therapeutic options. Front. Microbiol. 2015, 6, 893. [Google Scholar] [CrossRef]
- Díez-Aguilar, M.; Ekkelenkamp, M.; Morosini, M.I.; Merino, I.; de Dios Caballero, J.; Jones, M.; van Westreenen, M.; Tunney, M.M.; Cantón, R.; Fluit, A.C. Antimicrobial susceptibility of non-fermenting Gram-negative pathogens isolated from cystic fibrosis patients. Int. J. Antimicrob. Agents 2019, 53, 84–88. [Google Scholar] [CrossRef] [Green Version]
- Ko, J.H.; Kang, C.I.; Cornejo-Juárez, P.; Yeh, K.M.; Wang, C.H.; Cho, S.Y.; Gözel, M.G.; Kim, S.H.; Hsueh, P.R.; Sekiya, N.; et al. Fluoroquinolones versus trimethoprim-sulfamethoxazole for the treatment of Stenotrophomonas maltophilia infections: A systematic review and meta-analysis. Clin. Microbiol. Infect. 2019, 25, 546–554. [Google Scholar] [CrossRef]
- Pompilio, A.; Crocetta, V.; De Nicola, S.; Verginelli, F.; Fiscarelli, E.; Di Bonaventura, G. Cooperative pathogenicity in cystic fibrosis: Stenotrophomonas maltophilia modulates Pseudomonas aeruginosa virulence in mixed biofilm. Front. Microbiol. 2015, 6, 951. [Google Scholar] [CrossRef]
- Green, H.; Jones, A. The microbiome and emerging pathogens in cystic fibrosis and non-cystic fibrosis bronchiectasis. Semin. Respir. Crit. Care Med. 2015, 36, 225–235. [Google Scholar] [CrossRef]
- Baldassarri, L.; Creti, R.; Recchia, S.; Imperi, M.; Facinelli, B.; Giovanetti, E.; Pataracchia, M.; Alfarone, G.; Orefici, G. Therapeutic failures of antibiotics used to treat macrolide-susceptible Streptococcus pyogenes infections may be due to biofilm formation. J. Clin. Microbiol. 2006, 44, 2721–2727. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain ID | Isolation Site a | PFGE Type | Susceptibility (S) or Resistance (R) to the Following Antibiotics: | Resistance Phenotype b | Biofilm Formation | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| SXT | CHL | LVX | CPX | TZP | MER | Mean Value (OD492) | Class c | ||||
| SM31 | sputum | 1.1 | S | R | S | S | R | R | MDR | <0.096 | NP |
| SM10 | sputum | 2.1 | S | R | S | R | R | R | XDR | 0.633 | S |
| SM172 | sputum | 3.1 | S | R | S | R | R | R | XDR | 0.432 | S |
| SM176 | sputum | 3.1 | S | R | S | R | S | R | MDR | 0.615 | S |
| SM186 | blood | 3.1 | S | S | S | S | S | S | non-MDR | 0.575 | S |
| SM185 | blood | 3.1 | S | S | R | R | R | R | MDR | 2.282 | S |
| SM143 | sputum | 3.1 | S | R | S | R | R | R | XDR | 2.750 | S |
| SM27 | sputum | 3.1 | S | R | S | R | R | R | XDR | 0.455 | S |
| SM173 | sputum | 4.1 | S | S | S | R | R | R | MDR | 0.574 | S |
| SM139 | CF | 5.1 | S | R | S | R | S | R | MDR | 1.269 | S |
| SM115 | CF | 6.1 | S | S | S | R | R | R | MDR | 0.329 | M |
| SM119 | CF | 7.1 | S | R | S | R | R | R | XDR | 0.236 | M |
| SM108 | CF | 8.1 | R | S | S | R | R | R | XDR | 0.165 | M |
| SM190 | CF | 9.1 | R | R | R | R | R | R | PDR | 0.263 | M |
| SM192 | CF | 9.1 | R | R | R | R | R | R | PDR | 0.446 | S |
| SM193 | CF | 9.1 | R | S | S | R | R | R | XDR | 0.239 | M |
| SM194 | CF | 9.1 | S | S | R | R | R | R | MDR | 0.184 | W |
| SM195 | CF | 9.1 | S | R | R | R | R | R | XDR | 0.195 | M |
| SM134 | CF | 9.1 | S | S | R | R | R | R | XDR | 0.547 | S |
| SM135 | CF | 9.1 | S | S | R | R | R | R | MDR | 0.583 | S |
| SM136 | CF | 9.1 | R | R | R | R | R | R | PDR | 0.541 | S |
| SM106 | CF | 9.1 | S | S | S | R | R | R | MDR | 0.495 | S |
| SM110 | CF | 9.1 | R | R | R | R | R | R | PDR | 0.475 | S |
| SM171 | sputum | 10.1 | S | S | S | R | R | S | non-MDR | 1.099 | S |
| SM123 | CF | 11.1 | S | S | S | R | R | R | MDR | 0.608 | S |
| SM118 | CF | 12.1 | S | S | S | R | S | R | non-MDR | 0.755 | S |
| SM116 | CF | 13.1 | S | R | S | R | R | R | MDR | 0.293 | M |
| SM138 | CF | 14.1 | S | R | S | R | R | R | XDR | 0.391 | S |
| SM109 | CF | 15.1 | S | R | S | S | R | R | MDR | 1.035 | S |
| SM117 | CF | 16.1 | S | R | S | R | R | R | XDR | <0.096 | NP |
| SM113 | CF | 17.1 | R | S | S | R | S | S | non-MDR | <0.096 | NP |
| SM142 | CF | 18.1 | S | R | S | R | R | R | XDR | 1.545 | S |
| SM47 | sputum | 19.1 | S | R | S | S | S | R | non-MDR | 0.100 | W |
| SM157 | CF | 20.1 | S | S | S | S | S | S | non-MDR | <0.096 | NP |
| SM30 | sputum | 21.1 | S | S | S | R | S | R | non-MDR | 0.604 | S |
| SM175 | sputum | 21.1 | S | R | S | R | R | R | XDR | 0.979 | S |
| SM49 | vaginal swab | 22.1 | S | R | S | S | S | S | non-MDR | 0.746 | S |
| SM130 | CF | 23.1 | S | S | S | R | R | R | MDR | 0.546 | S |
| SM184 | blood | 24.1 | S | R | S | R | R | R | XDR | <0.096 | NP |
| SM42 | pharynx swab | 25.1 | S | S | S | R | R | R | MDR | 0.323 | M |
| SM159 | CF | 26.1 | R | S | S | S | R | R | MDR | 0.370 | M |
| SM4 | urine | 27.1 | R | R | R | R | S | R | XDR | 0.732 | S |
| SM8 | urine | 27.1 | R | S | R | R | S | R | MDR | 1.767 | S |
| SM24 | sputum | 27.1 | R | S | R | R | S | R | MDR | 0.629 | S |
| SM37 | blood | 27.1 | R | S | R | R | S | R | MDR | 0.653 | S |
| SM39 | genital swab | 27.1 | R | R | R | R | R | R | PDR | 0.459 | S |
| SM51 | blood | 28.1 | S | R | S | S | S | S | non-MDR | 1.066 | S |
| SM191 | CF | 29.1 | S | R | S | R | R | R | XDR | 0.337 | M |
| SM144 | CF | 31.1 | S | R | S | R | R | R | XDR | 0.590 | S |
| SM174 | sputum | 32.1 | S | S | S | R | R | R | MDR | 0.532 | S |
| SM150 | CF | 33.1 | S | S | S | R | R | R | MDR | 0.391 | S |
| SM180 | blood | 34.1 | S | R | S | R | S | R | MDR | 1.014 | S |
| SM181 | blood | 35.1 | S | R | S | R | R | S | MDR | <0.096 | NP |
| SM137 | CF | 36.1 | S | S | S | R | R | R | MDR | 1.123 | S |
| SM140 | CF | 37.1 | S | R | S | R | R | R | XDR | 0.233 | M |
| SM32 | sputum | 38.1 | S | S | S | S | S | S | non-MDR | 0.509 | S |
| SM21 | rectal swab | 39.1 | S | R | R | R | R | R | XDR | 0.320 | M |
| SM36 | sputum | 40.1 | S | S | S | R | S | S | non-MDR | 3.646 | S |
| SM48 | blood | 40.1 | S | S | S | R | S | R | non-MDR | 0.299 | M |
| SM43 | sputum | 41.1 | S | S | R | S | R | R | MDR | 0.470 | S |
| SM29 | sputum | 42.1 | S | S | S | R | R | R | MDR | 1.720 | S |
| SM50 | sputum | 43.1 | S | R | S | R | R | R | XDR | 2.295 | S |
| SM183 | blood | 44.1 | S | S | S | R | R | R | MDR | 0.397 | S |
| SM107 | CF | 45.1 | S | S | S | R | R | R | MDR | 0.593 | S |
| SM182 | blood | 46.1 | S | S | S | R | S | R | non-MDR | 0.831 | S |
| SM104 | CF | 47.1 | S | R | R | R | R | R | XDR | 0.349 | M |
| SM105 | CF | 48.1 | R | R | R | R | R | R | PDR | 0.581 | S |
| SM120 | CF | 49.1 | R | R | R | R | R | R | PDR | 0.352 | M |
| SM111 | CF | 50.1 | S | S | R | R | R | R | MDR | <0.096 | NP |
| SM112 | CF | 50.1 | S | S | R | R | R | R | MDR | 0.106 | W |
| SM114 | sputum | 51.1 | S | R | S | S | R | R | MDR | <0.096 | NP |
| SM122 | CF | 52.1 | S | S | S | R | R | R | MDR | 1.300 | S |
| SM45 | sputum | 53.1 | S | S | S | R | S | R | non-MDR | 0.596 | S |
| SM170 | sputum | 53.2 | S | R | R | R | R | R | XDR | 0.521 | S |
| SM46 | blood | 54.1 | S | R | S | R | S | R | MDR | 0.888 | S |
| SM5 | sputum | 55.1 | R | R | R | R | R | R | PDR | <0.096 | NP |
| SM177 | sputum | 56.1 | S | R | S | R | R | R | XDR | 0.354 | M |
| SM6 | sputum | 57.1 | S | S | S | S | S | R | non-MDR | 2.517 | S |
| SM38 | sputum | 58.1 | S | S | S | S | S | R | non-MDR | 0.896 | S |
| SM156 | CF | 59.1 | S | S | S | R | R | R | MDR | 0.276 | M |
| SM7 | pharynx swab | 60.1 | S | R | S | S | S | R | non-MDR | 1.111 | S |
| SM124 | CF | 61.1 | S | S | S | S | R | R | non-MDR | <0.096 | NP |
| SM14 | sputum | 62.1 | S | S | S | S | S | S | non-MDR | 0.847 | S |
| SM40 | pharynx swab | 63.1 | S | R | S | R | R | R | XDR | 0.476 | S |
| SM103 | CF | 64.1 | S | S | S | R | R | R | MDR | 0.400 | S |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pompilio, A.; Savini, V.; Fiscarelli, E.; Gherardi, G.; Di Bonaventura, G. Clonal Diversity, Biofilm Formation, and Antimicrobial Resistance among Stenotrophomonas maltophilia Strains from Cystic Fibrosis and Non-Cystic Fibrosis Patients. Antibiotics 2020, 9, 15. https://doi.org/10.3390/antibiotics9010015
Pompilio A, Savini V, Fiscarelli E, Gherardi G, Di Bonaventura G. Clonal Diversity, Biofilm Formation, and Antimicrobial Resistance among Stenotrophomonas maltophilia Strains from Cystic Fibrosis and Non-Cystic Fibrosis Patients. Antibiotics. 2020; 9(1):15. https://doi.org/10.3390/antibiotics9010015
Chicago/Turabian StylePompilio, Arianna, Vincenzo Savini, Ersilia Fiscarelli, Giovanni Gherardi, and Giovanni Di Bonaventura. 2020. "Clonal Diversity, Biofilm Formation, and Antimicrobial Resistance among Stenotrophomonas maltophilia Strains from Cystic Fibrosis and Non-Cystic Fibrosis Patients" Antibiotics 9, no. 1: 15. https://doi.org/10.3390/antibiotics9010015
APA StylePompilio, A., Savini, V., Fiscarelli, E., Gherardi, G., & Di Bonaventura, G. (2020). Clonal Diversity, Biofilm Formation, and Antimicrobial Resistance among Stenotrophomonas maltophilia Strains from Cystic Fibrosis and Non-Cystic Fibrosis Patients. Antibiotics, 9(1), 15. https://doi.org/10.3390/antibiotics9010015