Improved Cell Selectivity of Pseudin-2 via Substitution in the Leucine-Zipper Motif: In Vitro and In Vivo Antifungal Activity

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
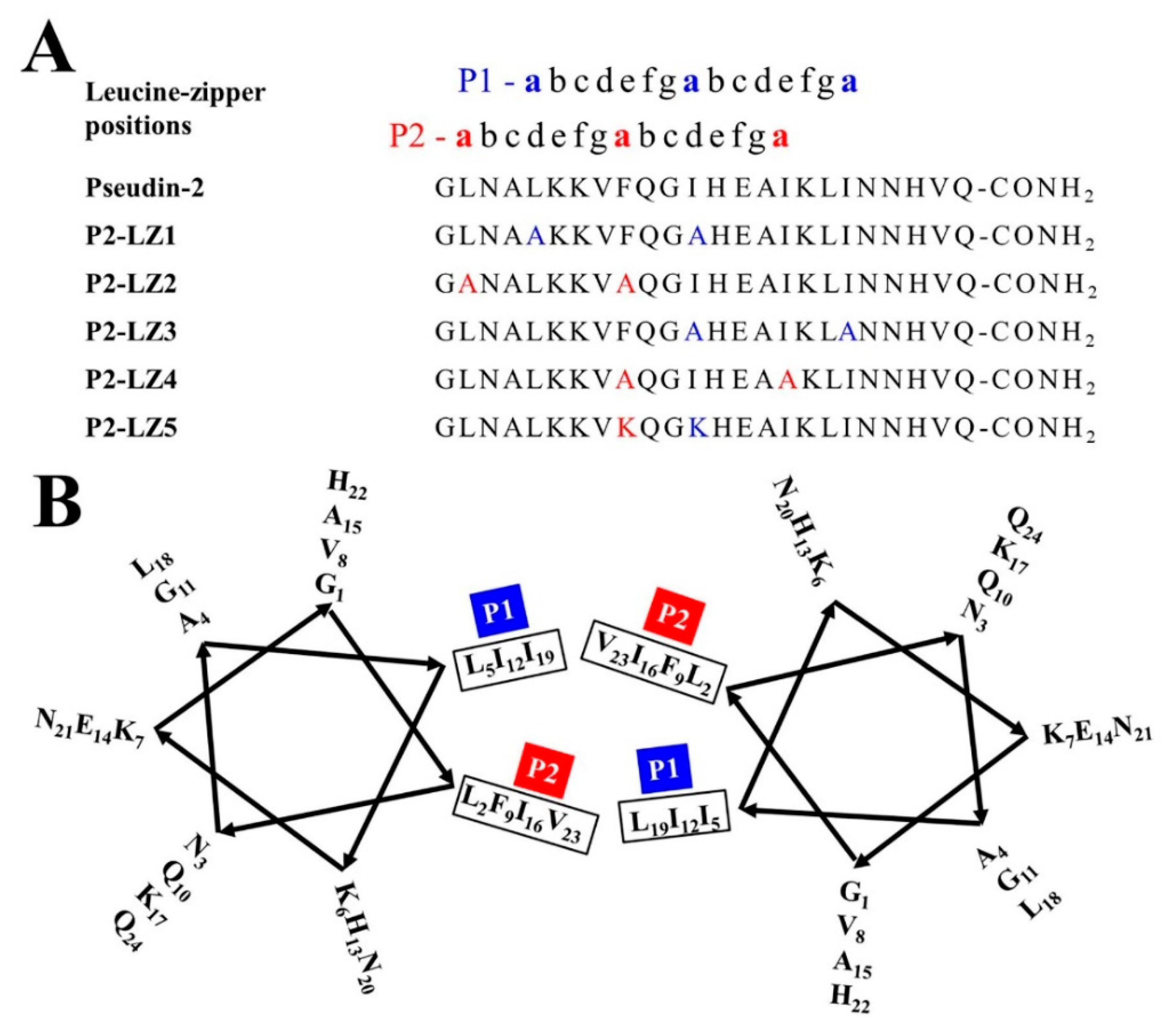
2.1. Designation of Pseudin-2 Derivatives Based on the LZ Motif and Structural Orientation
2.2. Cell-Selective Antifungal Activity of the Designed Peptides
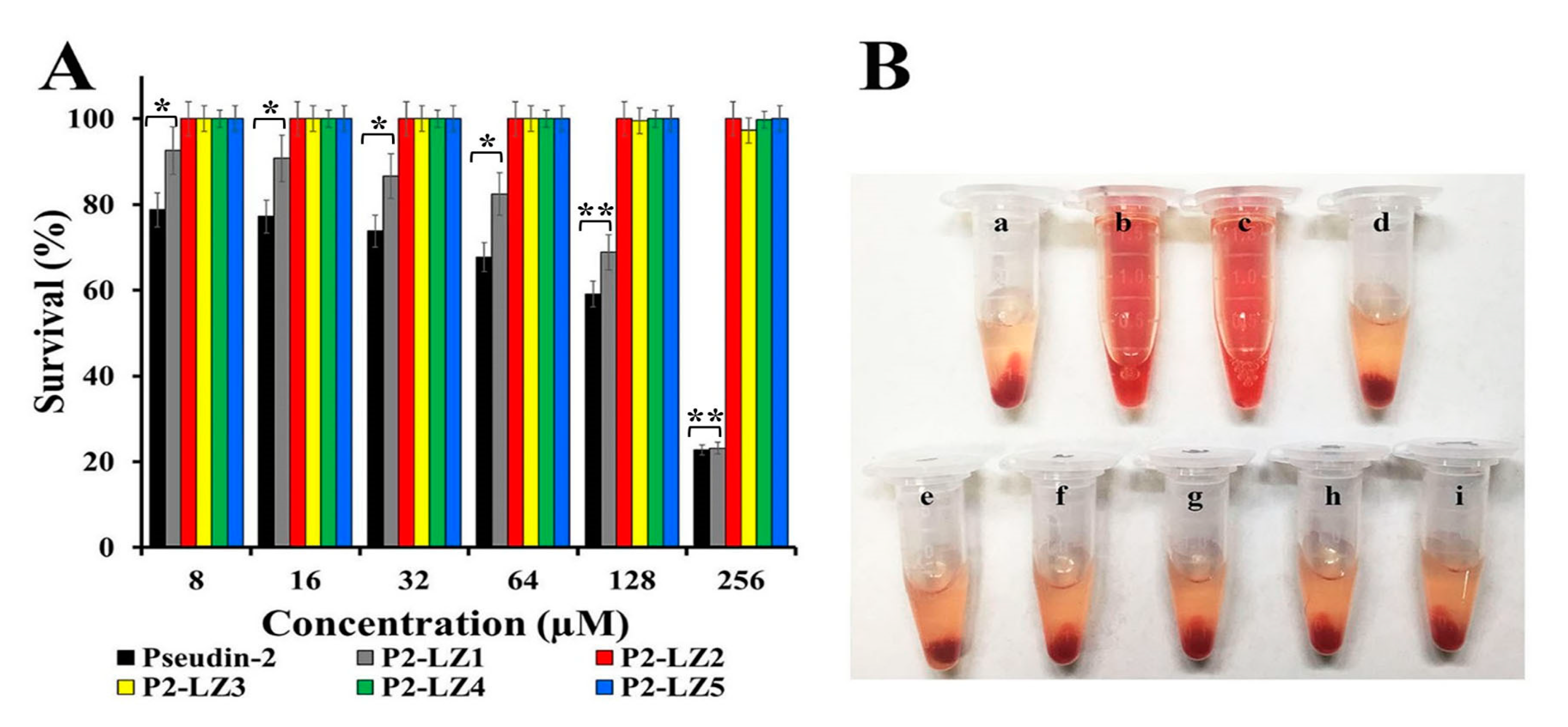
2.2.1. In Vitro Antifungal Activity
2.2.2. In Vitro Cytotoxic Effects
2.3. Mode of Action of the Designed Peptides
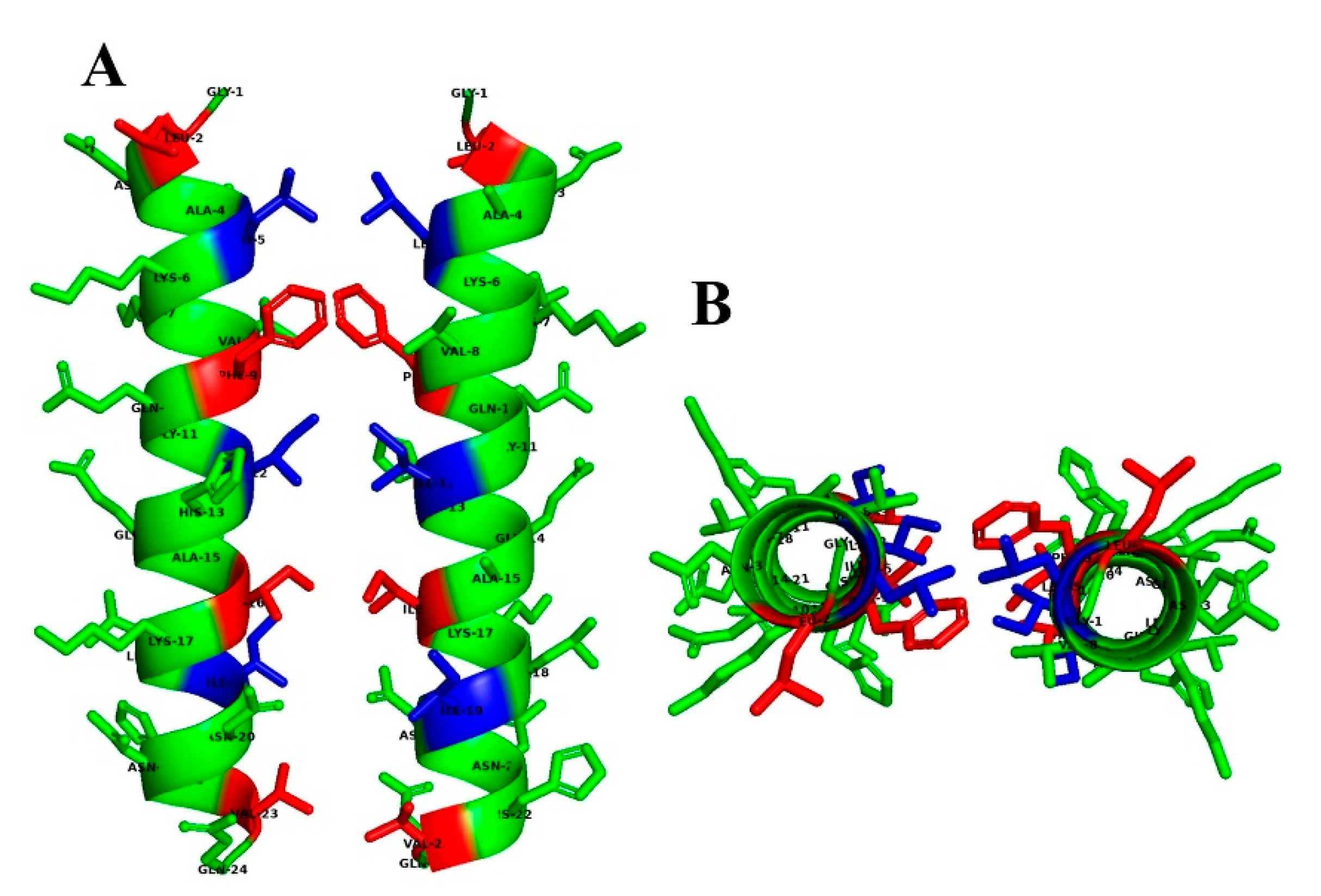
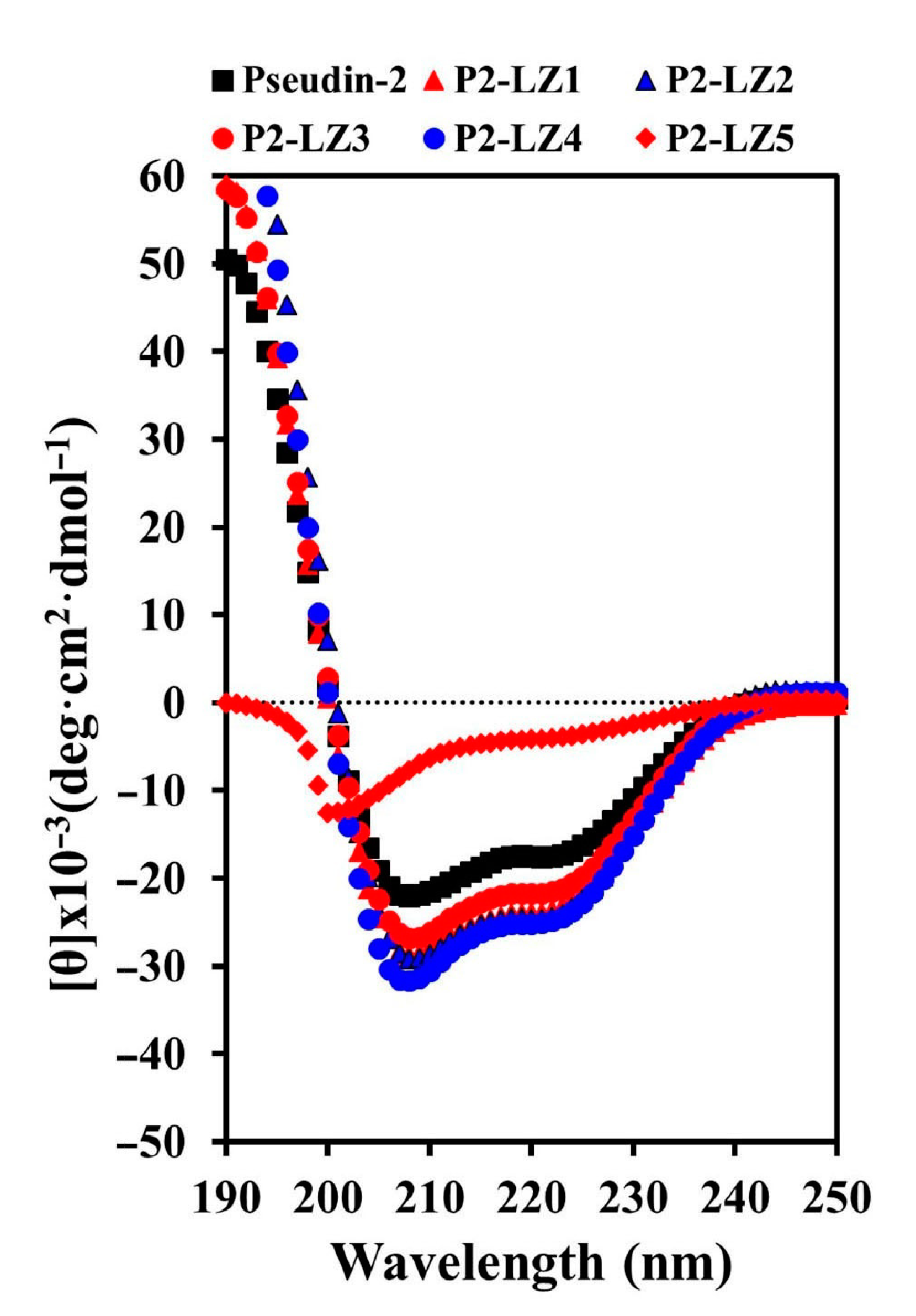
2.3.1. Secondary and Predicted 3D Structures of the Designed Peptides
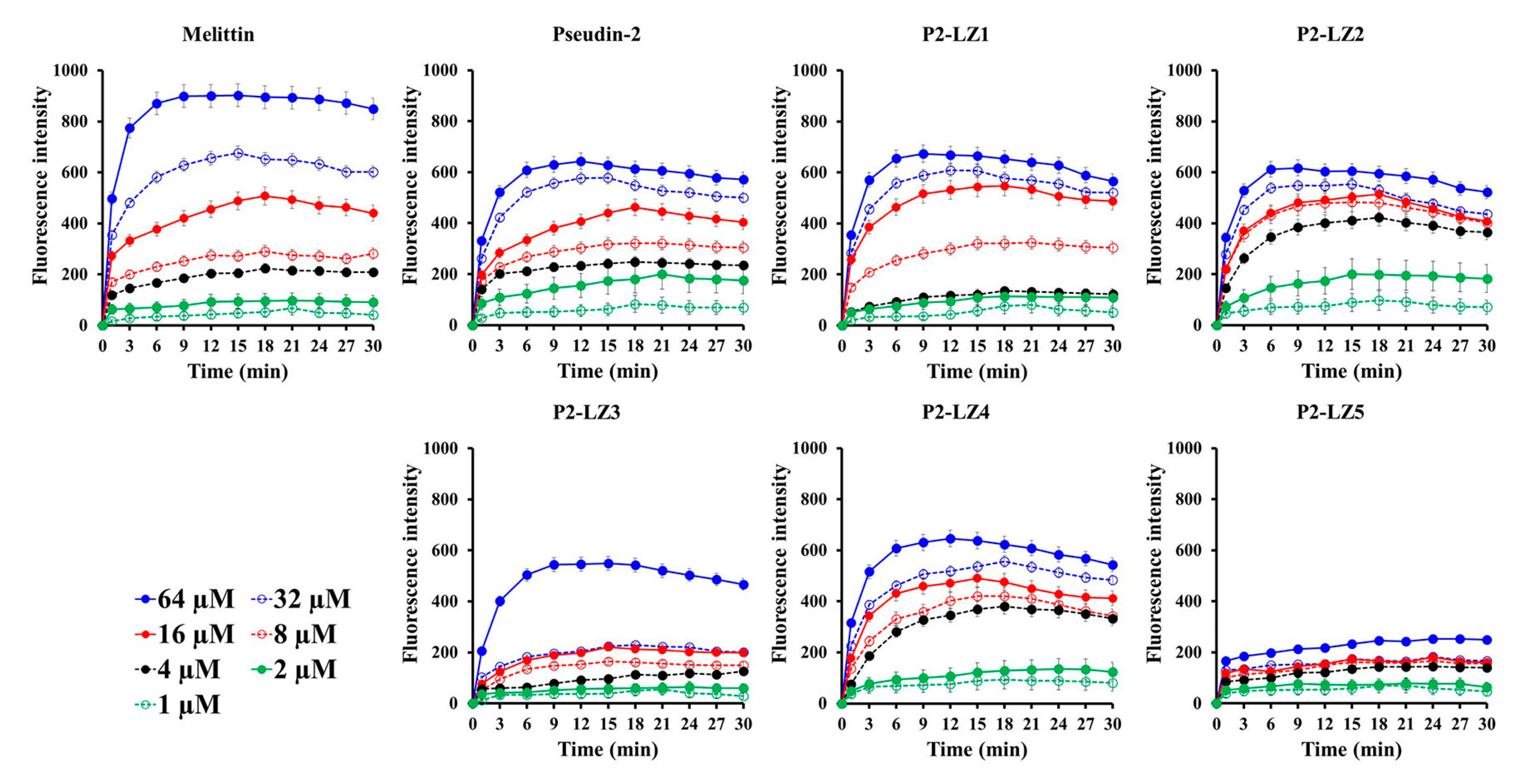
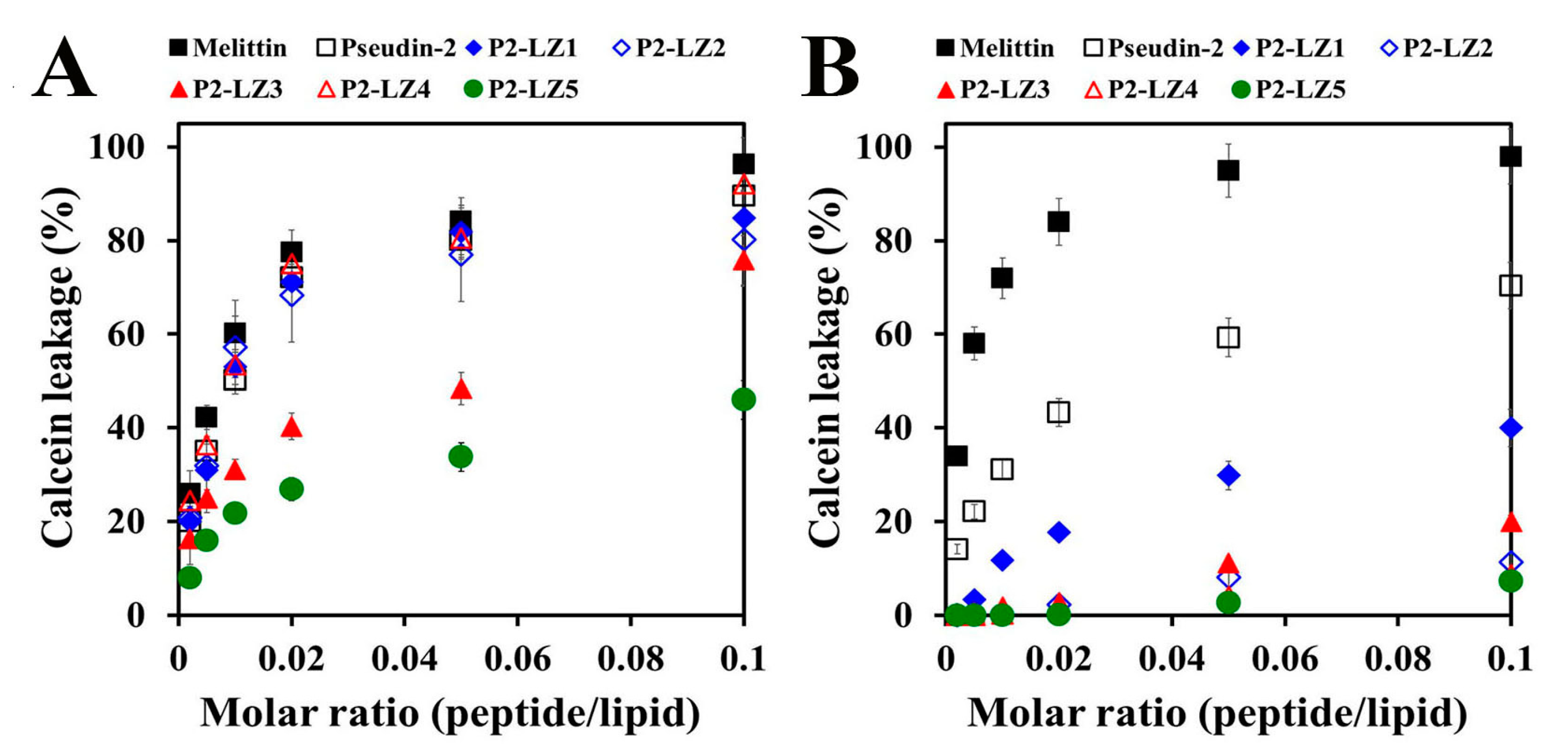
2.3.2. Mode of the Antifungal Actions of the Designed Peptides
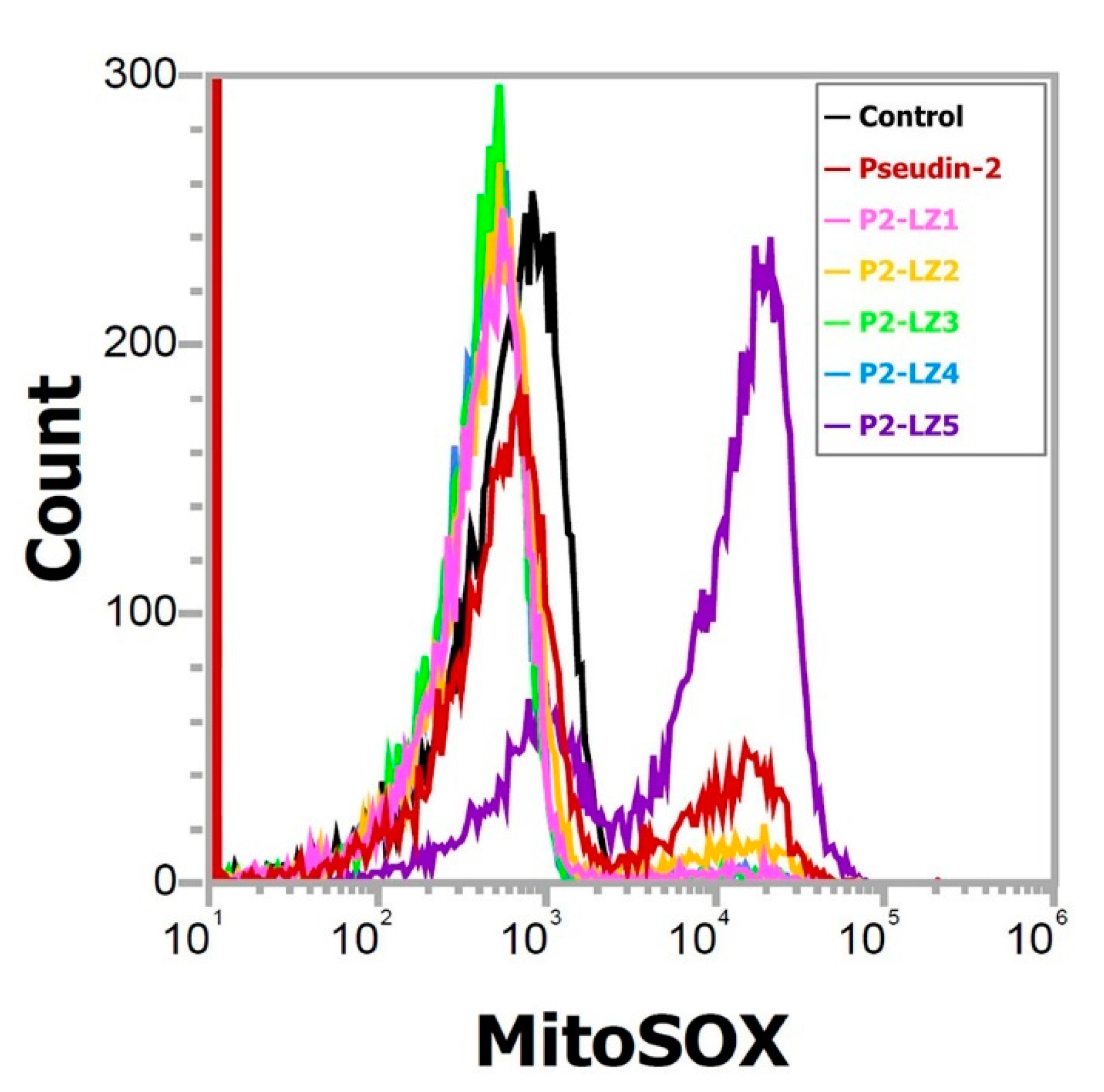
2.4. Mitochondrial Reactive Oxygen Species (ROS) Generation in Response to P2-LZ5
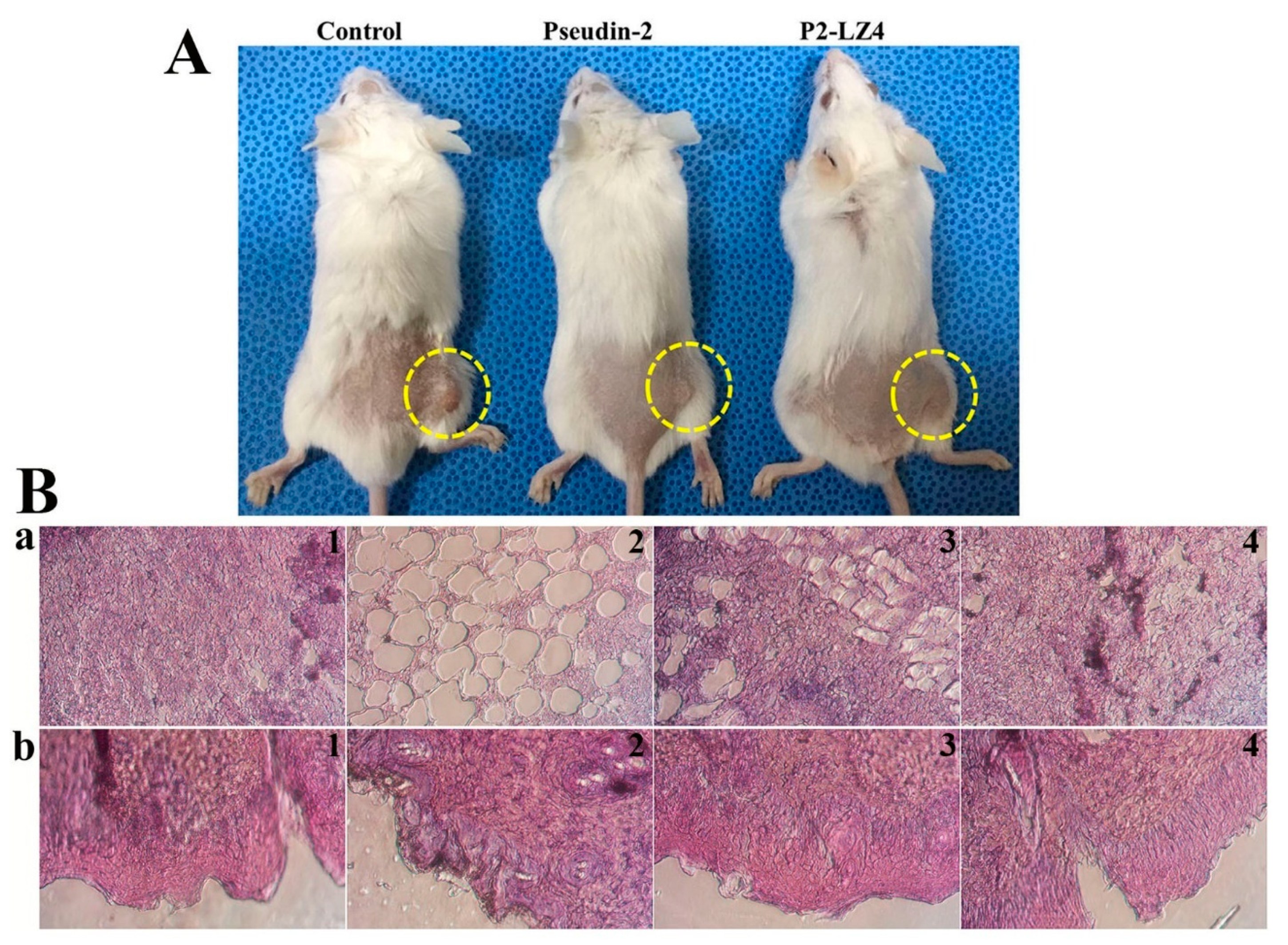
2.5. In Vivo Antifungal Effects
3. Materials and Methods
3.1. Materials
3.2. Fungal Strains and Growth
3.3. Peptide Synthesis
3.4. Prediction of the 3D Structure
3.5. Antifungal Assay
3.6. Cytotoxic and Hemolytic Assay
3.7. CD Analysis
3.8. CLSM
3.9. SYTOX Green Uptake
3.10. Calcein Leakage from Artificial Vesicles
3.11. SEM
3.12. Measurement of Mitochondrial SOX
3.13. In Vivo Animal Experiment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Microbiol. 2005, 3, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Cruz, J.; Ortiz, C.; Guzmán, F.; Fernández-Lafuente, R.; Torres, R. Antimicrobial peptides: Promising compounds against pathogenic microorganisms. Curr. Med. Chem. 2014, 21, 2299–2321. [Google Scholar] [CrossRef] [PubMed]
- Conlon, J.M.; Mechkarska, M.; Lukic, M.L.; Flatt, P.R. Potential therapeutic applications of multifunctional host-defense peptides from frog skin as anti-cancer, anti-viral, immunomodulatory, and anti-diabetic agents. Peptides 2014, 57, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.H.; Lu, T.K. Development and challenges of antimicrobial peptides for therapeutic applications. Antibiotics 2020, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Raheem, N.; Straus, S.K. Mechanisms of action for antimicrobial peptides with antibacterial and antibiofilm functions. Front. Microbiol. 2019, 10, 2866. [Google Scholar] [CrossRef] [Green Version]
- Boparai, J.K.; Sharma, P.K. Mini review on antimicrobial peptides, sources, mechanism and recent applications. Protein Pept. Lett. 2020, 2, 4–16. [Google Scholar] [CrossRef]
- Haney, E.F.; Straus, S.K.; Hancock, R.E.W. Reassessing the host defense peptide landscape. Front. Chem. 2019, 7, 43. [Google Scholar] [CrossRef] [Green Version]
- van der Does, A.M.; Hiemstra, P.S.; Mookherjee, N. Antimicrobial host defense peptides: Immunomodulatory functions and translational prospects. Adv. Exp. Med. Biol. 2019, 1117, 149–171. [Google Scholar]
- Rajchakit, U.; Sarojini, V. Recent developments in antimicrobial-peptide-conjugated gold nanoparticles. Bioconjug. Chem. 2017, 28, 2673–2686. [Google Scholar] [CrossRef]
- Scorciapino, M.A.; Serra, I.; Manzo, G.; Rinaldi, A.C. Antimicrobial dendrimeric peptides: Structure, activity and new therapeutic applications. Int. J. Mol. Sci. 2017, 18, 542. [Google Scholar] [CrossRef] [Green Version]
- Kouzarides, T.; Ziff, E. The role of the leucine zipper in the fos-jun interaction. Nature 1988, 336, 646–651. [Google Scholar] [CrossRef] [PubMed]
- Abbanat, D.; Morrow, B.; Bush, K. New agents in development for the treatment of bacterial infections. Curr. Opin. Pharm. 2008, 8, 582–592. [Google Scholar] [CrossRef]
- Karaiskos, I.; Giamarellou, H. Multidrug-resistant and extensively drug-resistant Gram-negative pathogens: Current and emerging therapeutic approaches. Expert Opin. Pharm. 2014, 15, 1351–1370. [Google Scholar] [CrossRef] [PubMed]
- Park, S.C.; Park, Y.; Hahm, K.S. The role of antimicrobial peptides in preventing multidrug-resistant bacterial infections and biofilm formation. Int. J. Mol. Sci. 2011, 12, 5971–5992. [Google Scholar] [CrossRef] [Green Version]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Pάl, T.; Sonnevend, A.; Galadari, S.; Conlon, J.M. Design of potent, non-toxic antimicrobial agents based upon the structure of the frog skin peptide, pseudin-2. Regul. Pept. 2005, 129, 85–91. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, J.Y.; Jeong, C.; Yoo, S.; Hahm, K.S.; Park, Y. A plausible mode of action of pseudin-2, an antimicrobial peptide from Pseudis paradoxa. Biochim. Biophys. Acta 2011, 1808, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.; Kim, J.; Choi, Y.; Bang, J.; Kim, Y. Antiseptic Effect of Ps-K18: Mechanism of Its Antibacterial and Anti-Inflammatory Activities. Int. J. Mol. Sci. 2019, 20, 4895. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.K.; Seo, C.H.; Luchian, T.; Park, Y. Pse-T2, an Antimicrobial Peptide with High-Level, Broad-Spectrum Antimicrobial Potency and Skin Biocompatibility against Multidrug-Resistant Pseudomonas aeruginosa Infection. Antimicrob. Agents Chemother. 2018, 62, e01493-18. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, A.; Azmi, S.; Srivastava, S.; Kumar, A.; Tripathi, J.K.; Mishra, N.N.; Shukla, P.K.; Ghosh, J.K. Design and characterization of short antimicrobial peptides using leucine zipper templates with selectivity towards microorganisms. Amino Acids 2014, 46, 2531–2543. [Google Scholar] [CrossRef]
- Asthana, N.; Yadav, S.P.; Ghosh, J.K. Dissection of antibacterial and toxic activity of melittin: A leucine zipper motif plays a crucial role in determining its hemolytic activity but not antibacterial activity. J. Biol. Chem. 2004, 279, 55042–55050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, J.K.; Ovadia, M.; Shai, Y. A leucine zipper motif in the ectodomain of Sendai virus fusion assembles in solution and in membranes and specifically binds biologically-active peptides and the virus. Biochemistry 1997, 36, 15451–15462. [Google Scholar] [CrossRef] [PubMed]
- Chambers, P.; Pringle, C.R.; Easton, A.J. Heptad repeat sequences are located adjacent to hydrophobic regions in several types of virus fusion glycoproteins. J. Gen. Virol. 1990, 71 Pt 12, 3075–3080. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Kumari, T.; Tandon, A.; Sayeed, M.; Afshan, T.; Kathuria, M.; Shukla, P.K.; Mitra, K.; Ghosh, J.K. Selective phenylalanine to proline substitution for improved antimicrobial and anticancer activities of peptides designed on phenylalanine heptad repeat. Acta Biomater. 2017, 57, 170–186. [Google Scholar] [CrossRef] [PubMed]
- Omar, R.; Yadav, A. The remarkable cationic peptides: A boon to pharmaceutical sciences? J. Pharm. Pharm. Sci. 2018, 21, 60–72. [Google Scholar] [CrossRef] [PubMed]
- Thévenet, P.; Shen, Y.; Maupetit, J.; Guyon, F.; Derreumaux, P.; Tufféry, P. PEP-FOLD: An updated de novo structure prediction server for both linear and disulfide bonded cyclic peptides. Nucleic Acids Res. 2012, 40, W288–W293. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Maupetit, J.; Derreumaux, P.; Tufféry, P. Improved PEP-FOLD approach for peptide and miniprotein structure prediction. J. Chem. Theory Comput. 2014, 10, 4745–4758. [Google Scholar] [CrossRef]
- Eisenberg, D.; Weiss, R.M.; Terwilliger, T.C. The helical hydrophobic moment: A measure of the amphiphilicity of a helix. Nature 1982, 299, 371–374. [Google Scholar] [CrossRef]
- Terwilliger, T.C.; Weissman, L.; Eisenberg, D. The structure of melittin in the form I crystals and its implication for melittin’s lytic and surface activities. Biophys. J. 1982, 37, 353–361. [Google Scholar] [CrossRef] [Green Version]
- Park, S.C.; Kim, J.Y.; Shin, S.O.; Jeong, C.Y.; Kim, M.H.; Shin, S.Y.; Cheong, G.W.; Park, Y.; Hahm, K.S. Investigation of toroidal pore and oligomerization by melittin using transmission electron microscopy. Biochem. Biophys. Res. Commun. 2004, 321, 631–637. [Google Scholar] [CrossRef]
- Lee, J.K.; Park, S.C.; Hahm, K.S.; Park, Y. A helix-PXXP-helix peptide with antibacterial activity without cytotoxicity against MDRPA-infected mice. Biomaterials 2014, 35, 1025–1039. [Google Scholar] [CrossRef] [PubMed]
- Ajish, C.; Yang, S.; Kumar, S.D.; Shin, S.Y. Proadrenomedullin N-terminal 20 peptide (PAMP) and its C-terminal 12-residue peptide, PAMP(9-20): Cell selectivity and antimicrobial mechanism. Biochem. Biophys. Res. Commun. 2020, 527, 744–750. [Google Scholar] [CrossRef] [PubMed]
- Sharon, A.; Finkelstein, A.; Shlezinger, N.; Hatam, I. Fungal apoptosis: Function, genes and gene function. FEMS Microbiol. Rev. 2009, 33, 833–854. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Park, S.C.; Noh, G.; Kim, H.; Yoo, S.H.; Kim, I.R.; Lee, J.R.; Jang, M.K. Antifungal Effect of A Chimeric Peptide Hn-Mc against Pathogenic Fungal Strains. Antibiotics 2020, 9, 454. [Google Scholar] [CrossRef]
- Park, S.C.; Cheong, M.S.; Kim, E.J.; Kim, J.H.; Chi, Y.H.; Jang, M.K. Antifungal Effect of Arabidopsis SGT1 Proteins via Mitochondrial Reactive Oxygen Species. J. Agric. Food. Chem. 2017, 65, 8340–8347. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Calculated Mass | Observed Mass 1 | H2 | μH3 | RT (min) 4 | Net Charge |
|---|---|---|---|---|---|---|
| Pseudin-2 | 2682.5 | 2685.8 | 0.407 | 0.547 | 45.974 | +3 |
| P2-LZ1 | 2598.4 | 2601.8 | 0.287 | 0.430 | 43.920 | +3 |
| P2-LZ2 | 2564.5 | 2567.6 | 0.287 | 0.514 | 38.558 | +3 |
| P2-LZ3 | 2598.4 | 2601.4 | 0.283 | 0.428 | 43.549 | +3 |
| P2-LZ4 | 2564.5 | 2567.3 | 0.283 | 0.474 | 44.849 | +3 |
| P2-LZ5 | 2678.6 | 2681.2 | 0.175 | 0.382 | 33.234 | +5 |
| Fungi | MIC (µM) | ||||||
|---|---|---|---|---|---|---|---|
| Melittin | Pseudin-2 | P2-LZ1 | P2-LZ2 | P2-LZ3 | P2-LZ4 | P2-LZ5 | |
| Mold | |||||||
| A. flavus | 6 (4) | 64 (16) | 64 (12) | 32 (12) | 64 (32) | 32 (16) | 16 (>64) |
| A. fumigatus | 6 (4) | 64 (24) | 64 (16) | 24 (16) | 64 (16) | 32 (16) | 16 (>64) |
| F. moniliforme | 32 (32) | 32 (64) | 48 (48) | 24 (16) | 64 (32) | 32 (32) | 64 (>64) |
| F. oxysporum | 64 (32) | 4 (16) | 4 (12) | 2 (16) | 4 (16) | 2 (16) | 6 (16) |
| Yeast | |||||||
| C. albicans | 32 (8) | 12 (32) | 8 (32) | 4 (32) | 8 (16) | 4 (8) | 32 (64) |
| C. krusei | 8 (4) | 8 (8) | 8 (8) | 6 (4) | 8 (8) | 6 (8) | 48 (16) |
| C. parapsilosis | 16 (8) | >64 (>64) | > 64 (>64) | 48 (64) | >64 (>64) | 48 (>64) | >64 (>64) |
| C. tropicalis | 16 (8) | 4 (2) | 4 (2) | 2 (1.5) | 2 (2) | 2 (1) | 8 (16) |
| T. beigelii | 2 (2) | 2 (2) | 2 (2) | 2 (2) | 6 (2) | 1.5 (1) | 8 (32) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.-C.; Kim, H.; Kim, J.-Y.; Kim, H.; Cheong, G.-W.; Lee, J.R.; Jang, M.-K. Improved Cell Selectivity of Pseudin-2 via Substitution in the Leucine-Zipper Motif: In Vitro and In Vivo Antifungal Activity. Antibiotics 2020, 9, 921. https://doi.org/10.3390/antibiotics9120921
Park S-C, Kim H, Kim J-Y, Kim H, Cheong G-W, Lee JR, Jang M-K. Improved Cell Selectivity of Pseudin-2 via Substitution in the Leucine-Zipper Motif: In Vitro and In Vivo Antifungal Activity. Antibiotics. 2020; 9(12):921. https://doi.org/10.3390/antibiotics9120921
Chicago/Turabian StylePark, Seong-Cheol, Heabin Kim, Jin-Young Kim, Hyeonseok Kim, Gang-Won Cheong, Jung Ro Lee, and Mi-Kyeong Jang. 2020. "Improved Cell Selectivity of Pseudin-2 via Substitution in the Leucine-Zipper Motif: In Vitro and In Vivo Antifungal Activity" Antibiotics 9, no. 12: 921. https://doi.org/10.3390/antibiotics9120921
APA StylePark, S. -C., Kim, H., Kim, J. -Y., Kim, H., Cheong, G. -W., Lee, J. R., & Jang, M. -K. (2020). Improved Cell Selectivity of Pseudin-2 via Substitution in the Leucine-Zipper Motif: In Vitro and In Vivo Antifungal Activity. Antibiotics, 9(12), 921. https://doi.org/10.3390/antibiotics9120921