1. Introduction
Acinetobacter baumannii infection has become a research priority due to its exceptional ability to acquire antimicrobial resistance (AMR) [
1,
2]. Strains of this Gram negative bacterium have been shown to have high genetic diversity and large genetic ‘resistance islands’ [
3], and there have been cases of pan-drug resistant isolates that are resistant to all known clinical antibiotics [
4]. This has made
A. baumannii a formidable pathogen in the nosocomial environment, particularly against immunocompromised and intensive care unit (ICU) patients. Often targeting moist tissues such as mucous membranes [
5],
A. baumannii infection will commonly result in urinary tract infections, meningitis, pneumonia, and wound site infections. Forming strong biofilms,
A. baumannii is also commonly connected with the use of catheters and ventilators [
6], the latter of which is associated with particularly high mortality rates [
7].
Current therapies against
A. baumannii infections include the beta-lactamase inhibitor sulbactam, carbapenems, aminoglycosides, and tetracyclines [
8]. However, there is no longer an empirical antimicrobial agent treatment [
8] and the extensive use of the broad spectrum carbapenems [
9] has led the World Health Organization to highlight carbapenem resistant
A. baumannii as a Class 1: Priority Critical pathogen for the focus of research and development of novel antibiotics in 2017 [
1]. End line treatment options include tigecycline and colistin [
10], both of which have been associated with severe side effects, including hepatotoxicity, nephrotoxicity, and neurotoxicity [
6].
A. baumannii infections are of particular concern in Thailand [
11] where this pathogen is the most common cause of death by AMR associated nosocomial infection [
7]. According to NARST (National Antimicrobial Resistance Surveillance Centre, Thailand), 98% of
A. baumannii isolates were susceptible to the carbapenem imipenem in 1998 [
1,
12], but by 2002, this had dropped to 79%, and by 2008 only 43% of isolates screened were susceptible. Data collected for 2018 indicated that just over 32% of
A. baumannii isolates in Thailand were susceptible to imipenem and 31.8% to meropenem. In the same year, only 30.7% of 42,212 isolates were susceptible to sulbactam, 39.1% to the aminoglycoside gentamycin, and 18.5% to tetracycline [
12]. Currently, 97.1% of isolates are recorded to be susceptible to the last resort treatment, colistin. However, resistance genes to this antibiotic were discovered in 2011 [
13] and if the trends follow those seen for the carbapenems, the number of colistin susceptible
A. baumannii isolates could be reduced by two thirds within the next 20 years.
Bacteriophages are an attractive alternative to treating this drug resistant pathogen in the upcoming ‘post antibiotic era’ [
14]. The application of lytic bacteriophages could be prepared as a cocktail or as a combinatorial therapy, thereby reducing the chances of novel resistances being developed [
15]. Phage therapy [
16] has been shown to be successful in treating mouse models for
A. baumannii pneumonia [
17] and wound infections [
18], with the first human trial of bacteriophages being used against
A. baumannii published in 2017 [
19]. Gaining regulatory approval for phage therapy is complex and slow however, and as yet, there are no approved mainstream bacteriophage medicines in the EU or USA. In 2007 though, the Food and Drug Agency (FDA) approved the use of bacteriophages as a food additive to protect consumers against
Listeria monocytogenes in meat [
20], and over time, the regulation of phage therapy may become more straight forward. Until this time, bacteriophages still have the potential to be utilised for the benefit of human health in hand sanitizers, surface disinfectants, or antimicrobials [
21], and should not be overlooked as a powerful disease control tool. We are only just beginning to touch on the scope of how bacteriophages could be used in the pursuit of substantial human health benefits.
This study initially investigated five bacteriophages, isolated from hospital wastewater treatment plants in the lower northern Thai province of Phitsanulok, due to their good host range against 150 multidrug resistant (MDR)
A. baumannii isolates [
11,
22]. A preliminary investigation using 24-h growth curves indicated that bacteriophage vB_AbaM_PhT2 not only had the highest relative host range of those screened, but also maintained low
A. baumannii turbidity for the longest period of the five isolates during growth curves. This bacteriophage was therefore carried forward for a more detailed investigation, with the aim of potentially developing a surface disinfectant. Upon sequencing of the vB_AbaM_PhT2 genome, there were no known virulence factors or antibiotic resistance genes found. The treatment of human brain and bladder cell lines with this bacteriophage produced a significant reduction in relative cytotoxicity caused to the human cells by being grown in the presence of
A. baumannii. There was also an indication of vB_AbaM_PhT2 synergy with colistin, potentially allowing the clinical dosage of this nephrotoxic drug to be reduced. We therefore propose vB_AbaM_PhT2 as a candidate for future research and development into a tailored surface antimicrobial for use in hospitals.
3. Discussion
The five bacteriophages in this study had a host range of between 22% and 28% of 150 MDR
A. baumannii isolates. When analysing host range data and discussing the ‘broadness’ of the range, the diversity of location sources of the hosts should be taken into consideration [
40]. Bacteriophages isolated from hospital waste-water are more likely to be efficacious against clinical isolates from the same hospital or region than independent sources, due to coevolution [
28,
29]. The
A. baumannii isolates screened in this study were from five hospitals spread across Thailand, a country of over 500,000 km
2 with a population of around 70 million inhabitants [
41]. Hospitals were chosen to be representative of a range of different environments including industrial and rural areas, international borders, the presence of tourists and foreign workers, as well as geographic locations and regional climates. All five bacteriophages were from two hospitals in lower northern Thailand. Thus, the bacteriophages appear to have the potential to be used in a range of Thai hospital locations and possibly the rest of the world. In fact, a screen of the NCBI database [
33] showed vPht2 to be of the same species (% ANI > 95) as a bacteriophage isolated from Israel, AbTZA1 (accession number: MK278860). Groupings produced by the UPGMA dendogram, also revealed that there are two groups of bacteriophage host ranges within the five bacteriophages screened. If applied as a cocktail of cluster I (vPhT2, vPhT4, vPhT39) and II (vPhT29 and vPhT44), the overall host ranges could be expected to be higher [
29]. In fact, 60% (90 out of 150) of the clinical isolates were killed by at least one of the five bacteriophages.
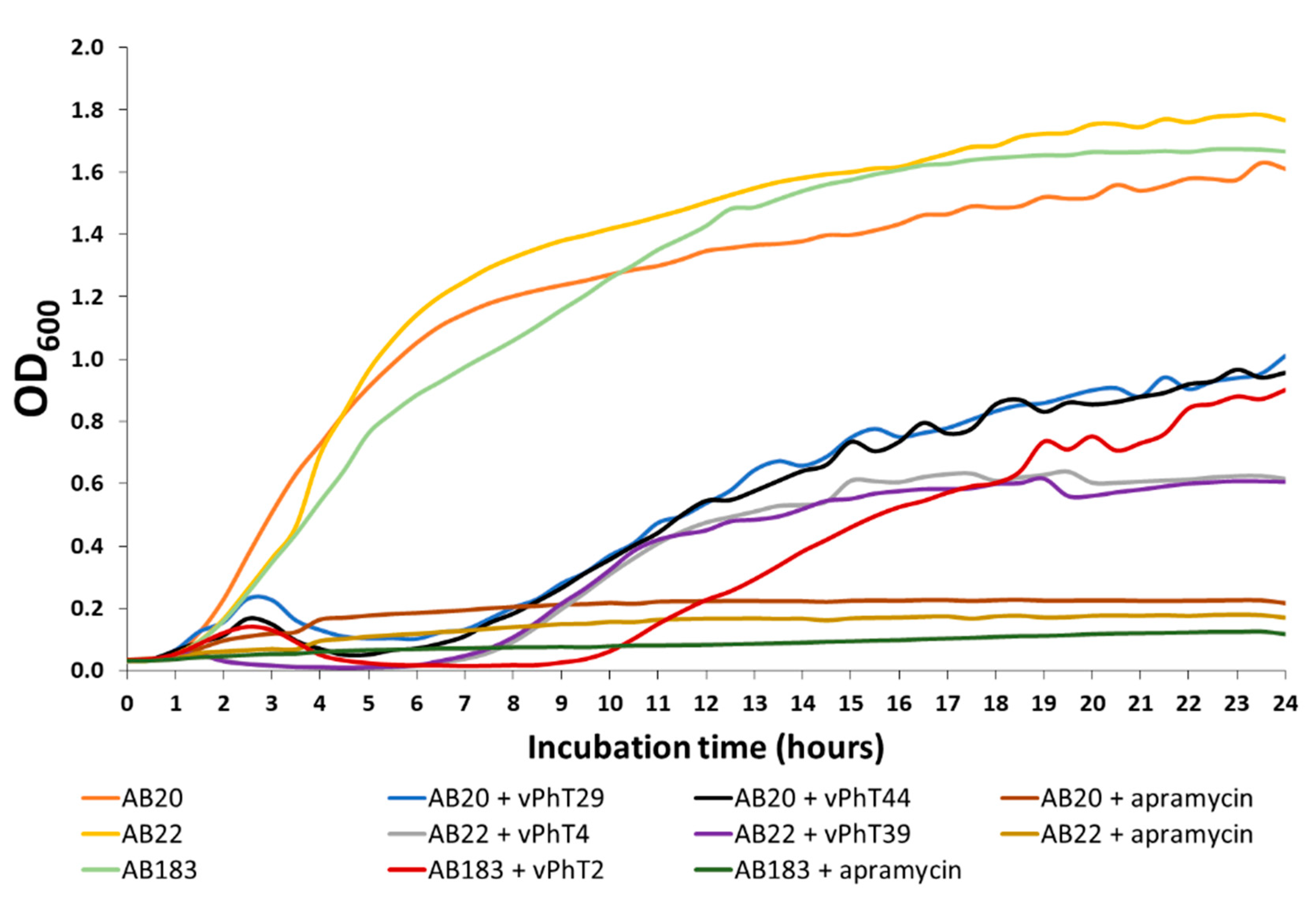
Over the course of a 24-h growth curve, the OD600 of samples treated with the bacteriophages did not increase past 60.84% of the negative controls. Although there was a significant reduction in host turbidity by all bacteriophages at all-time points analysed, bacteriophage vPhT4 produced the greatest relative reductions in A. baumannii metabolic activity (TTC assay) and turbidity (OD600), whereas bacteriophage vPhT2 maintained the low OD600 for the longest period (nine hours compared to 5.5–6.5 h for all others). Bacteriophage vPhT2 also had the highest host range and for this reason was selected for further investigation and characterisation.
The production of sequencing data on vPhT2 means there is now the option for the genetic engineering of this bacteriophage, which could potentially lead to the optimisation of its use for multiple commercial applications. An analysis of the vPhT2 genome showed that it contained the gene to carry a lysozyme domain fused to its tail, suggesting that this bacteriophage encodes enzymes capable of degrading host’s cell wall at the phage’s first contact with the cell surface and the last step of the phage lytic cycle [
42]. As the genome did not harbour any known virulence and antibiotic resistance genes that could potentially confer to a host, it shows promise as a suitable candidate for further applications in phage therapy.
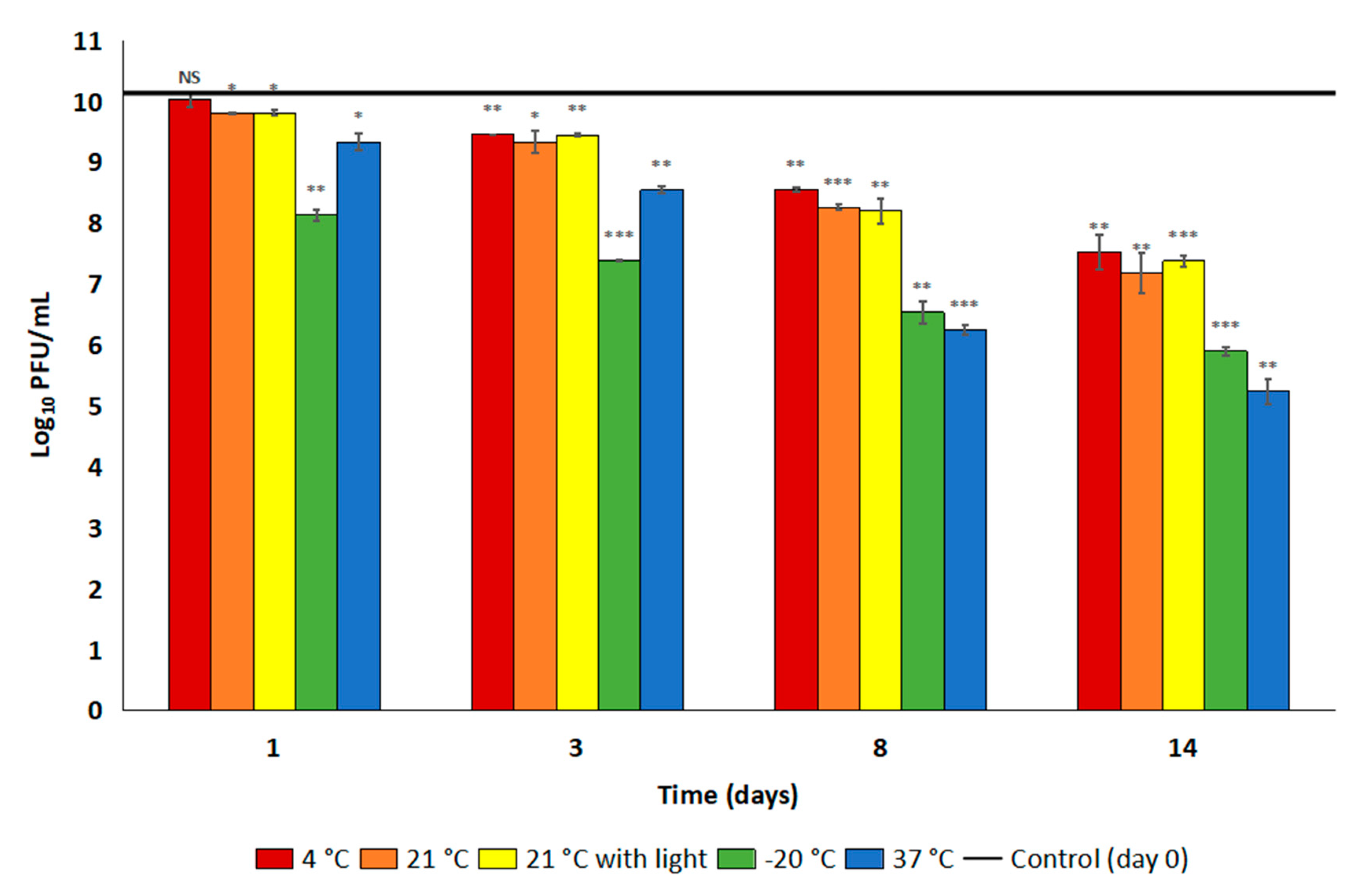
Stability and a sensible shelf-life are important for the accessibility and usefulness of an antibacterial product in the clinical environment. Any specific storage conditions could complicate existing hospital procedures and possibly lead to incorrect usage by untrained individuals. It is promising therefore that the vPhT2 titre was stable at room temperature, the most likely routine storage temperature for a disinfectant, when stored in LB (both in the presence and absence of light) for 14 days. After 10 months in the fridge in this same diluent, there was just over a one log reduction in vPhT2 titre, indicating that this is a possible long-term storage option. However, lysogeny broth is not a common commercial storage medium for bacteriophages and it is unclear why the bacteriophage was less stable in SM buffer II. Further investigation is therefore required to find a more suitable, commercially relevant diluent for vPhT2. We also suggest the use of an opaque storage vessel in order to protect the long-term efficacy of bacteriophage suspensions.
Endotoxins are part of the outer membrane of the cell wall of Gram negative bacteria and are a known pyrogenic contaminant of injectable drugs [
42]. Endotoxin levels for purified vPhT2 (1 × 10
8 PFU/mL) were shown to be below the threshold of 200 EU/mL as proposed by Brito et al. for endotoxin levels in live attenuated vaccines going through preclinical trials, based on United States Pharmacopoeia (USP) advice [
42,
43]. Should a higher dose or volume be required, endotoxin levels for dosages up to almost 20 times higher would still be below these guideline amounts and well within the USP recommendation of 5 EU/kg/hr [
44].
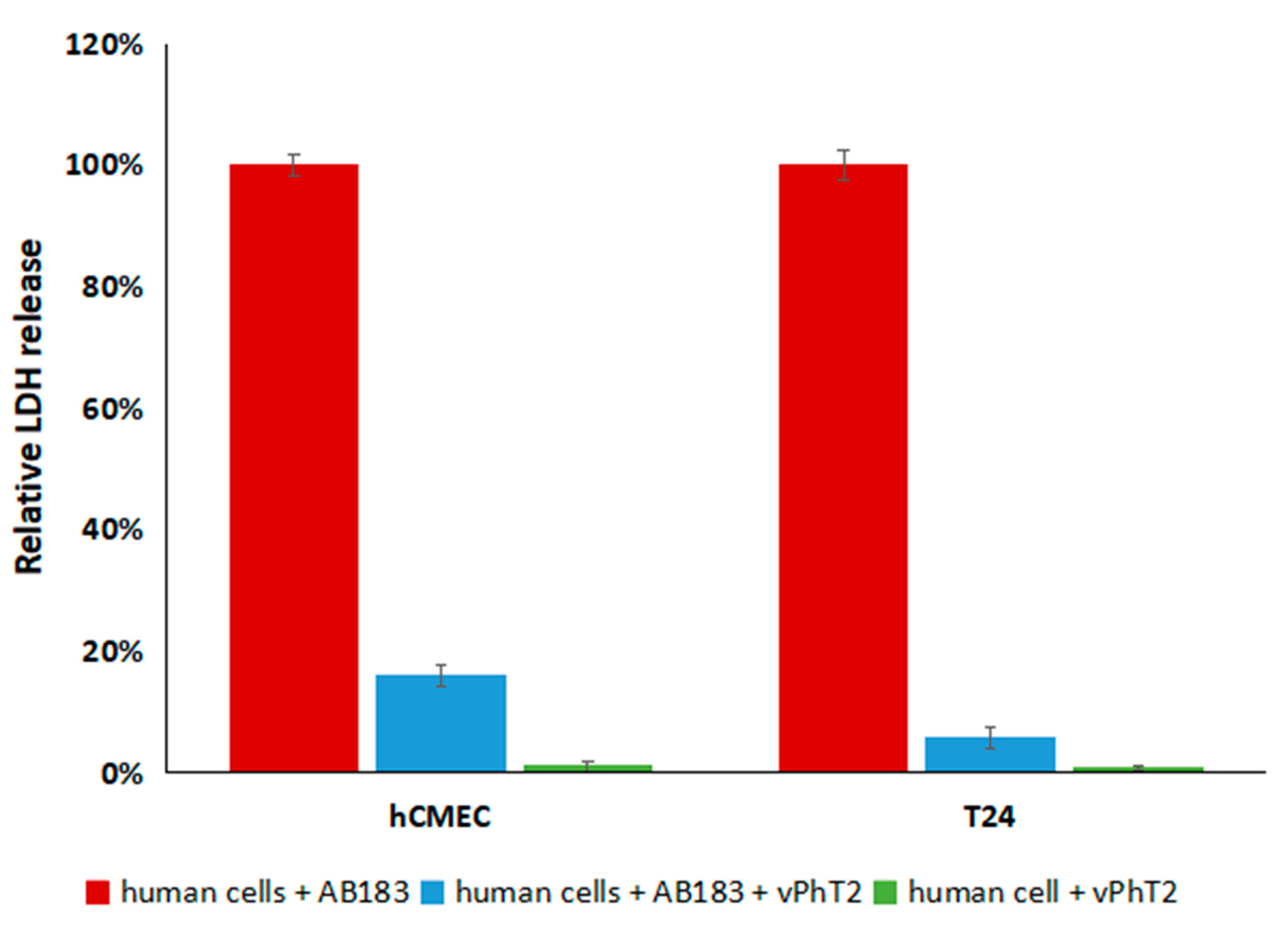
The treatment of human T24 bladder and hCMEC/D3 brain cell lines with A. baumannii and vPhT2 resulted in a significant reduction in cytotoxicity (as measured by relative LDH levels) compared to when no bacteriophage was present. In addition, neither human cell line grown in the presence of vPhT2 (with no A. baumannii) released significantly more LDH than controls grown without exposure to bacteriophages indicating that vPhT2 is not cytotoxic.
Previous trials with purified
A. baumannii bacteriophage endolysins showed broad range host activity and synergy with colistin [
27]. The polymyxin colistin, with all its associated side effects, is one of the end line treatments for
A. baumannii infection. With an FICI of 0.35, the indication that colistin can work synergistically with bacteriophage vPhT2 is very promising. Developing vPhT2 for use in the clinical environment, for example through developing an encapsulated delivery system [
45] or further stability trials are therefore to be considered as the next steps for bacteriophage vPhT2.

 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}