Genetic Profiles and Antimicrobial Resistance Patterns of Salmonella Infantis Strains Isolated in Italy in the Food Chain of Broiler Meat Production

 ,
,  ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Results
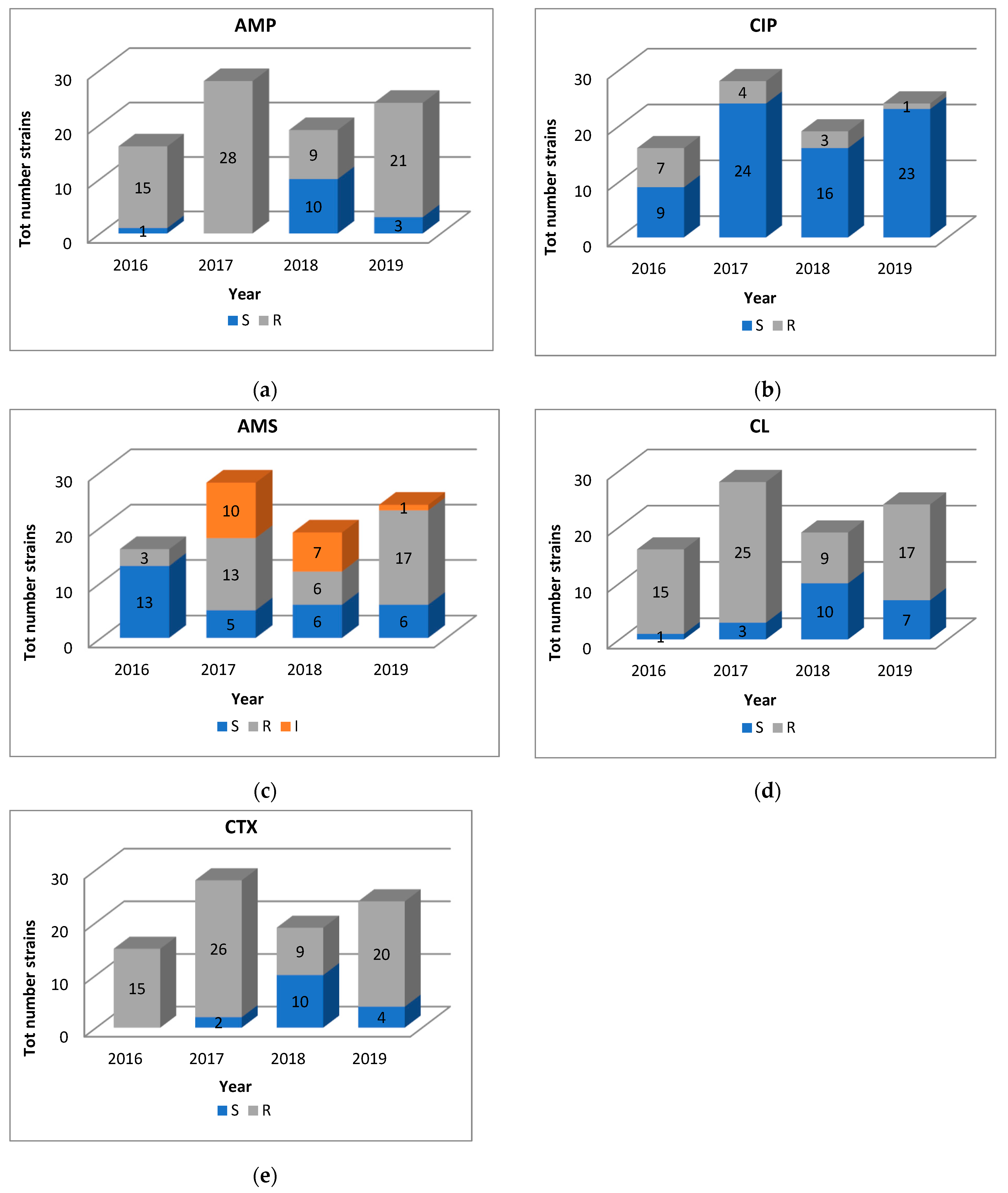
2.1. Antimicrobial Susceptibility and Characterization of ESBL-Encoding and AmpC β-Lactamase Genes
2.2. Macrorestriction PFGE Cluster Analysis
3. Discussion
4. Materials and Methods
4.1. Collection and Identification of Isolates
4.2. Antimicrobial Susceptibility Testing and Detection of ESBL
4.3. Characterization of ESBL-Encoding and AmpC β-Lactamase Genes
4.4. Pulsed Field Gel Electrophoresis (PFGE) Analyses
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- EFSA. The European Union One Health 2018 zoonoses report. EFSA J. 2019, 17, e05926. [Google Scholar]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2017. EFSA J. 2018. Available online: https://www.ecdc.europa.eu/sites/portal/files/documents/zoonoese-food-borne-outbreaks-surveillance-2017-updated.pdf (accessed on 8 October 2020).
- Alba, P.; Leekitcharoenphon, P.; Carfora, V.; Amoruso, R.; Cordaro, G.; Di Matteo, P.; Ianzano, A.; Iurescia, M.; Diaconu, E.L.; Pedersen, S.K.; et al. Molecular epidemiology of salmonella infantis in europe: Insights into the success of the bacterial host and its parasitic pesi-like megaplasmid. Microb. Genom. 2020, 6, e000365. [Google Scholar] [CrossRef]
- Authority, E.F. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 2018, 16. [Google Scholar] [CrossRef]
- Fonseca, E.L.; Mykytczuk, O.L.; Asensi, M.D.; Reis, E.M.F.; Ferraz, L.R.; Paula, F.L.; Ng, L.K.; Rodrigues, D.P. Clonality and antimicrobial resistance gene profiles of multidrug-resistant Salmonella enterica serovar infantis isolates from four public hospitals in Rio de Janeiro, Brazil. J. Clin. Microbiol. 2006, 44, 2767–2772. [Google Scholar] [CrossRef] [Green Version]
- Gal-Mor, O.; Valinsky, L.; Weinberger, M.; Guy, S.; Jaffe, J.; Schorr, Y.I.; Raisfeld, A.; Agmon, V.; Nissan, I. Multidrug resistant salmonella enterica serovar infantis, Israel. Emerg. Infect. Dis. 2010, 16, 1754–1757. [Google Scholar] [CrossRef]
- Hindermann, D.; Gopinath, G.; Chase, H.; Negrete, F.; Althaus, D.; Zurfluh, K.; Tall, B.D.; Stephan, R.; Nüesch-Inderbinen, M. Salmonella enterica serovar infantis from food and human infections, Switzerland, 2010–2015: Poultry-related multidrug resistant clones and an emerging ESBL producing clonal lineage. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Pate, M.; Mičunovič, J.; Golob, M.; Vestby, L.K.; Ocepek, M. Salmonella Infantis in Broiler Flocks in Slovenia: The Prevalence of Multidrug Resistant Strains with High Genetic Homogeneity and Low Biofilm-Forming Ability. Biomed. Res. Int. 2019, 2019. [Google Scholar] [CrossRef] [Green Version]
- Nógrády, N.; Király, M.; Davies, R.; Nagy, B. Multidrug resistant clones of Salmonella Infantis of broiler origin in Europe. Int. J. Food Microbiol. 2012, 157, 108–112. [Google Scholar] [CrossRef]
- García-Soto, S.; Abdel-Glil, M.Y.; Tomaso, H.; Linde, J.; Methner, U. Emergence of Multidrug-Resistant Salmonella enterica Subspecies enterica Serovar Infantis of Multilocus Sequence Type 2283 in German Broiler Farms. Front. Microbiol. 2020, 11, 1741. [Google Scholar] [CrossRef]
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Di Giannatale, E.; et al. Emergence of a Clonal Lineage of Multidrug-Resistant ESBL-Producing Salmonella Infantis Transmitted from Broilers and Broiler Meat to Humans in Italy between 2011 and 2014. PLoS ONE 2015, 10, e0144802. [Google Scholar] [CrossRef] [Green Version]
- Dionisi, A.M.; Lucarelli, C.; Benedetti, I.; Owczarek, S.; Luzzi, I. Molecular characterisation of multidrug-resistant Salmonella enterica serotype Infantis from humans, animals and the environment in Italy. Int. J. Antimicrob. Agents 2011, 38, 384–389. [Google Scholar] [CrossRef]
- Helms, M.; Vastrup, P.; Gerner-Smidt, P.; Mølbak, K. Excess mortality associated with antimicrobial drug-resistant Salmonella typhimurium. Emerg. Infect. Dis. 2002, 8, 490–495. [Google Scholar] [CrossRef]
- EFSA; ECDC. The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-Borne Outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar]
- Collignon, P.C.; Conly, J.M.; Andremont, A.; McEwen, S.A.; Aidara-Kane, A.; Griffin, P.M.; Agerso, Y.; Dang Ninh, T.; Donado-Godoy, P.; Fedorka-Cray, P.; et al. World Health Organization Ranking of Antimicrobials According to Their Importance in Human Medicine: A Critical Step for Developing Risk Management Strategies to Control Antimicrobial Resistance from Food Animal Production. In Proceedings of the Clinical Infectious Diseases; Oxford University Press: Oxford, UK, 2016; Volume 63, pp. 1087–1093. [Google Scholar]
- Aviv, G.; Tsyba, K.; Steck, N.; Salmon-Divon, M.; Cornelius, A.; Rahav, G.; Grassl, G.A.; Gal-Mor, O. A unique megaplasmid contributes to stress tolerance and pathogenicity of an emergent Salmonella enterica serovar Infantis strain. Environ. Microbiol. 2014, 16, 977–994. [Google Scholar] [CrossRef]
- Carfora, V.; Alba, P.; Leekitcharoenphon, P.; Ballarò, D.; Cordaro, G.; Di Matteo, P.; Donati, V.; Ianzano, A.; Iurescia, M.; Stravino, F.; et al. Colistin resistance mediated by mcr-1 in ESBL-producing, multidrug resistant Salmonella infantis in broiler chicken industry, Italy (2016–2017). Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef]
- Koutsoumanis, K.; Allende, A.; Alvarez-Ordóñez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; De Cesare, A.; Herman, L.; Hilbert, F.; Lindqvist, R.; et al. Salmonella control in poultry flocks and its public health impact. EFSA J. 2019, 17, e05596. [Google Scholar] [CrossRef]
- Musa, L.; Casagrande Proietti, P.; Branciari, R.; Menchetti, L.; Bellucci, S.; Ranucci, D.; Marenzoni, M.L.; Franciosini, M.P. Antimicrobial Susceptibility of Escherichia coli and ESBL-Producing Escherichia coli Diffusion in Conventional, Organic and Antibiotic-Free Meat Chickens at Slaughter. Animals 2020, 10, 1215. [Google Scholar] [CrossRef]
- Newton, K.; Gosling, B.; Rabie, A.; Davies, R. Field investigations of multidrug resistant Salmonella Infantis epidemic strain incursions into broiler flocks in England and Wales. Avian Pathol. 2020, 1–28. [Google Scholar] [CrossRef]
- Wassenaar, T.M. Use of antimicrobial agents in veterinary medicine and implications for human health. Crit. Rev. Microbiol. 2005, 31, 155–169. [Google Scholar] [CrossRef]
- Roschanski, N.; Fischer, J.; Falgenhauer, L.; Pietsch, M.; Guenther, S.; Kreienbrock, L.; Chakraborty, T.; Pfeifer, Y.; Guerra, B.; Roesler, U.H. Retrospective analysis of bacterial cultures sampled in German chicken-fattening farms during the years 2011-2012 revealed additional VIM-1 carbapenemase-producing Escherichia coli and a serologically rough Salmonella enterica serovar infantis. Front. Microbiol. 2018, 9, 538. [Google Scholar] [CrossRef] [PubMed]
- Thakur, Y.R.; Bajaj, B.K. Antibiotic resistance and molecular characterization of poultry isolates of Salmonella by RAPD-PCR. World J. Microbiol. Biotechnol. 2006, 22, 1177–1183. [Google Scholar] [CrossRef]
- Boy-Roura, M.; Mas-Pla, J.; Petrovic, M.; Gros, M.; Soler, D.; Brusi, D.; Menció, A. Towards the understanding of antibiotic occurrence and transport in groundwater: Findings from the Baix Fluvià alluvial aquifer (NE Catalonia, Spain). Sci. Total Environ. 2018, 612, 1387–1406. [Google Scholar] [CrossRef] [PubMed]
- Wright, G.D. Antibiotic resistance in the environment: A link to the clinic? Curr. Opin. Microbiol. 2010, 13, 589–594. [Google Scholar] [CrossRef] [PubMed]
- Bonardi, S.; Pitino, R. Carbapenemase-producing bacteria in food-producing animals, wildlife and environment: A challenge for human health. Ital. J. Food Saf. 2019, 8, 7956. [Google Scholar] [CrossRef]
- Fu, Y.; Xu, X.; Zhang, L.; Xiong, Z.; Ma, Y.; Wei, Y.; Chen, Z.; Bai, J.; Liao, M.; Zhang, J. Fourth Generation Cephalosporin Resistance Among Salmonella enterica Serovar Enteritidis Isolates in Shanghai, China Conferred by blaCTX–M–55 Harboring Plasmids. Front. Microbiol. 2020, 11, 910. [Google Scholar] [CrossRef]
- European Medicine Agency. Available online: https://www.ema.europa.eu/en/documents/press-release/categorisation-antibiotics-used-animals-promotes-responsible-use-protect-public-animal-health_en.pdf (accessed on 19 October 2020).
- Andersson, D.I.; Hughes, D. Evolution of antibiotic resistance at non-lethal drug concentrations. Drug Resist. Updates 2012, 15, 162–172. [Google Scholar] [CrossRef]
- Dionisi, A.M.; Owczarek, S.; Benedetti, I.; Luzzi, I.; García-Fernández, A. Extended-spectrum β-lactamase-producing Salmonella enterica serovar Infantis from humans in Italy. Int. J. Antimicrob. Agents 2016, 48, 345–346. [Google Scholar] [CrossRef]
- ISO. ISO 6579-1:2017/Amd 1:2020—Microbiology of the Food Chain—Horizontal Method for the Detection, Enumeration and Serotyping of Salmonella—Part 1: Detection of Salmonella spp.—Amendment 1: Broader Range of Incubation Temperatures, Amendment to the Status of Annex D, and Correction of the Composition of MSRV and SC. Available online: https://www.iso.org/standard/76671.html (accessed on 17 September 2020).
- Grimont, P.A.D.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Center for Reference and Research on Salmonella, Institut Pasteur: Paris, France, 2007; Available online: http://www.scacm.org/free/Antigenic%20Formulae%20of%20the%20Salmonella%20Serovars%202007%209th%20edition.pdf (accessed on 17 September 2020).
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; CLSI Supplement M100; CLSI: Wayne, PA, USA, 2020. [Google Scholar]
- European Committee on Antimicrobial Susceptibility Testing. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/Breakpoint_tables/v_10.0_Breakpoint_Tables.pdf (accessed on 2 October 2020).
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [Green Version]
- Woodford, N.; Fagan, E.J.; Ellington, M.J. Multiplex PCR for rapid detection of genes encoding CTX-M extended-spectrum β-lactamases [4]. J. Antimicrob. Chemother. 2006, 57, 154–155. [Google Scholar] [CrossRef] [Green Version]
- Heider, L.C.; Hoet, A.E.; Wittum, T.E.; Khaitsa, M.L.; Love, B.C.; Huston, C.L.; Morley, P.S.; Funk, J.A.; Gebreyes, W.A. Genetic and phenotypic characterization of the blaCMY gene from escherichia coli and salmonella enterica isolated from food-producing animals, humans, the environment, and retail meat. Foodborne Pathog. Dis. 2009, 6, 1235–1240. [Google Scholar] [CrossRef] [PubMed]
- Ribot, E.M.; Fair, M.A.; Gautom, R.; Cameron, D.N.; Hunter, S.B.; Swaminathan, B.; Barrett, T.J. Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 2006, 3, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Menchetti, L.; Padalino, B.; Brasileiro Fernandes, F.; Nanni Costa, L. Comparison of nonlinear growth models and factors affecting body weight at different ages in Toy Poodles. Ital. J. Anim. Sci. 2020, 19, 792–802. [Google Scholar] [CrossRef]
- Peduzzi, P.; Concato, J.; Feinstein, A.R.; Holford, T.R. Importance of events per independent variable in proportional hazards regression analysis II. Accuracy and precision of regression estimates. J. Clin. Epidemiol. 1995, 48, 1503–1510. [Google Scholar] [CrossRef] [PubMed]
- Watson, P.F.; Petrie, A. Method agreement analysis: A review of correct methodology. Theriogenology 2010, 73, 1167–1179. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Antimicrobials | Number of Isolates (%) | ||
|---|---|---|---|
| R | I | S | |
| Ampicillin-AMP | 73 (84) | 0 (-) | 14 (16) |
| Cefalexin-CL | 66 (76) | 0 (-) | 21 (24) |
| Cefoxitin-FOX | 0 (-) | 22 (25) | 65 (75) |
| Cefotaxime-CTX | 70 (80.5) | 0 (-) | 17 (19.5) |
| Ceftazidime-CAZ | 10 (11.5) | 30 (34.5) | 47(54) |
| Cefepime-FEP | 63 (72.4) | 12 (13.8) | 12 (13.8) |
| Nalidixic acid-NA | 85 (97.7) | 2 (2.3) | 0 (-) |
| Ciprofloxacin-CIP | 15 (18) | 49 (56.3) | 23(26.4) |
| Tetracycline-TE | 84 (96.5) | 3 (3.5) | 0 (-) |
| Trimethoprim/sulphametoxazole-SUL | 79 (91) | 1 (1) | 7 (8) |
| Cloramphenicols-CLOR | 18 (20.7) | 20 (23) | 49 (56.3) |
| Gentamicin-GENT | 14 (16) | 0 (-) | 73 (84) |
| Imipenem-IMP | 0 (-) | 9 (10.3) | 78 (89.7) |
| Meropenem-MEM | 5 (5.7) | 10 (11.5) | 72 (82.8) |
| Ampicillin/sulbactam-AMS | 39 (44.8) | 18 (20.7) | 30 (34.5) |
| Amoxicillin/clavulanic acid-AMC | 14 (16) | 15 (17.3) | 58 (66.7) |
| No of Resistance Antibiotic Classes | Antimicrobial Resistance Pattern | N° of Isolates | % of Isolates | blaCTX-M1 | blaCMY-2 |
|---|---|---|---|---|---|
| 1 | beta | 1 | 1.1% | 0 | 0 |
| 2 | quin/tetra | 1 | 1.1% | 0 | 0 |
| 3 | beta/quin/tetra | 3 | 3.4% | 2 | 2 |
| quin/sulph/tetra | 6 | 6.9% | 0 | 0 | |
| amino/sulph/tetra | 1 | 1.1% | 0 | 0 | |
| 4 | beta/amino/quin/tetra | 4 | 4.6% | 3 | 0 |
| beta/quin/sulph/tetra | 45 | 51.7% | 40 | 0 | |
| beta/clor/sulph/tetra beta/quin/clor/sulph | 1 1 | 1.1% 1.1% | 1 1 | 0 | |
| 5 | beta/amino/quin/sulph/tetra | 6 | 6.9% | 6 | 0 |
| beta/carba/quin/sulph/tetra | 5 | 5.8% | 4 | 0 | |
| beta/quin/clor/sulph/tetra | 8 | 9.2% | 8 | 0 | |
| 6 | beta/amino/carba/quin/sulph/tetra | 1 | 1.1% | 1 | 0 |
| beta/amino/quin/clor/sulph/tetra | 4 | 4.6% | 4 | 0 |
| No of Resistance Antibiotic Classes | Antimicrobial Resistance Pattern | N° of Isolates | % of Isolates |
|---|---|---|---|
| 1 | beta | 1 | 5.9% |
| 2 | quin/tetra | 1 | 5.9% |
| 3 | beta/quin/tetra | 1 | 5.9% |
| quin/sulph/tetra | 8 | 47% | |
| amino/sulph/tetra | 1 | 5.9% | |
| amino/quin/tetra | 1 | 5.9% | |
| beta/quin/sulph/tetra | 3 | 17.6% | |
| carba/quin/sulph/tetra | 1 | 5.9% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proietti, P.C.; Stefanetti, V.; Musa, L.; Zicavo, A.; Dionisi, A.M.; Bellucci, S.; Mensa, A.L.; Menchetti, L.; Branciari, R.; Ortenzi, R.; et al. Genetic Profiles and Antimicrobial Resistance Patterns of Salmonella Infantis Strains Isolated in Italy in the Food Chain of Broiler Meat Production. Antibiotics 2020, 9, 814. https://doi.org/10.3390/antibiotics9110814
Proietti PC, Stefanetti V, Musa L, Zicavo A, Dionisi AM, Bellucci S, Mensa AL, Menchetti L, Branciari R, Ortenzi R, et al. Genetic Profiles and Antimicrobial Resistance Patterns of Salmonella Infantis Strains Isolated in Italy in the Food Chain of Broiler Meat Production. Antibiotics. 2020; 9(11):814. https://doi.org/10.3390/antibiotics9110814
Chicago/Turabian StyleProietti, Patrizia Casagrande, Valentina Stefanetti, Laura Musa, Alessia Zicavo, Anna Maria Dionisi, Sara Bellucci, Agnese La Mensa, Laura Menchetti, Raffaella Branciari, Roberta Ortenzi, and et al. 2020. "Genetic Profiles and Antimicrobial Resistance Patterns of Salmonella Infantis Strains Isolated in Italy in the Food Chain of Broiler Meat Production" Antibiotics 9, no. 11: 814. https://doi.org/10.3390/antibiotics9110814
APA StyleProietti, P. C., Stefanetti, V., Musa, L., Zicavo, A., Dionisi, A. M., Bellucci, S., Mensa, A. L., Menchetti, L., Branciari, R., Ortenzi, R., & Franciosini, M. P. (2020). Genetic Profiles and Antimicrobial Resistance Patterns of Salmonella Infantis Strains Isolated in Italy in the Food Chain of Broiler Meat Production. Antibiotics, 9(11), 814. https://doi.org/10.3390/antibiotics9110814