Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union

 ,
,  ,
,  and
and 
Abstract
:1. Introduction
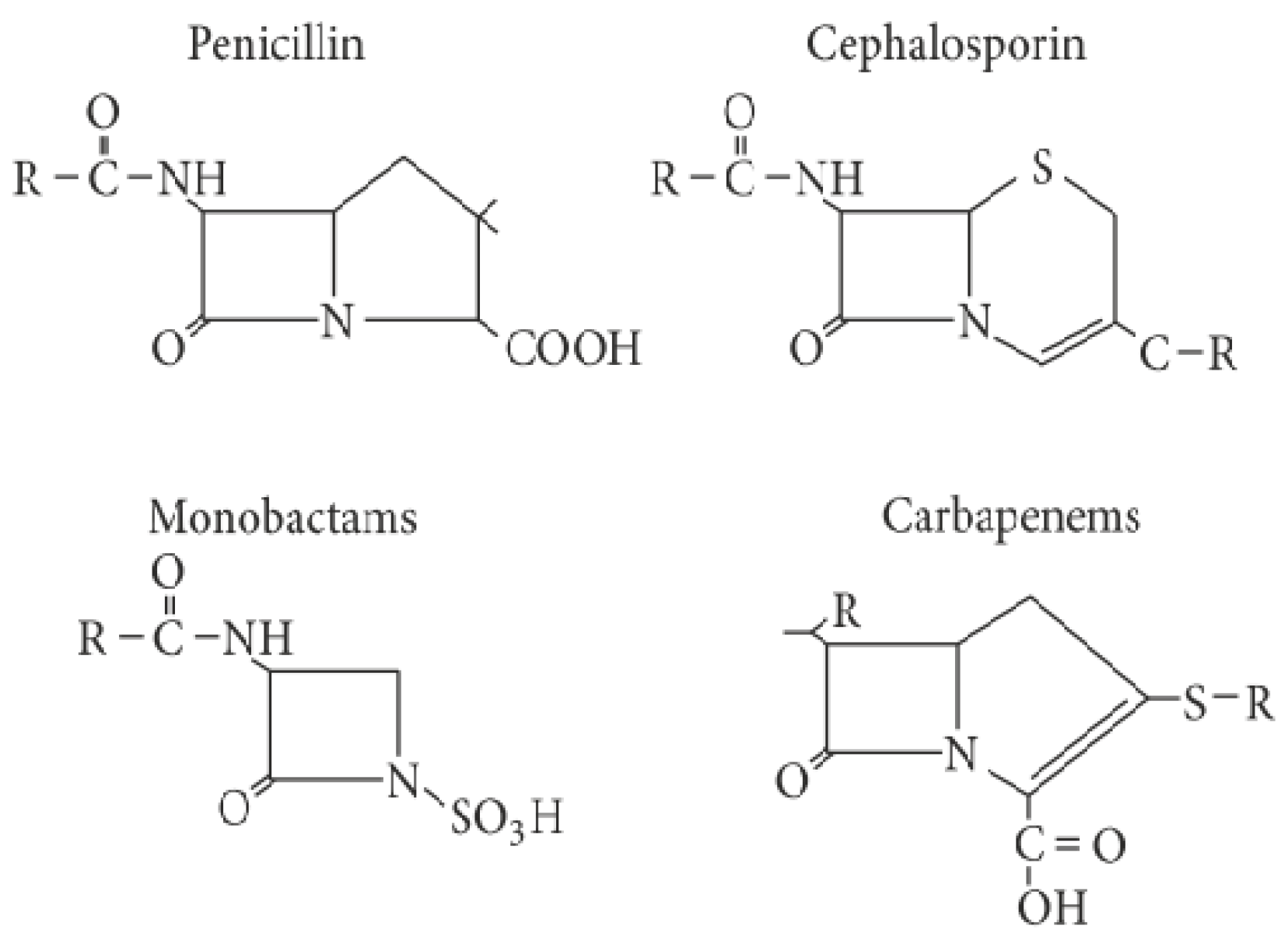
2. Classification of ESBLs
3. Dissemination of ESBLs
3.1. Dissemination Pathways of Antibiotic Resistance Genes
3.2. Dissemination of ESBL-Producing E. coli in the Pork Production Chain
4. Use of β-Lactam Antibiotics in Pig Production
4.1. Usage of β-lactam Antibiotics and Emergence of Related AMR Determinants in Pig Production
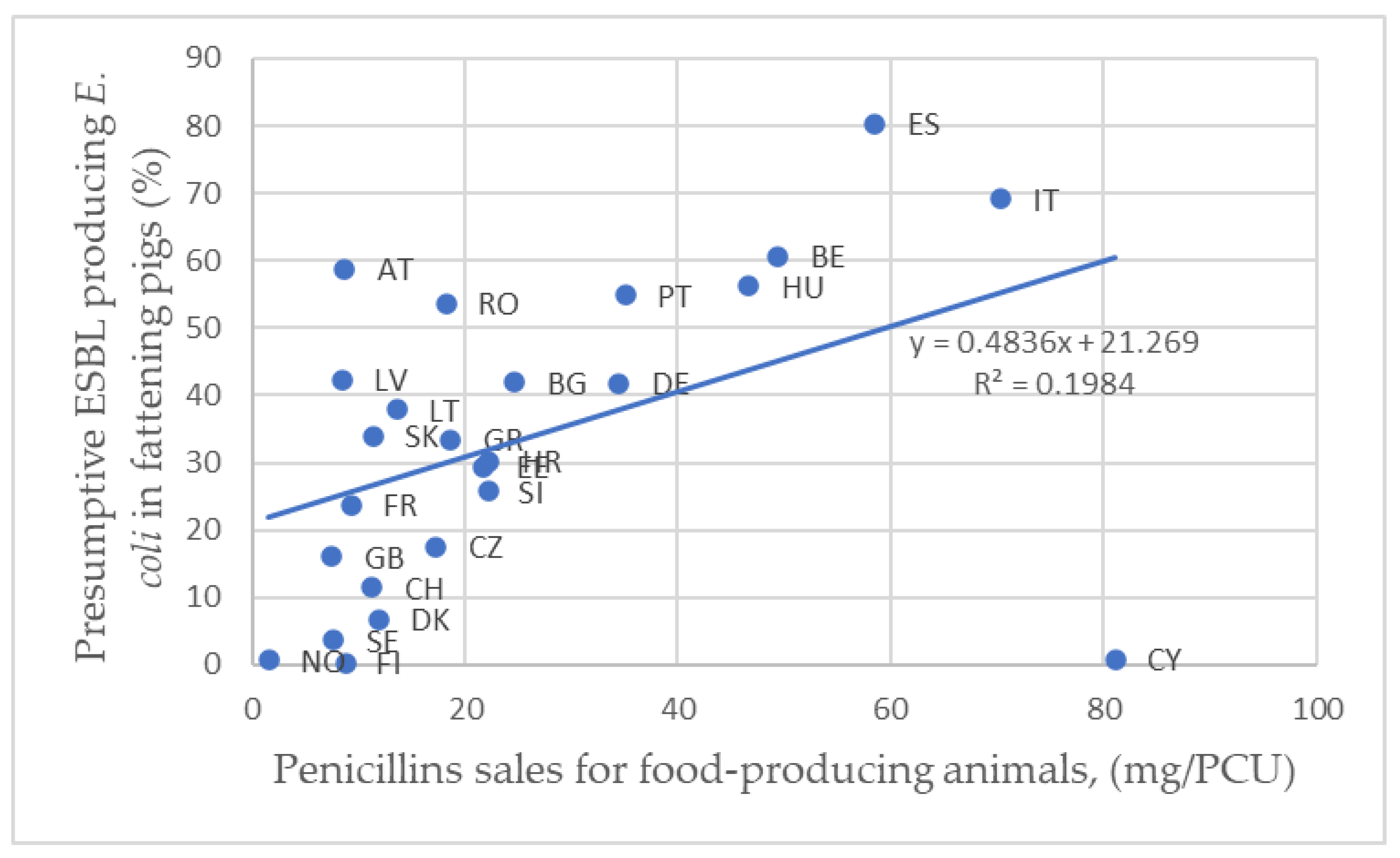
4.2. Sales of Penicillins and Cephalosporins in the EU
5. ESBL Producing E. coli in the Pork Production Chain
5.1. Prevalence of ESBL Producing E. coli in Fattening Pigs
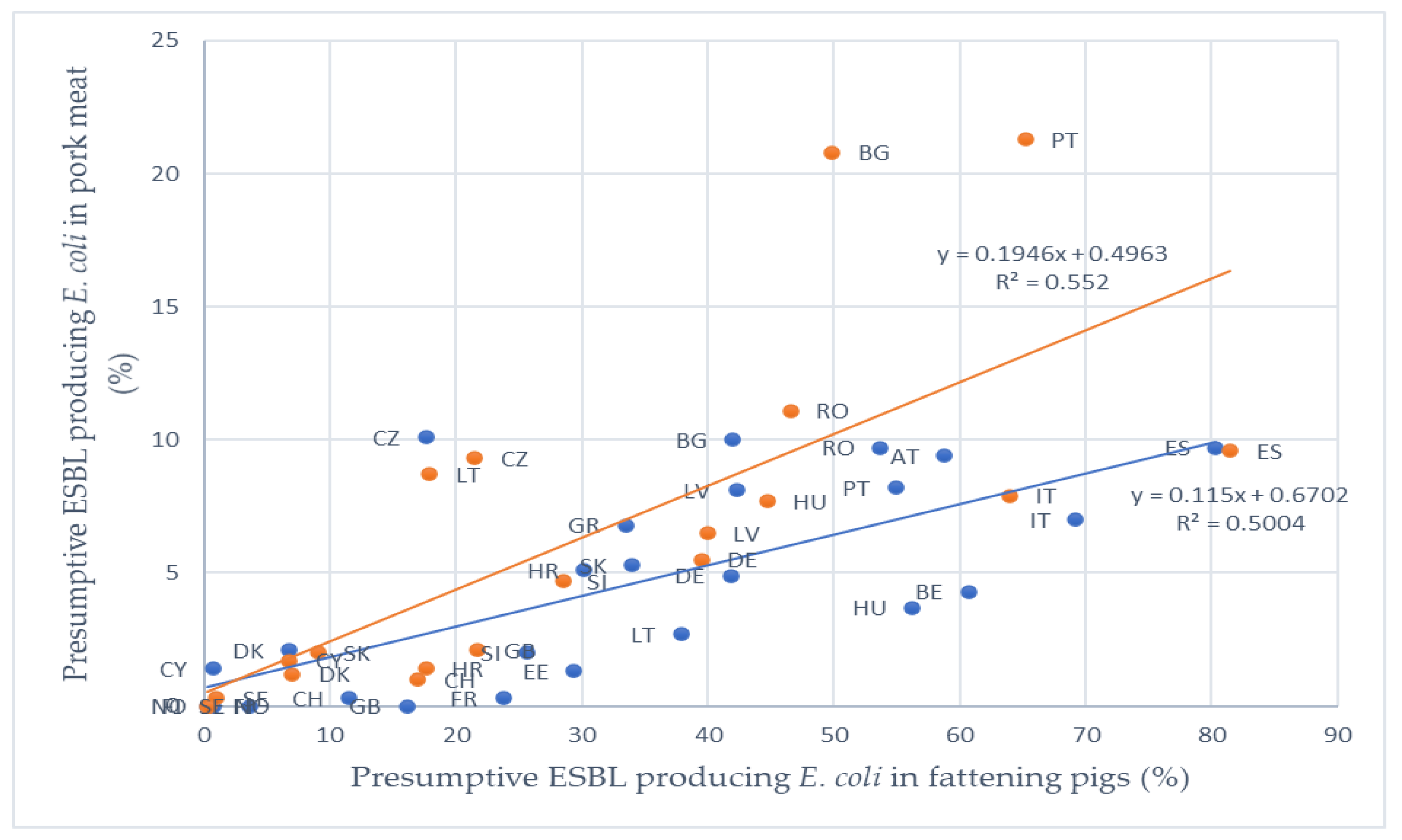
5.2. Prevalence of ESBL Producing E. coli in Pork Meat
6. Literature Review on the Prevalence of ESBL Producing E. coli in Pork Meat
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- WHO Antimicrobial Resistance. Available online: https://www.who.int/news-room/fact-sheets/detail/antimicrobial-resistance (accessed on 22 June 2020).
- Williams, K.J. The introduction of “chemotherapy” using arsphenamine—The first magic bullet. J. R. Soc. Med. 2009, 102, 343–348. [Google Scholar] [CrossRef] [Green Version]
- Livermore, D.M.; Woodford, N. The β-lactamase threat in Enterobacteriaceae, Pseudomonas and Acinetobacter. Trends Microbiol. 2006, 14, 413–420. [Google Scholar] [CrossRef] [PubMed]
- Peirano, G.; Pitout, J.D.D. Extended-Spectrum β-Lactamase-Producing Enterobacteriaceae: Update on Molecular Epidemiology and Treatment Options. Drugs 2019, 79, 1529–1541. [Google Scholar] [CrossRef] [PubMed]
- World Health Organisation. Prioritization of Pathogens to Guide Discovery, Research and Development of New Antibiotics for Drug-Resistant Bacterial Infections, including Tuberculosis; World Health Organisation: Geneva, Switzerland, 2017; pp. 1–88. [Google Scholar]
- Smet, A.; Martel, A.; Persoons, D.; Dewulf, J.; Heyndrickx, M.; Herman, L.; Haesebrouck, F.; Butaye, P. Broad-spectrum β-lactamases among Enterobacteriaceae of animal origin: Molecular aspects, mobility and impact on public health. FEMS Microbiol. Rev. 2010, 34, 295–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, E.P.; Chain, E. An Enzyme from Bacteria able to Destroy Penicillin. Nature 1940, 3713, 837. [Google Scholar] [CrossRef]
- Pitout, J.D.D.; Laupland, K.B. Extended-spectrum β-lactamase-producing Enterobacteriaceae: An emerging public-health concern. Lancet Infect. Dis. 2010, 8, 159–166. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Webber, M.A.; Baylay, A.J.; Ogbolu, D.O.; Piddock, L.J.V. Molecular mechanisms of antibiotic resistance. Nat. Rev. Microbiol. 2015, 13, 42–51. [Google Scholar] [CrossRef] [PubMed]
- Bradford, P.A. Extended-Spectrum-Lactamases in the 21st Century: Characterization, Epidemiology, and Detection of This Important Resistance Threat. Clin. Microbiol. Rev. 2001, 14, 933–951. [Google Scholar] [CrossRef] [Green Version]
- Bush, K.; Bradford, P.A. Epidemiology of β-lactamase-producing pathogens. Clin. Microbiol. Rev. 2020, 33, 1–37. [Google Scholar] [CrossRef]
- Ur Rahman, S.; Ali, T.; Ali, I.; Khan, N.A.; Han, B.; Gao, J. The Growing Genetic and Functional Diversity of Extended Spectrum Beta-Lactamases. Biomed. Res. Int. 2018, 2018. [Google Scholar] [CrossRef]
- Medeiros, A.A. Evolution and Dissemination of β-Lactamases Accelerated by Generations of β-Lactam Antibiotics. Clin. Infect. Dis. 1997, 24, 19–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, N.; Kontomichalou, P. Penicillinase synthesis controlled by infectious R factors in Enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef] [PubMed]
- Kliebe, C.; Nies, B.A.; Meyer, J.F.; Tolxdorff-Neutzling, R.M.; Wiedemann, B. Evolution of plasmid-coded resistance to broad-spectrum cephalosporins. Antimicrob. Agents Chemother. 1985, 28, 302–307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirot, D.; Sirot, J.; Labia, R.; Morand, A.; Courvalin, P.; Darfeuille-michaud, A.; Perroux, R.; Cluzel, R. Transferable resistance to third-generation cephalosporins in clinical isolates of klebsiella pneumoniae: Identification of CTX-1, a novel β-lactamase. J. Antimicrob. Chemother. 1987, 20, 323–334. [Google Scholar] [CrossRef]
- Irrgang, A.; Hammerl, J.A.; Falgenhauer, L.; Guiral, E.; Schmoger, S.; Imirzalioglu, C.; Fischer, J.; Guerra, B.; Chakraborty, T.; Käsbohrer, A. Diversity of CTX-M-1-producing E. coli from German food samples and genetic diversity of the blaCTX-M-1 region on IncI1 ST3 plasmids. Vet. Microbiol. 2018, 221, 98–104. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2015. EFSA J. 2017, 15, e04694. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2017. EFSA J. 2019, 17, 5598. [Google Scholar] [CrossRef]
- Schmithausen, R.M.; Schulze-Geisthoevel, S.V.; Heinemann, C.; Bierbaum, G.; Exner, M.; Petersen, B.; Steinhoff-Wagner, J. Reservoirs and transmission pathways of resistant indicator bacteria in the Biotope pig stable and along the food chain: A review froma One Health Perspective. Sustainability 2018, 10, 3967. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Dalsgaard, A.; Vieira, A.R.; Emborg, H.D.; Jensen, L.B. Prevalence of tetracycline resistance and genotypic analysis of populations of Escherichia coli from animals, carcasses and cuts processed at a pig slaughterhouse. Int. J. Food Microbiol. 2009, 135, 254–259. [Google Scholar] [CrossRef]
- Warriner, K.; Aldsworth, T.G.; Kaur, S.; Dodd, C.E.R. Cross-contamination of carcasses and equipment during pork processing. J. Appl. Microbiol. 2002, 93, 169–177. [Google Scholar] [CrossRef]
- Ambler, R.P. The structure of β-lactamases. Phil. Trans. R. Soc. L. 1980, 289, 321–331. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A.; Medeiros, A.A. A functional classification scheme for β-lactamases and its correlation with molecular structure. Antimicrob. Agents Chemother. 1995, 39, 1211–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schill, F.; Abdulmawjood, A.; Klein, G.; Reich, F. Prevalence and characterization of extended-spectrum β-lactamase (ESBL) and AmpC β-lactamase producing Enterobacteriaceae in fresh pork meat at processing level in Germany. Int. J. Food Microbiol. 2017, 257, 58–66. [Google Scholar] [CrossRef] [PubMed]
- Palzkill, T. Metallo-β-lactamase structure and function. Ann. N. Y. Acad. Sci. 2013, 1277, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R. Epidemiology and Evolution of Beta-Lactamases. Evol. Biol. Bact. Fungal Pathog. 2008, 249–270. [Google Scholar] [CrossRef]
- Vahaboglu, H.; Öztürk, R.; Aygün, G.; Coşkunkan, F.; Yaman, A.; Kaygusuz, A.; Leblebicioglu, H.; Balik, I.; Aydin, K.; Otkun, M. Widespread detection of per-1-type extended-spectrum β-lactamases among nosocomial Acinetobacter and Pseudomonas aeruginosa isolates in Turkey: A nationwide multicenter study. Antimicrob. Agents Chemother. 1997, 41, 2265–2269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, Y.; Ikeda, F.; Kamimura, T.; Yokota, Y.; Mine, Y. Novel plasmid-mediated β-lactamase from Escherichia coli that inactivates oxyimino-cephalosporins. Antimicrob. Agents Chemother. 1988, 32, 1243–1246. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Inoue, M. Characterization of SFO-1, a plasmid-mediated inducible class A β- lactamase from Enterobacter cloacae. Antimicrob. Agents Chemother. 1999, 43, 307–313. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Le Thomas, I.; Naas, T.; Karim, A.; Nordmann, P. Biochemical sequence analyses of GES-1, a novel class A extended- spectrum β-lactamase, and the class 1 integron In52 from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 2000, 44, 622–632. [Google Scholar] [CrossRef] [Green Version]
- Rossolini, G.M.; Franceschini, N.; Lauretti, L.; Caravelli, B.; Riccio, M.L.; Galleni, M.; Frère, J.M.; Amicosante, G. Cloning of a Chryseobacterium (Flavobacterium) meningosepticum chromosomal gene (blaA(CME)) encoding an extended-spectrum class a β- lactamase related to the Bacteroides cephalosporinases and the VEB-1 and PER β-lactamases. Antimicrob. Agents Chemother. 1999, 43, 2193–2199. [Google Scholar] [CrossRef] [Green Version]
- Nordmann, P.; Ronco, E.; Naas, T.; Duport, C.; Michel-Briand, Y.; Labia, R. Characterization of a novel extended-spectrum β-lactamase from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1993, 37, 962–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonnet, R.; Sampaio, J.L.M.; Chanal, C.; Sirot, D.; De Champs, C.; Viallard, J.L.; Labia, R.; Sirot, J. A novel class A extended-spectrum β-lactamase (BES-1) in Serratia marcescens isolated in Brazil. Antimicrob. Agents Chemother. 2000, 44, 3061–3068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, J.; Aguilar, C.; Ayala, G.; Estrada, M.A.; Garza-Ramos, A.; Lara-Lemus, R.; Ledezma, L. TLA-1 : A New Plasmid-Mediated Extended-Spectrum β-Lactamase from Escherichia coli. Antimicrob. Agents Chemother. 2000, 44, 997–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danel, F.; Hall, L.M.C.; Gur, D.; Livermore, D.M. OXA-14, another extended-spectrum variant of OXA-10 (PSE-2) β-lactamase from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1995, 39, 1881–1884. [Google Scholar] [CrossRef] [Green Version]
- Danel, F.; Hall, L.M.C.; Gur, D.; Livermore, D.M. OXA-16, a further extended-spectrum variant of OXA-10 β-lactamase, from two Pseudomonas aeruginosa isolates. Antimicrob. Agents Chemother. 1998, 42, 3117–3122. [Google Scholar] [CrossRef] [Green Version]
- Danel, F.; Hall, L.M.C.; Duke, B.; Gur, D.; Livermore, D.M. OXA-17, a further extended-spectrum variant of OXA-10 β-lactamase, isolated from Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 1999, 43, 1362–1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mugnier, P.; Casin, I.; Bouthors, A.T.; Collatz, E. Novel OXA-10-derived extended-spectrum β-lactamases selected in vivo or in vitro. Antimicrob. Agents Chemother. 1998, 42, 3113–3116. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Girlich, D.; Naas, T.; Nordmann, P. OXA-28, an extended-spectrum variant of OXA-10 β-lactamase from Pseudomonas aeruginosa and its plasmid- and integron-located gene. Antimicrob. Agents Chemother. 2001, 45, 447–453. [Google Scholar] [CrossRef] [Green Version]
- Danel, F.; Hall, L.M.C.; Gur, D.; Livermore, D.M. OXA-15, an extended-spectrum variant of OXA-2 β-lactamase, isolated from a Pseudomonas aeruginosa strain. Antimicrob. Agents Chemother. 1997, 41, 785–790. [Google Scholar] [CrossRef] [Green Version]
- Evans, B.A.; Amyes, S.G.B. OXA β-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef] [Green Version]
- D’Andrea, M.M.; Arena, F.; Pallecchi, L.; Rossolini, G.M. CTX-M-type β-lactamases: A successful story of antibiotic resistance. Int. J. Med. Microbiol. 2013, 303, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Bush, K.; Macielag, M.J. New β-lactam antibiotics and β-lactamase inhibitors. Expert Opin. Ther. Patents 2010, 20, 1277–1293. [Google Scholar] [CrossRef] [PubMed]
- Pleiss, J. The Lactamase Engineering Database. Available online: http://www.laced.uni-stuttgart.de/ (accessed on 22 June 2020).
- Clasen, J.; Birkegård, A.C.; Græsbøll, K.; Folkesson, A. Evolution of TEM-type extended-spectrum β-lactamases in Escherichia coli by cephalosporins. J. Glob. Antimicrob. Resist. 2019, 19, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A review of SHV extended-spectrum β-lactamases: Neglected yet ubiquitous. Front. Microbiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Poirel, L.; He, C.; Podglajen, I.; Sougakoff, W.; Gutmann, L.; Nordmann, P. Emergence in Klebsiella pneumoniae of a Chromosome-Encoded SHV β-Lactamase That Compromises the Efficacy of Imipenem. Antimi 2003, 47, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Tzouvelekis, L.; Bonomo, R.A. SHV-type beta-lactamases. Curr Pharm Des. 1999, 5, 847–864. [Google Scholar]
- Pitton, J.S. Mechanisms of Bacterial resistance to antibiotics. Rev. Physiol. 1972, 15–92. [Google Scholar]
- Bauernfeind, A.; Stemplinger, I.; Jungwirth, R.; Ernst, S.; Casellas, J.M. Sequences of β-lactamase genes encoding CTX-M-1 (MEN-1) and CTX-M-2 and relationship of their amino acid sequences with those of other β-lactamases. Antimicrob. Agents Chemother. 1996, 40, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Gniadkowski, M.; Schneider, I.; Pałucha, A.; Jungwirth, R.; Mikiewicz, B.; Bauernfeind, A. Cefotaxime-resistant Enterobacteriaceae isolates from a hospital in Warsaw, Poland: Identification of a new CTX-M-3 cefotaxime-hydrolyzing β- lactamase that is closely related to the CTX-M-1/MEN-1 enzyme. Antimicrob. Agents Chemother. 1998, 42, 827–832. [Google Scholar] [CrossRef] [Green Version]
- Bonnet, R.; Sampaio, J.L.M.; Labia, R.; De Champs, C.; Sirot, D.; Chanal, C.; Sirot, J. A novel CTX-M β-lactamase (CTX-M-8) in cefotaxime-resistant Enterobacteriaceae isolated in Brazil. Antimicrob. Agents Chemother. 2000, 44, 1936–1942. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 2009, 53, 2227–2238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, A.B.; Silverman, M.; Boyd, D.A.; McGeer, A.; Willey, B.M.; Pong-Porter, V.; Daneman, N.; Mulvey, M.R. Identification of a progenitor of the CTX-M-9 group of extended-spectrum β-lactamases from Kluyvera georgiana isolated in Guyana. Antimicrob. Agents Chemother. 2005, 49, 2112–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Lagacé-Wiens, P.R.S.; Tailor, F.; Simner, P.; DeCorby, M.; Karlowsky, J.A.; Walkty, A.; Hoban, D.J.; Zhanel, G.G. Activity of NXL104 in combination with β-lactams against genetically characterized Escherichia coli and Klebsiella pneumoniae isolates producing class A extended-spectrum β-lactamases and class C β-lactamases. Antimicrob. Agents Chemother. 2011, 55, 2434–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauernfeind, A.; Schweighart, S.; Grimm, H. A new plasmidic cefotaximase in a clinical isolate of Escherichia coli. Infection 1990, 18, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, A.; Holley, M.; Jungwirth, R.; Mangold, P.; Röhnisch, T.; Schweighart, S.; Wilhelm, R.; Casellas, J.M.; Goldberg, M. A new plasmidic cefotaximase from patients infected with Salmonella typhimurium. Infection 1992, 20, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Hawkey, P.M.; Jones, A.M. The changing epidemiology of resistance. J. Antimicrob. Chemother. 2009, 64, i3–i10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Wintersdorff, C.J.H.; Penders, J.; Van Niekerk, J.M.; Mills, N.D.; Majumder, S.; Van Alphen, L.B.; Savelkoul, P.H.M.; Wolffs, P.F.G. Dissemination of antimicrobial resistance in microbial ecosystems through horizontal gene transfer. Front. Microbiol. 2016, 7, 173. [Google Scholar] [CrossRef] [Green Version]
- Lenski, R.E. Experimental evolution and the dynamics of adaptation and genome evolution in microbial populations. ISME J. 2017, 11, 2181–2194. [Google Scholar] [CrossRef] [Green Version]
- Norman, A.; Hansen, L.H.; Sørensen, S.J. Conjugative plasmids: Vessels of the communal gene pool. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 2275–2289. [Google Scholar] [CrossRef] [Green Version]
- Dhanji, H.; Khan, P.; Cottell, J.L.; Piddock, L.J.V.; Zhang, J.; Livermore, D.M.; Woodford, N. Dissemination of pCT-Like IncK plasmids harboring CTX-M-14 extended-spectrum β-lactamase among clinical Escherichia coli isolates in the United Kingdom. Antimicrob. Agents Chemother. 2012, 56, 3376–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossolini, G.M.; D’Andrea, M.M.; Mugnaioli, C. The spread of CTX-M-type extended-spectrum β-lactamases. Clin. Microbiol. Infect. 2008, 14, 33–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toleman, M.A.; Bennett, P.M.; Walsh, T.R. ISCR Elements: Novel Gene-Capturing Systems of the 21st Century? Microbiol. Mol. Biol. Rev. 2006, 70, 296–316. [Google Scholar] [CrossRef] [Green Version]
- Lartigue, M.F.; Poirel, L.; Aubert, D.; Nordmann, P. In vitro analysis of ISEcp1B-mediated mobilization of naturally occurring β-lactamase gene blaCTX-M of Kluyvera ascorbata. Antimicrob. Agents Chemother. 2006, 50, 1282–1286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, A.; Coque, T.M.; Alonso, D.; Valverde, A.; Baquero, F.; Cantón, R. CTX-M-10 linked to a Phage-Related Element is Widely Disseminated among Enterobacteriaceae in a Spanish Hospital. Antimicrob. Agents Chemother. 2005, 49, 1567–1571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, R.M.; Collis, C.M. Mobile gene cassettes and integrons: Capture and spread of genes by site-specific recombination. Mol. Microbiol. 1995, 15, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Leverstein-van Hall, M.A.; Blok, H.E.M.; Donders, A.R.T.; Paauw, A.; Fluit, A.C.; Verhoef, J. Multidrug Resistance among Enterobacteriaceae Is Strongly Associated with the Presence of Integrons and Is Independent of Species or Isolate Origin. J. Infect. Dis. 2003, 187, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Carattoli, A. Animal reservoirs for extended spectrum β-lactamase producers. Clin. Microbiol. Infect. 2008, 14, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Poirel, L.; Bonnin, R.A.; Nordmann, P. Genetic support and diversity of acquired extended-spectrum β-lactamases in Gram-negative rods. Infect. Genet. Evol. 2012, 12, 883–893. [Google Scholar] [CrossRef]
- Yuan, W.; Tian, T.; Yang, Q.; Riaz, L. Transfer potentials of antibiotic resistance genes in Escherichia spp. strains from different sources. Chemosphere 2020, 246, 125736. [Google Scholar] [CrossRef]
- O’Brien, T.F. Emergence, Spread, and Environmental Effect of Antimicrobial Resistance: How Use of an Antimicrobial Anywhere Can Increase Resistance to Any Antimicrobial Anywhere Else. Clin. Infect. Dis. 2002, 34, S78–S84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and combating antibiotic resistance from One Health and Global Health perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.M.; Stegger, M.; Aziz, M.; Johnson, T.J.; Waits, K.; Nordstrom, L.; Gauld, L.; Weaver, B.; Statham, S.; Horwinski, J.; et al. Escherichia coli ST131-H22 as a Foodborne Uropathogen. MBio 2018, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, B.A.; Sidjabat, H.E.; Paterson, D.L. Escherichia coli O25b-ST131: A pandemic, multiresistant, community-associated strain. J. Antimicrob. Chemother. 2011, 66, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Vieira, A.R.; Collignon, P.; Aarestrup, F.M.; McEwen, S.A.; Hendriksen, R.S.; Hald, T.; Wegener, H.C. Association between antimicrobial resistance in Escherichia coli isolates from food animals and blood stream isolates from humans in Europe: An ecological study. Foodborne Pathog. Dis. 2011, 8, 1295–1301. [Google Scholar] [CrossRef] [PubMed]
- de Been, M.; Lanza, V.F.; de Toro, M.; Scharringa, J.; Dohmen, W.; Du, Y.; Hu, J.; Lei, Y.; Li, N.; Tooming-Klunderud, A.; et al. Dissemination of Cephalosporin Resistance Genes between Escherichia coli Strains from Farm Animals and Humans by Specific Plasmid Lineages. PLoS Genet. 2014, 10, e1004776. [Google Scholar] [CrossRef]
- Madec, J.; Haenni, M.; Ponsin, C.; Kieffer, N.; Rion, E.; Gassilloud, B. Sequence Type 48 Escherichia coli Carrying the blaCTX-M-1 IncI1/ST3 Plasmid in Drinking Water in France. Antimicrob. Agents Chemother. 2016, 60, 6430–6432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schijven, J.F.; Blaak, H.; Schets, F.M.; De Roda Husman, A.M. Fate of Extended-Spectrum β-Lactamase-Producing Escherichia coli from Faecal Sources in Surface Water and Probability of Human Exposure through Swimming. Environ. Sci. Technol. 2015, 49, 11825–11833. [Google Scholar] [CrossRef]
- Savin, M.; Bierbaum, G.; Hammerl, J.A.; Heinemann, C.; Parcina, M.; Sib, E.; Voigt, A.; Kreyenschmidt, J. Antibiotic-resistant bacteria and antimicrobial residues in wastewater and process water from German pig slaughterhouses and their receiving municipal wastewater treatment plants. Sci. Total Environ. 2020, 727, 138788. [Google Scholar] [CrossRef]
- Schmithausen, R.M.; Schulze-Geisthoevel, S.V.; Stemmer, F.; El-Jade, M.; Reif, M.; Hack, S.; Meilaender, A.; Montabauer, G.; Fimmers, R.; Parcina, M.; et al. Analysis of transmission of MRSA and ESBL-E among pigs and farm personnel. PLoS ONE 2015, 10, e0138173. [Google Scholar] [CrossRef]
- Oniciuc, E.A.; Likotrafiti, E.; Alvarez-Molina, A.; Prieto, M.; López, M.; Alvarez-Ordóñez, A. Food processing as a risk factor for antimicrobial resistance spread along the food chain. Curr. Opin. Food Sci. 2019, 30, 21–26. [Google Scholar] [CrossRef]
- Lavilla, S.; González-López, J.J.; Miró, E.; Domínguez, A.; Llagostera, M.; Bartolomé, R.M.; Mirelis, B.; Navarro, F.; Prats, G. Dissemination of extended-spectrum β-lactamase-producing bacteria: The food-borne outbreak lesson. J. Antimicrob. Chemother. 2008, 61, 1244–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerum, A.M.; Heuer, O.E. Human Health Hazards from Antimicrobial-Resistant Escherichia coli of Animal Origin. Clin. Infect. Dis. 2009, 48, 916–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barton, M.D. Impact of antibiotic use in the swine industry. Curr. Opin. Microbiol. 2014, 19, 9–15. [Google Scholar] [CrossRef]
- European Parliament and the Council of the European Union. Regulation (EC) No 1831/2003 of the European Parliament and of the Council on additives for use in animal nutrition. Off. J. Eur. Union 2003, 4, 29–43. Available online: http://eur-lex.europa.eu/legal-content/PT/TXT/?uri=celex:32003R1831 (accessed on 22 June 2020).
- Xiao, Y. A National Action Plan to Contain Antimicrobial Resistance in China: Contents, Actions and Expectations. Available online: http://resistancecontrol.info/2017/a-national-action-plan-to-contain-antimicrobial-resistance-in-china-contents-actions-and-expectations/ (accessed on 23 July 2020).
- Yang, H.; Paruch, L.; Chen, X.; van Eerde, A.; Skomedal, H.; Wang, Y.; Liu, D.; Clarke, J.L. Antibiotic application and resistance in swine production in China: Current situation and future perspectives. Front. Vet. Sci. 2019, 6, 136. [Google Scholar] [CrossRef] [Green Version]
- Chantziaras, I.; Boyen, F.; Callens, B.; Dewulf, J. Correlation between veterinary antimicrobial use and antimicrobial resistance in food-producing animals: A report on seven countries. J. Antimicrob. Chemother. 2014, 69, 827–834. [Google Scholar] [CrossRef] [Green Version]
- Fournier, C.; Aires-de-Sousa, M.; Nordmann, P.; Poirel, L. Occurrence of CTX-M-15- and MCR-1-producing Enterobacterales in pigs in Portugal: Evidence of direct links with antibiotic selective pressure. Int. J. Antimicrob. Agents 2020, 55, 105802. [Google Scholar] [CrossRef]
- Done, H.Y.; Venkatesan, A.K.; Halden, R.U. Does the Recent Growth of Aquaculture Create Antibiotic Resistance Threats Different from those Associated with Land Animal Production in Agriculture? AAPS J. 2015, 17, 513–524. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency, European Surveillance of Veterinary Antimicrobial Consumption, 2019. Sales of Veterinary Antimicrobial Agents in 31 European Countries in 2017. Available online: https://www.ema.europa.eu/en/documents/report/sales-veterinary-antimicrobial-agents-31-european-countries-2017_en.pdf?elqTrackId=2383f3a8f966419299c3ce3b4258052f&elq=65220e1bd1e34bcaa96b0addf44fdf3e&elqaid=28856&elqat=1&elqCampaignId=20867 (accessed on 23 July 2020).
- Dohmen, W.; Schmitt, H.; Bonten, M.; Heederik, D. Air exposure as a possible route for ESBL in pig farmers. Environ. Res. 2017, 155, 359–364. [Google Scholar] [CrossRef]
- European Food Safety Authority; European Centre for Disease Prevention and Control. The European Union Summary Report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2012. EFSA J. 2014, 12, 3590. [Google Scholar] [CrossRef]
- AgersoØ, Y.; Aarestrup, F.M. Voluntary ban on cephalosporin use in Danish pig production has effectively reduced extended-spectrum cephalosporinase-producing Escherichia coli in slaughter pigs. J. Antimicrob. Chemother. 2013, 68, 569–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hammerum, A.M.; Larsen, J.; Andersen, V.D.; Lester, C.H.; Skytte, T.S.S.; Hansen, F.; Olsen, S.S.; Mordhorst, H.; Skov, R.L.; Aarestrup, F.M.; et al. Characterization of extended-spectrum β-lactamase (ESBL)-producing Escherichia coli obtained from Danish pigs, pig farmers and their families from farms with high or no consumption of third- or fourth-generation cephalosporins. J. Antimicrob. Chemother. 2014, 69, 2650–2657. [Google Scholar] [CrossRef] [PubMed]
- Andersen, V.D.; Jensen, V.F.; Vigre, H.; Andreasen, M.; Agersø, Y. The use of third and fourth generation cephalosporins affects the occurrence of extended-spectrum cephalosporinase-producing Escherichia coli in Danish pig herds. Vet. J. 2015, 204, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Wheatley, P.; Giotis, E.S.; McKevitt, A.I. Effects of slaughtering operations on carcass contamination in an Irish pork production plant. Ir. Vet. J. 2014, 67, 1–6. [Google Scholar] [CrossRef] [Green Version]
- European Commission. Commission Regulation (EC) No 2073/2005 on microbiological criteria for foodstuffs. Off. J. Eur. Union 2005, 338, 1–26. [Google Scholar] [CrossRef]
- Barco, L.; Belluco, S.; Roccato, A.; Ricci, A. Escherichia coli and Enterobacteriaceae counts on pig and ruminant carcasses along the slaughterline, factors influencing the counts and relationship between visual faecal contamination of carcasses and counts: A review. EFSA Support. Publ. 2014, 11, 634E. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United States. Animal Production and Health. Available online: http://www.fao.org/ag/againfo/themes/en/meat/backgr_sources.html (accessed on 22 June 2020).
- European Commission. Information on Pork Production, Legal Bases, Market Monitoring, Trade, Carcass Classification, Price Reporting, Regulation of Supply of PDO/PGI Ham. Available online: https://ec.europa.eu/info/food-farming-fisheries/animals-and-animal-products/animal-products/pork_en#introduction (accessed on 22 June 2020).
- European Commission. Pork Production up in the EU. 2017. Available online: https://ec.europa.eu/eurostat/web/products-eurostat-news/-/DDN-20170919-1 (accessed on 22 June 2020).
- Aasmäe, B.; Häkkinen, L.; Kaart, T.; Kalmus, P. Antimicrobial resistance of Escherichia coli and Enterococcus spp. isolated from Estonian cattle and swine from 2010 to 2015. Acta Vet. Scand. 2019, 61, 5. [Google Scholar] [CrossRef]
- Kojima, A.; Ishii, Y.; Ishihara, K.; Esaki, H.; Asai, T.; Oda, C.; Tamura, Y.; Yamaguchi, K. Extended-Spectrum-Lactamase-Producing. Antimicrob. Agents Chemother. 2005, 49, 3533–3537. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; White, D.G.; Mcdermott, P.F.; Friedman, S.; English, L.; Ayers, S.; Meng, J.; Maurer, J.J.; Holland, R.; Walker, R.D. Identifcation and Expression of Cephamycinase blaCMY Genes in Escherichia coli and Salmonella Isolates from Food Animals and Ground Meat. Antimicrob. Agents Chemother. 2001, 45, 3647–3650. [Google Scholar] [CrossRef] [Green Version]
- van Hoek, A.H.A.M.; Veenman, C.; van Overbeek, W.M.; Lynch, G.; de Roda Husman, A.M.; Blaak, H. Prevalence and characterization of ESBL- and AmpC-producing Enterobacteriaceae on retail vegetables. Int. J. Food Microbiol. 2015, 204, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Moodley, A.; Guardabassi, L. Transmission of IncN plasmids carrying blaCTX-M-1 between commensal escherichia coli in pigs and farm workers. Antimicrob. Agents Chemother. 2009, 53, 1709–1711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dahliani, E.R.; Rahmatan, H. Djufri The correlation between students’ interest and learning outcomes in biology. J. Phys. Conf. Ser. 2020, 1460, 1–6. [Google Scholar] [CrossRef]
- Agersø, Y.; Aarestrup, F.M.; Pedersen, K.; Seyfarth, A.M.; Struve, T.; Hasman, H. Prevalence of extended-spectrum cephalosporinase (ESC)-producing Escherichia coli in Danish slaughter pigs and retail meat identified by selective enrichment and association with cephalosporin usage. J. Antimicrob. Chemother. 2012, 67, 582–588. [Google Scholar] [CrossRef] [Green Version]
- Skočková, A.; Koláčková, I.; Bogdanovičová, K.; Karpíšková, R. Characteristic and antimicrobial resistance in Escherichia coli from retail meats purchased in the Czech Republic. Food Control. 2015, 47, 401–406. [Google Scholar] [CrossRef]
- Barilli, E.; Vismarra, A.; Villa, Z.; Bonilauri, P.; Bacci, C. ESβL E. coli isolated in pig’s chain: Genetic analysis associated to the phenotype and biofilm synthesis evaluation. Int. J. Food Microbiol. 2019, 289, 162–167. [Google Scholar] [CrossRef]
- Biasino, W.; De Zutter, L.; Garcia-Graells, C.; Uyttendaele, M.; Botteldoorn, N.; Gowda, T.; Van Damme, I. Quantification, distribution and diversity of ESBL/AmpC-producing Escherichia coli on freshly slaughtered pig carcasses. Int. J. Food Microbiol. 2018, 281, 32–35. [Google Scholar] [CrossRef]
- Sabia, C.; Stefani, S.; Messi, P.; de Niederhäusern, S.; Bondi, M.; Condò, C.; Iseppi, R.; Anacarso, I. Extended-Spectrum Β-Lactamase and Plasmid-Mediated AMPC Genes in Swine and Ground Pork. J. Food Saf. 2017, 37, e12282. [Google Scholar] [CrossRef] [Green Version]
- Randall, L.P.; Lodge, M.P.; Elviss, N.C.; Lemma, F.L.; Hopkins, K.L.; Teale, C.J.; Woodford, N. Evaluation of meat, fruit and vegetables from retail stores in five United Kingdom regions as sources of extended-spectrum beta-lactamase (ESBL)-producing and carbapenem-resistant Escherichia coli. Int. J. Food Microbiol. 2017, 241, 283–290. [Google Scholar] [CrossRef]
- Vogt, D.; Overesch, G.; Endimiani, A.; Collaud, A.; Thomann, A.; Perreten, V. Occurrence and genetic characteristics of third-generation cephalosporin-resistant Escherichia coli in Swiss retail meat. Microb. Drug Resist. 2014, 20, 485–494. [Google Scholar] [CrossRef] [Green Version]
- Kaesbohrer, A.; Bakran-Lebl, K.; Irrgang, A.; Fischer, J.; Kämpf, P.; Schiffmann, A.; Werckenthin, C.; Busch, M.; Kreienbrock, L.; Hille, K. Diversity in prevalence and characteristics of ESBL/pAmpC producing E. coli in food in Germany. Vet. Microbiol. 2019, 233, 52–60. [Google Scholar] [CrossRef] [PubMed]
- Wasiński, B.; Rózanska, H.; Osek, J. Occurrence of extended spectrum ß-Lactamaseand AmpC-Producing Escherichia coli in meat samples. Bull. Vet. Inst. Pulawy 2013, 57, 513–517. [Google Scholar] [CrossRef] [Green Version]
- Egervärn, M.; Börjesson, S.; Byfors, S.; Finn, M.; Kaipe, C.; Englund, S.; Lindblad, M. Escherichia coli with extended-spectrum beta-lactamases or transferable AmpC beta-lactamases and Salmonella on meat imported into Sweden. Int. J. Food Microbiol. 2014, 171, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Springer, B.; Bruckner, K. Characterization of extended-spectrum beta-lactamase (ESBL) producing Escherichia coli from raw meat and comparison to human isolates. Wien. Tierarztl Monatsschr 2012, 99, 44–50. [Google Scholar]
- Thorsteinsdottir, T.R.; Haraldsson, G.; Fridriksdottir, V.; Kristinsson, K.G.; Gunnarsson, E. Prevalence and genetic relatedness of antimicrobial-resistant escherichia coli isolated from animals, foods and humans in Iceland. Zoonoses Public Health 2010, 57, 189–196. [Google Scholar] [CrossRef]
- Paterson, D.L.; Egea, P.; Pascual, A.; López-Cerero, L.; Navarro, M.D.; Adams-Haduch, J.M.; Qureshi, Z.A.; Sidjabat, H.E.; Rodríguez-Baño, J. Extended-spectrum and CMY-type β-lactamase-producing Escherichia coli in clinical samples and retail meat from Pittsburgh, USA and Seville, Spain. Clin. Microbiol. Infect. 2010, 16, 33–38. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| β-Lactamase | Molecular Weight of the Enzyme | Bush-Jacoby Medeiros Class | Ambler Class | Active Site | Preferential Substrate | Gene | Organisms |
|---|---|---|---|---|---|---|---|
| TEM | ~29,000 Da | 2be | A | Serine | Penicillins and 1st–4th generation cephalosporins | Pl* | Enterobacteriaceae, Haemophilus influenzae, Neisseria gonorrhoeae, Pseudomonas aeruginosa |
| SHV | Enterobacteriaceae | ||||||
| CTX-M | Enterobacteriaceae |
| Region | Country | Penicillin Sales for Food-Producing Animals, (mg/PCU) in 2017 | 1st- and 2nd-gen. Cephalosporin Sales for Food-Producing Animals, (mg/PCU) in 2017 | 3rd- and 4th-gen. Cephalosporin Sales for Food-Producing Animals, (mg/PCU) in 2017 |
|---|---|---|---|---|
| Northern Europe | United Kingdom | 7.5 | 0.1 | 0.1 |
| Sweden | 7.7 | <0.01 | <0.01 | |
| Finland | 8.8 | 0.03 | <0.01 | |
| Latvia | 8.5 | 0.2 | 0.3 | |
| Lithuania | 13.6 | 0.1 | 0.2 | |
| Ireland | 9.7 | 0.3 | 0.1 | |
| Estonia | 21.7 | 0.2 | 0.8 | |
| Denmark | 11.9 | 0.02 | <0.01 | |
| Iceland | 3.3 | 0 | <0.01 | |
| Norway | 1.6 | 0 | <0.01 | |
| Western Europe | Luxembourg | 6.8 | 0.1 | 0.6 |
| Netherlands | 12.6 | 0.04 | <0.01 | |
| France | 9.3 | 0.2 | 0.02 | |
| Germany | 34.5 | 0.1 | 0.4 | |
| Belgium | 49.4 | 0.2 | 0.1 | |
| Austria | 8.6 | 0.05 | 0.2 | |
| Switzerland | 11.2 | 0.1 | 0.2 | |
| Eastern Europe | Bulgaria | 24.7 | 0.02 | 0.1 |
| Czech Republic | 17.3 | 0.3 | 0.5 | |
| Croatia | 22.3 | 0.2 | 0.2 | |
| Hungary | 46.7 | 0.1 | 0.5 | |
| Poland | 54.1 | 0.1 | 0.2 | |
| Romania | 18.2 | <0.01 | 0.2 | |
| Slovakia | 11.4 | 0.4 | 0.4 | |
| Southern Europe | Cyprus | 81.1 | 0.03 | 0.4 |
| Spain | 58.5 | 0.03 | 0.2 | |
| Greece | 18.6 | <0.01 | 0.1 | |
| Italy | 70.3 | 0.2 | 0.4 | |
| Malta | 12.1 | 0.1 | 0.2 | |
| Portugal | 35.1 | 0.1 | 0.6 | |
| Slovenia | 22.2 | 0.1 | 0.2 | |
| Total | 23.2 | 0.1 | 0.3 |
| Region | Country | Resistance to Cefotaxime in E. coli Isolates from Pigs % (2012) | Resistance to Ceftazidime in E. coli Isolates from Pigs % (2012) | Presumptive ESBL E. coli Prevalence in Fattening Pigs % (2015) | Presumptive ESBL E. coli Prevalence in Pork Meat % (2015) | Presumptive ESBL E. coli Prevalence in Fattening Pigs % (2017) | Presumptive ESBL E. coli Prevalence in Pork Meat % (2017) |
|---|---|---|---|---|---|---|---|
| Northern Europe | United Kingdom | - | - | 21.7 | 2.1 | 16.1 | 0 |
| Sweden | - | - | 1 | 0.3 | 3.7 | 0 | |
| Finland | - | - | 0.3 | 0 | 0.3 | 0 | |
| Latvia | - | - | 40 | 6.5 | 42.3 | 8.1 | |
| Lithuania | - | - | 17.9 | 8.7 | 37.9 | 2.7 | |
| Ireland | - | - | 10.3 | - | 18.5 | 2.3 | |
| Estonia | - | - | 29.9 | 2.7 | 29.4 | 1.3 | |
| Denmark | 0.7 | - | 7 | 1.2 | 6.8 | 2.1 | |
| Iceland | - | - | - | - | 0 | 0 | |
| Norway | - | - | 0.4 | 0 | 0.7 | 0 | |
| Western Europe | Luxembourg | - | - | 48.3 | - | 38.1 | 0 |
| Netherlands | 0 | 0 | 10.3 | - | 11 | 0.7 | |
| France | 2 | 2 | 34.7 | 1.5 | 23.8 | 0.3 | |
| Germany | - | - | 39.5 | 5.5 | 41.8 | 4.9 | |
| Belgium | 2.9 | 3.4 | 54.7 | 11.8 | 60.7 | 4.3 | |
| Austria | 0 | 0 | 48.2 | 9.3 | 58.8 | 9.4 | |
| Switzerland | 1.1 | 1.1 | 17 | 1 | 11.5 | 0.3 | |
| Eastern Europe | Bulgaria | - | - | 49.8 | 20.8 | 42 | 10 |
| Czech Republic | - | - | 21.5 | 9.3 | 17.6 | 10.1 | |
| Croatia | - | - | 17.6 | 1.4 | 30.2 | 5.1 | |
| Hungary | 1.5 | - | 44.7 | 7.7 | 56.2 | 3.7 | |
| Poland | 2.6 | 2.6 | 26.8 | - | 31.9 | 3.7 | |
| Romania | - | - | 46.6 | 11.1 | 53.7 | 9.7 | |
| Slovakia | - | - | 9.1 | 2 | 34 | 5.3 | |
| Southern Europe | Cyprus | - | - | 6.8 | 1.7 | 0.8 | 1.4 |
| Spain | - | - | 81.5 | 9.6 | 80.3 | 9.7 | |
| Greece | - | - | 32.8 | 5.8 | 33.5 | 6.8 | |
| Italy | - | - | 64a | 7.9a | 69.2 | 7 | |
| Malta | - | - | - | - | 17.9 | 11.1 | |
| Portugal | - | - | 65.2 | 21.3 | 54.9 | 8.2 | |
| Slovenia | - | - | 28.5 | 4.7 | 25.7 | 2 | |
| Total | 1.4 | 1.5 | 30.2 | 6.2 | 30.6 | 4.2 |
| Antibiotic Sales | Prevalence | Correlation Coefficient | Correlation Level [111] | |
|---|---|---|---|---|
| Penicillin sales for food-producing animals, (mg/PCU) | & | Presumptive ESBL E. coli prevalence in fattening pigs | 0.464 | Middle |
| 1st and 2nd generation cephalosporin sales for food-producing animals, (mg/PCU) | & | Presumptive ESBL E. coli prevalence in fattening pigs | 0.180 | Very low |
| 3rd and 4th generation cephalosporin sales for food-producing animals, (mg/PCU) | & | Presumptive ESBL E. coli prevalence in fattening pigs | 0.074 | Very low |
| Prevalence in Fattening Pigs | Prevalence in Pork Meat | Correlation Coefficient | Correlation Level [111] | |
|---|---|---|---|---|
| Presumptive ESBL E. coli prevalence in fattening pigs in 2017 | & | Presumptive ESBL E. coli prevalence in pork meat in 2017 | 0.626 | Strong |
| Presumptive ESBL E. coli prevalence in fattening pigs in 2015 | & | Presumptive ESBL E. coli prevalence in pork meat in 2015 | 0.735 | Strong |
| Meat Origin | Sampling Period | Type of Meat | Meat Origin | Number of Samples | Prevalence of ESBL Producing E. coli (%) | ESBL Encoding Genes | Reference | ||
|---|---|---|---|---|---|---|---|---|---|
| CTX-M (%) | TEM (%) | SHV (%) | |||||||
| Italy | 2016–2017 | Carcasses | Slaughterhouse | 200 | 10 | 11.5 | 3.5 | 0.5 | [114] |
| Sausages, meat slices, loin, salami dough, cotechino, thighs for ham production | Supermarkets | 446 | 2 | 1.8 | 1.4 | 0.2 | |||
| Belgium | 2015–2016 | Head | Slaughterhouse | 104 | 25 | - | - | - | [115] |
| Belly | Slaughterhouse | 104 | 7 | - | - | - | |||
| Ham | Slaughterhouse | 103 | 3 | - | - | - | |||
| Loin | Slaughterhouse | 104 | 1 | - | - | - | |||
| Germany | 2014 | Carcasses | Meat processing company | 63 | 17.5 | 15.9 | 4.8 | 0 | [25] |
| Italy | 2013–2014 | Ground pork | Food market | 200 | 0 | - | - | - | [116] |
| England, Wales, and Scotland | 2013–2014 | Pork | Supermarkets, discount store, convenience stores, butchers | 79 | 3 | 2 | 0 | 0 | [117] |
| Switzerland | 2013 | Pork | Meat-packaging plant | 50 | 0 | - | - | - | [118] |
| Germany | 2012–2013 | Pork meat | Butcher shops, supermarkets, farmer’s markets, direct marketer, restaurants, canteens | 282 | 12.1 | 11.3 | 0.7 | 0.4 | [119] |
| Ground pork | 214 | 13.6 | 10.7 | 0.9 | 0.9 | ||||
| Poland | 2012–2013 | Pork | Slaughterhouse | 78 | 5 | 5.13 | 0 | 3.85 | [120] |
| Czech Republic | 2012–2013 | Pork | Supermarkets | 110 | 3.1 | 0 | 40.6 | 0 | [113] |
| Denmark | 2010–2011 | Frozen or fresh pork | Retail stores, outlets | 44 | 2 | 2.3 | 2.3 | 0 | [121] |
| Germany | 44 | 7 | 6.8 | 4.5 | 0 | ||||
| Italy | 20 | 15 | 10 | 10 | 0 | ||||
| Finland, The Netherlands, Poland, Spain | 31 | 9.6 | 0 | 0 | 0 | ||||
| Denmark | 2009 | Frozen or fresh pork | Retail store, outlets | 153 | 2 | 1.31 | 0 | 0 | [112] |
| Germanya | 142 | 0.7 | - | - | - | ||||
| The Netherlands | 16 | 0 | - | - | - | ||||
| Austria | 2009 | Ground pork | Supermarkets | 27 | 3.7 | - | - | - | [122] |
| Iceland | 2006–2007 | Pork | Meat-processing plants | 60 | 0 | - | - | - | [123] |
| Spain | 2006–2007 | Pork | Supermarket | 12 | 25 | - | - | - | [124] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bergšpica, I.; Kaprou, G.; Alexa, E.A.; Prieto, M.; Alvarez-Ordóñez, A. Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union. Antibiotics 2020, 9, 678. https://doi.org/10.3390/antibiotics9100678
Bergšpica I, Kaprou G, Alexa EA, Prieto M, Alvarez-Ordóñez A. Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union. Antibiotics. 2020; 9(10):678. https://doi.org/10.3390/antibiotics9100678
Chicago/Turabian StyleBergšpica, Ieva, Georgia Kaprou, Elena A. Alexa, Miguel Prieto, and Avelino Alvarez-Ordóñez. 2020. "Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union" Antibiotics 9, no. 10: 678. https://doi.org/10.3390/antibiotics9100678
APA StyleBergšpica, I., Kaprou, G., Alexa, E. A., Prieto, M., & Alvarez-Ordóñez, A. (2020). Extended Spectrum β-Lactamase (ESBL) Producing Escherichia coli in Pigs and Pork Meat in the European Union. Antibiotics, 9(10), 678. https://doi.org/10.3390/antibiotics9100678