Metal–Peptide Complexes as Promising Antibiotics to Fight Emerging Drug Resistance: New Perspectives in Tuberculosis



Abstract
:1. Introduction
Antimicrobial Peptides
2. Metal-Based Antibiotic Therapeutic Agents
2.1. Antimicrobial Metal Complexes
2.2. Metal-Antimicrobial Peptides
2.3. Antibiotic Metal Complexes in Mtb Treatment
3. Delivery Systems for Antibacterial Compounds in Tb
4. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. World Health Organization global tuberculosis control report 2009. In Global Tuberculosis Control; World Health Organization: Geneva, Switzerland, 2011. [Google Scholar]
- Gandhi, N.R.; Nunn, P.; Dheda, K.; Schaaf, H.S.; Zignol, M.; Van Soolingen, D.; Jensen, P.; Bayona, J. Multidrug-resistant and extensively drug-resistant tuberculosis: A threat to global control of tuberculosis. Lancet 2010, 375, 1830–1843. [Google Scholar] [CrossRef]
- Maitra, A.; Munshi, T.; Healy, J.; Martin, L.T.; Vollmer, W.; Keep, N.H.; Bhakta, S. Cell wall peptidoglycan in Mycobacterium tuberculosis: An Achilles’ heel for the TB-causing pathogen. FEMS Microbiol. Rev. 2019, 43, 548–575. [Google Scholar] [CrossRef] [Green Version]
- Dulberger, C.L.; Rubin, E.J.; Boutte, C.C. The mycobacterial cell envelope–A moving target. Nat. Rev. Microbiol. 2019, 18, 1–13. [Google Scholar] [CrossRef]
- Smith, T.; Wolff, K.A.; Nguyen, L. Molecular biology of drug resistance in Mycobacterium tuberculosis. Curr. Top. Microbiol. Immunol. 2013, 374, 53–80. [Google Scholar]
- Rodriguez, C.A.; Papanastasiou, E.A.; Juba, M.; Bishop, B. Covalent modification of a ten-residue cationic antimicrobial peptide with levofloxacin. Front. Chem. 2014, 2, 71. [Google Scholar] [CrossRef]
- La Manna, S.; Di Natale, C.; Florio, D.; Marasco, D. Peptides as therapeutic agents for inflammatory-related diseases. Int. J. Mol. Sci. 2018, 19, 2714. [Google Scholar] [CrossRef] [Green Version]
- Di Somma, A.; Moretta, A.; Canè, C.; Cirillo, A.; Duilio, A. Antimicrobial and Antibiofilm Peptides. Biomolecules 2020, 10, 652. [Google Scholar] [CrossRef] [Green Version]
- Lazzaro, B.P.; Zasloff, M.; Rolff, J. Antimicrobial peptides: Application informed by evolution. Science 2020, 368, eaau5480. [Google Scholar] [CrossRef]
- Brown-Elliott, B.A.; Nash, K.A.; Wallace, R.J. Antimicrobial susceptibility testing, drug resistance mechanisms, and therapy of infections with nontuberculous mycobacteria. Clin. Microbiol. Rev. 2012, 25, 545–582. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Wu, H.; Lu, H.; Li, G.; Huang, Q. LAMP: A database linking antimicrobial peptides. PLoS ONE 2013, 8, e66557. [Google Scholar] [CrossRef] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef]
- Craik, D.J.; Fairlie, D.P.; Liras, S.; Price, D. The future of peptide-based drugs. Chem. Biol. Drug Design 2013, 81, 136–147. [Google Scholar] [CrossRef]
- Henderson, J.M.; Waring, A.J.; Separovic, F.; Lee, K.Y.C. Antimicrobial peptides share a common interaction driven by membrane line tension reduction. Biophys. J. 2016, 111, 2176–2189. [Google Scholar] [CrossRef] [Green Version]
- Morrison, G.M.; Semple, C.A.; Kilanowski, F.M.; Hill, R.E.; Dorin, J.R. Signal sequence conservation and mature peptide divergence within subgroups of the murine β-defensin gene family. Mol. Biol. Evol. 2003, 20, 460–470. [Google Scholar] [CrossRef] [Green Version]
- Semple, C.A.; Rolfe, M.; Dorin, J.R. Duplication and selection in the evolution of primate β-defensin genes. Genome Biol. 2003, 4, R31. [Google Scholar] [CrossRef] [Green Version]
- Van Harten, R.M.; Van Woudenbergh, E.; Van Dijk, A.; Haagsman, H.P. Cathelicidins: Immunomodulatory antimicrobials. Vaccines 2018, 6, 63. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, M. Cathelicidins, multifunctional peptides of the innate immunity. J. Leukoc. Biol. 2004, 75, 39–48. [Google Scholar] [CrossRef]
- Mookherjee, N.; Rehaume, L.M.; Hancock, R.E. Cathelicidins and functional analogues as antisepsis molecules. Expert Opin. Ther. Targets 2007, 11, 993–1004. [Google Scholar] [CrossRef]
- Shukla, P. Applied Microbiology and Bioengineering: An Interdisciplinary Approach; Academic Press: Cambridge, MA, USA, 2018. [Google Scholar]
- Yathursan, S.; Wiles, S.; Read, H.; Sarojini, V. A review on anti-tuberculosis peptides: Impact of peptide structure on anti-tuberculosis activity. J. Pept. Sci. 2019, 25, e3213. [Google Scholar] [CrossRef]
- Teng, T.; Li, X.; Zhang, L.; Li, Y. Identification and Characterization of pantocin wh-1, a Novel Cyclic Polypeptide Produced by Pantoea dispersa W18. Molecules 2020, 25, 485. [Google Scholar] [CrossRef] [Green Version]
- Ramón-García, S.; Mikut, R.; Ng, C.; Ruden, S.; Volkmer, R.; Reischl, M.; Hilpert, K.; Thompson, C.J. Targeting Mycobacterium tuberculosis and other microbial pathogens using improved synthetic antibacterial peptides. Antimicrob. Agents Chemother. 2013, 57, 2295–2303. [Google Scholar] [CrossRef] [Green Version]
- Silva, S.; Santos-Silva, A.; Da Costa, J.M.C.; Vale, N. Potent cationic antimicrobial peptides against Mycobacterium tuberculosis in vitro. J. Glob. Antimicrob. Resist. 2019, 19, 132–135. [Google Scholar] [CrossRef]
- Rodriguez, A.; Pedersen, M.Ø.; Villegas, E.; Rivas-Santiago, B.; Villegas-Moreno, J.; Amero, C.; Norton, R.S.; Corzo, G. Antimicrobial activity and structure of a consensus human β-defensin and its comparison to a novel putative hBD10. Proteins Struct. Funct. Bioinf. 2020, 88, 175–186. [Google Scholar] [CrossRef]
- Tenland, E.; Krishnan, N.; Rönnholm, A.; Kalsum, S.; Puthia, M.; Mörgelin, M.; Davoudi, M.; Otrocka, M.; Alaridah, N.; Glegola-Madejska, I. A novel derivative of the fungal antimicrobial peptide plectasin is active against Mycobacterium tuberculosis. Tuberculosis 2018, 113, 231–238. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Jiménez-Vargas, J.M.; Rivas-Santiago, B.; Corzo, G.; Possani, L.D.; Becerril, B.; Ortiz, E. Peptides from the scorpion Vaejovis punctatus with broad antimicrobial activity. Peptides 2015, 73, 51–59. [Google Scholar] [CrossRef]
- Kling, A.; Lukat, P.; Almeida, D.V.; Bauer, A.; Fontaine, E.; Sordello, S.; Zaburannyi, N.; Herrmann, J.; Wenzel, S.C.; König, C. Targeting DnaN for tuberculosis therapy using novel griselimycins. Science 2015, 348, 1106–1112. [Google Scholar] [CrossRef]
- Gavrish, E.; Sit, C.S.; Cao, S.; Kandror, O.; Spoering, A.; Peoples, A.; Ling, L.; Fetterman, A.; Hughes, D.; Bissell, A. Lassomycin, a ribosomally synthesized cyclic peptide, kills Mycobacterium tuberculosis by targeting the ATP-dependent protease ClpC1P1P2. Chem. Biol. 2014, 21, 509–518. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Pérez, C.; Gracia, B.; Rodrigues, L.; Vitoria, A.; Cebrián, R.; Deboosère, N.; Song, O.-R.; Brodin, P.; Maqueda, M.; Aínsa, J.A. Synergy between circular bacteriocin AS-48 and ethambutol against Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62, e00359-18. [Google Scholar] [CrossRef] [Green Version]
- Gupta, K.; Singh, S.; Van Hoek, M.L. Short, synthetic cationic peptides have antibacterial activity against Mycobacterium smegmatis by forming pores in membrane and synergizing with antibiotics. Antibiotics 2015, 4, 358–378. [Google Scholar] [CrossRef] [Green Version]
- Rao, M.; Ligeiro, D.; Maeurer, M. Precision medicine in the clinical management of respiratory tract infections including multidrug-resistant tuberculosis: Learning from innovations in immuno-oncology. Curr. Opin. Pulm. Med. 2019, 25, 233–241. [Google Scholar] [CrossRef]
- Ahmed, S.; Raqib, R.; Guðmundsson, G.H.; Bergman, P.; Agerberth, B.; Rekha, R.S. Host-Directed Therapy as a Novel Treatment Strategy to Overcome Tuberculosis: Targeting Immune Modulation. Antibiotics 2020, 9, 21. [Google Scholar] [CrossRef] [Green Version]
- Golpour, A.; Bereswill, S.; Heimesaat, M.M. Antimicrobial and immune-modulatory effects of vitamin D provide promising antibiotics-independent approaches to tackle bacterial infections–lessons learnt from a literature survey. Eur. J. Microbiol. Immunol. 2019, 9, 80–87. [Google Scholar] [CrossRef]
- Degiacomi, G.; Personne, Y.; Mondésert, G.; Ge, X.; Mandava, C.S.; Hartkoorn, R.C.; Boldrin, F.; Goel, P.; Peisker, K.; Benjak, A. Micrococcin P1–A bactericidal thiopeptide active against Mycobacterium tuberculosis. Tuberculosis 2016, 100, 95–101. [Google Scholar] [CrossRef]
- Ling, L.L.; Schneider, T.; Peoples, A.J.; Spoering, A.L.; Engels, I.; Conlon, B.P.; Mueller, A.; Schäberle, T.F.; Hughes, D.E.; Epstein, S. A new antibiotic kills pathogens without detectable resistance. Nature 2015, 517, 455–459. [Google Scholar] [CrossRef]
- Gao, W.; Kim, J.-Y.; Chen, S.-N.; Cho, S.-H.; Choi, J.; Jaki, B.U.; Jin, Y.-Y.; Lankin, D.C.; Lee, J.-E.; Lee, S.-Y. Discovery and characterization of the tuberculosis drug lead ecumicin. Org. Lett. 2014, 16, 6044–6047. [Google Scholar] [CrossRef] [Green Version]
- Choules, M.P.; Wolf, N.M.; Lee, H.; Anderson, J.R.; Grzelak, E.M.; Wang, Y.; Ma, R.; Gao, W.; McAlpine, J.B.; Jin, Y.-Y. Rufomycin targets ClpC1 proteolysis in Mycobacterium tuberculosis and M. abscessus. Antimicrob. Agents Chemother. 2019, 63, e02204-18. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.-G.; Lee, S.J.; Chae, S.; Lee, K.-Y.; Kim, J.-H.; Lee, B.-J. Structural and functional studies of the Mycobacterium tuberculosis VapBC30 toxin-antitoxin system: Implications for the design of novel antimicrobial peptides. Nucleic Acids Res. 2015, 43, 7624–7637. [Google Scholar] [CrossRef] [Green Version]
- Kwok, P.C.L.; Grabarek, A.; Chow, M.Y.; Lan, Y.; Li, J.C.; Casettari, L.; Mason, A.J.; Lam, J.K. Inhalable spray-dried formulation of D-LAK antimicrobial peptides targeting tuberculosis. Int. J. Pharm. 2015, 491, 367–374. [Google Scholar] [CrossRef]
- Chingaté, S.; Delgado, G.; Salazar, L.M.; Soto, C.-Y. The ATPase activity of the mycobacterial plasma membrane is inhibited by the LL37-analogous peptide LLAP. Peptides 2015, 71, 222–228. [Google Scholar] [CrossRef]
- Sharma, A.; Pohane, A.A.; Bansal, S.; Bajaj, A.; Jain, V.; Srivastava, A. Cell penetrating synthetic antimicrobial peptides (SAMPs) exhibiting potent and selective killing of Mycobacterium by targeting its DNA. Chem. A Eur. J. 2015, 21, 3540–3545. [Google Scholar] [CrossRef]
- Huante-Mendoza, A.; Silva-García, O.; Oviedo-Boyso, J.; Hancock, R.E.; Baizabal-Aguirre, V.M. Peptide IDR-1002 inhibits NF-κB nuclear translocation by inhibition of IκBα degradation and activates p38/ERK1/2-MSK1-dependent CREB phosphorylation in macrophages stimulated with lipopolysaccharide. Front. Immunol. 2016, 7, 533. [Google Scholar] [CrossRef] [Green Version]
- Mansour, S.C.; De la Fuente-Núñez, C.; Hancock, R.E. Peptide IDR-1018: Modulating the immune system and targeting bacterial biofilms to treat antibiotic-resistant bacterial infections. J. Pept. Sci. 2015, 21, 323–329. [Google Scholar] [CrossRef]
- Rivas-Santiago, B.; Castañeda-Delgado, J.E.; Santiago, C.E.R.; Waldbrook, M.; González-Curiel, I.; León-Contreras, J.C.; Enciso-Moreno, J.A.; Del Villar, V.; Mendez-Ramos, J.; Hancock, R.E. Ability of innate defence regulator peptides IDR-1002, IDR-HH2 and IDR-1018 to protect against Mycobacterium tuberculosis infections in animal models. PLoS ONE 2013, 8, e59119. [Google Scholar] [CrossRef] [Green Version]
- Arranz-Trullén, J.; Lu, L.; Pulido, D.; Bhakta, S.; Boix, E. Host antimicrobial peptides: The promise of new treatment strategies against tuberculosis. Front. Immunol. 2017, 8, 1499. [Google Scholar] [CrossRef] [Green Version]
- Pulido, D.; Torrent, M.; Andreu, D.; Nogués, M.V.; Boix, E. Two human host defense ribonucleases against mycobacteria, the eosinophil cationic protein (RNase 3) and RNase 7. Antimicrob. Agents Chemother. 2013, 57, 3797–3805. [Google Scholar] [CrossRef] [Green Version]
- Sharma, R.; Saikia, U.N.; Sharma, S.; Verma, I. Activity of human beta defensin-1 and its motif against active and dormant Mycobacterium tuberculosis. Appl. Microbiol. Biotechnol. 2017, 101, 7239–7248. [Google Scholar] [CrossRef]
- Barbie, P.; Kazmaier, U. Total synthesis of cyclomarin A, a marine cycloheptapeptide with anti-tuberculosis and anti-malaria activity. Org. Lett. 2016, 18, 204–207. [Google Scholar] [CrossRef]
- Kiefer, A.; Bader, C.D.; Held, J.; Esser, A.; Rybniker, J.; Empting, M.; Müller, R.; Kazmaier, U. Synthesis of new cyclomarin derivatives and their biological evaluation towards Mycobacterium tuberculosis and Plasmodium falciparum. Chem. A Eur. J. 2019, 25, 8894–8902. [Google Scholar] [CrossRef]
- Rodríguez, A.; Villegas, E.; Montoya-Rosales, A.; Rivas-Santiago, B.; Corzo, G. Characterization of antibacterial and hemolytic activity of synthetic pandinin 2 variants and their inhibition against Mycobacterium tuberculosis. PLoS ONE 2014, 9, e101742. [Google Scholar] [CrossRef]
- Florio, D.; Iacobucci, I.; Ferraro, G.; Mansour, A.M.; Morelli, G.; Monti, M.; Merlino, A.; Marasco, D. Role of the Metal Center in the Modulation of the Aggregation Process of Amyloid Model Systems by Square Planar Complexes Bearing 2-(2′-pyridyl) benzimidazole Ligands. Pharmaceuticals 2019, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Marasco, D.; Messori, L.; Marzo, T.; Merlino, A. Oxaliplatin vs. cisplatin: Competition experiments on their binding to lysozyme. Dalton Trans. 2015, 44, 10392–10398. [Google Scholar] [CrossRef] [Green Version]
- Florio, D.; Malfitano, A.M.; Di Somma, S.; Mügge, C.; Weigand, W.; Ferraro, G.; Iacobucci, I.; Monti, M.; Morelli, G.; Merlino, A. Platinum (II) O, S Complexes Inhibit the Aggregation of Amyloid Model Systems. Int. J. Mol. Sci. 2019, 20, 829. [Google Scholar] [CrossRef] [Green Version]
- Kanayama, Y.; Tsuji, T.; Enomoto, S.; Amano, R. Multitracer screening: Brain delivery of trace elements by eight different administration methods. Biometals 2005, 18, 553–565. [Google Scholar] [CrossRef]
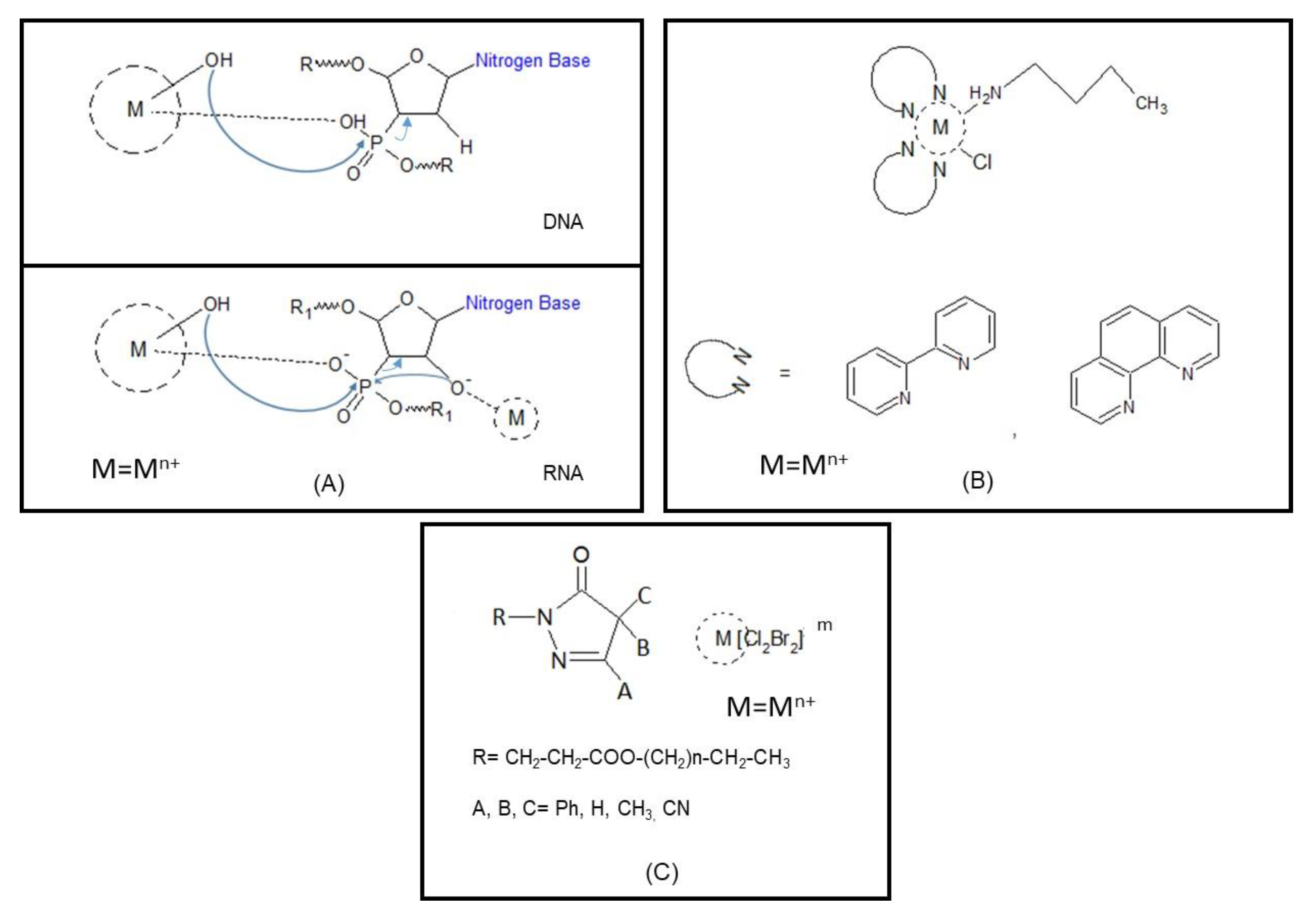
- Yu, Z.; Cowan, J.A. Metal complexes promoting catalytic cleavage of nucleic acids–Biochemical tools and therapeutics. Curr. Opin. Chem. Biol. 2018, 43, 37–42. [Google Scholar] [CrossRef]
- Lirussi, L.; Antoniali, G.; Scognamiglio, P.L.; Marasco, D.; Dalla, E.; D’Ambrosio, C.; Arena, S.; Scaloni, A.; Tell, G. Cleavage of the APE1 N-Terminal Domain in Acute Myeloid Leukemia Cells Is Associated with Proteasomal Activity. Biomolecules 2020, 10, 531. [Google Scholar] [CrossRef] [Green Version]
- Burra, S.; Marasco, D.; Malfatti, M.C.; Antoniali, G.; Virgilio, A.; Esposito, V.; Demple, B.; Galeone, A.; Tell, G. Human AP-endonuclease (Ape1) activity on telomeric G4 structures is modulated by acetylatable lysine residues in the N-terminal sequence. DNA Repair 2019, 73, 129–143. [Google Scholar] [CrossRef]
- Poletto, M.; Malfatti, M.C.; Dorjsuren, D.; Scognamiglio, P.L.; Marasco, D.; Vascotto, C.; Jadhav, A.; Maloney, D.J.; Wilson III, D.M.; Simeonov, A. Inhibitors of the apurinic/apyrimidinic endonuclease 1 (APE1)/nucleophosmin (NPM1) interaction that display anti-tumor properties. Mol. Carcinog. 2016, 55, 688–704. [Google Scholar] [CrossRef] [Green Version]
- Poletto, M.; Vascotto, C.; Scognamiglio, P.L.; Lirussi, L.; Marasco, D.; Tell, G. Role of the unstructured N-terminal domain of the hAPE1 (human apurinic/apyrimidinic endonuclease 1) in the modulation of its interaction with nucleic acids and NPM1 (nucleophosmin). Biochem. J. 2013, 452, 545–557. [Google Scholar] [CrossRef] [Green Version]
- Mancin, F.; Scrimin, P.; Tecilla, P.; Tonellato, U. Artificial metallonucleases. Chem. Commun. 2005, 20, 2540–2548. [Google Scholar] [CrossRef]
- Dupureur, C.M. Roles of metal ions in nucleases. Curr. Opin. Chem. Biol. 2008, 12, 250–255. [Google Scholar] [CrossRef]
- Baum, D.; Silverman, S.K. Deoxyribozymes: Useful DNA catalysts in vitro and in vivo. Cell. Mol. Life Sci. 2008, 65, 2156–2174. [Google Scholar] [CrossRef] [PubMed]
- Mancin, F.; Scrimin, P.; Tecilla, P. Progress in artificial metallonucleases. Chem. Commun. 2012, 48, 5545–5559. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Q.; Xiao, N.; Shi, P.; Zhu, Y.; Guo, Z. Design of artificial metallonucleases with oxidative mechanism. Coord. Chem. Rev. 2007, 251, 1951–1972. [Google Scholar] [CrossRef]
- Joyner, J.C.; Hodnick, W.; Cowan, A.S.; Tamuly, D.; Boyd, R.; Cowan, J. Antimicrobial metallopeptides with broad nuclease and ribonuclease activity. Chem. Commun. 2013, 49, 2118–2120. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Ji, H.; Liu, C.; Bing, W.; Wang, Z.; Qu, X. A Multinuclear Metal Complex Based DNase-Mimetic Artificial Enzyme: Matrix Cleavage for Combating Bacterial Biofilms. Angew. Chem. Int. Ed. 2016, 55, 10732–10736. [Google Scholar] [CrossRef]
- Schattschneider, C.; Kettenmann, S.D.; Hinojosa, S.; Heinrich, J.; Kulak, N. Biological activity of amphiphilic metal complexes. Coord. Chem. Rev. 2019, 385, 191–207. [Google Scholar] [CrossRef]
- Walker, G.W.; Geue, R.J.; Sargeson, A.M.; Behm, C.A. Surface-active cobalt cage complexes: Synthesis, surface chemistry, biological activity, and redox properties. Dalton Trans. 2003, 15, 2992–3001. [Google Scholar] [CrossRef]
- Kumar, R.S.; Arunachalam, S. Synthesis, micellar properties, DNA binding and antimicrobial studies of some surfactant–cobalt (III) complexes. Biophys. Chem. 2008, 136, 136–144. [Google Scholar] [CrossRef]
- Kaur, G.; Garg, P.; Kaur, B.; Chaudhary, G.R.; Kumar, S.; Dilbaghi, N.; Hassan, P.; Gawali, S.L. Cationic double chained metallosurfactants: Synthesis, aggregation, cytotoxicity, antimicrobial activity and their impact on the structure of bovine serum albumin. Soft Matter 2018, 14, 5306–5318. [Google Scholar] [CrossRef]
- Witthayapanyanon, A.; Phan, T.T.; Heitmann, T.C.; Harwell, J.H.; Sabatini, D.A. Interfacial properties of extended-surfactant-based microemulsions and related macroemulsions. J. Surfactants Deterg. 2010, 13, 127–134. [Google Scholar] [CrossRef]
- Negm, N.; Zaki, M. Structural and biological behaviors of some nonionic Schiff-base amphiphiles and their Cu (II) and Fe (III) metal complexes. Colloid. Surf. B Biointerfaces 2008, 64, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Pinkham, A.M.; Yu, Z.; Cowan, J. Attenuation of West Nile Virus NS2B/NS3 protease by amino terminal copper and nickel binding (ATCUN) peptides. J. Med. Chem. 2018, 61, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Hayden, R.M.; Goldberg, G.K.; Ferguson, B.M.; Schoeneck, M.W.; Libardo, M.D.J.; Mayeux, S.E.; Shrestha, A.; Bogardus, K.A.; Hammer, J.; Pryshchep, S. Complementary effects of host defense peptides piscidin 1 and piscidin 3 on DNA and lipid membranes: Biophysical insights into contrasting biological activities. J. Phys. Chem. B 2015, 119, 15235–15246. [Google Scholar] [CrossRef] [PubMed]
- Libardo, M.D.J.; Bahar, A.A.; Ma, B.; Fu, R.; McCormick, L.E.; Zhao, J.; McCallum, S.A.; Nussinov, R.; Ren, D.; Angeles-Boza, A.M. Nuclease activity gives an edge to host-defense peptide piscidin 3 over piscidin 1, rendering it more effective against persisters and biofilms. FEBS J. 2017, 284, 3662–3683. [Google Scholar] [CrossRef] [Green Version]
- Harford, C.; Sarkar, B. Amino terminal Cu (II)-and Ni (II)-binding (ATCUN) motif of proteins and peptides: Metal binding, DNA cleavage, and other properties. Acc. Chem. Res. 1997, 30, 123–130. [Google Scholar] [CrossRef]
- Rai, R.K.; De Angelis, A.; Greenwood, A.; Opella, S.J.; Cotten, M.L. Metal-ion Binding to Host Defense Peptide Piscidin 3 Observed in Phospholipid Bilayers by Magic Angle Spinning Solid-state NMR. ChemPhysChem 2019, 20, 295–301. [Google Scholar] [CrossRef]
- Oludiran, A.; Courson, D.S.; Stuart, M.D.; Radwan, A.R.; Poutsma, J.C.; Cotten, M.L.; Purcell, E.B. How Oxygen Availability Affects the Antimicrobial Efficacy of Host Defense Peptides: Lessons Learned from Studying the Copper-Binding Peptides Piscidins 1 and 3. Int. J. Mol. Sci. 2019, 20, 5289. [Google Scholar] [CrossRef] [Green Version]
- Dawson, R.M.; Liu, C.Q. Cathelicidin peptide SMAP-29: Comprehensive review of its properties and potential as a novel class of antibiotics. Drug Dev. Res. 2009, 70, 481–498. [Google Scholar] [CrossRef]
- Kalfa, V.; Jia, H.; Kunkle, R.; McCray, P.; Tack, B.; Brogden, K. Congeners of SMAP29 kill ovine pathogens and induce ultrastructural damage in bacterial cells. Antimicrob. Agents Chemother. 2001, 45, 3256–3261. [Google Scholar] [CrossRef] [Green Version]
- Saiman, L.; Tabibi, S.; Starner, T.D.; San Gabriel, P.; Winokur, P.L.; Jia, H.P.; McCray, P.B.; Tack, B.F. Cathelicidin peptides inhibit multiply antibiotic-resistant pathogens from patients with cystic fibrosis. Antimicrob. Agents Chemother. 2001, 45, 2838–2844. [Google Scholar] [CrossRef] [Green Version]
- Alexander, J.L.; Yu, Z.; Cowan, J. Amino terminal copper and nickel binding motif derivatives of ovispirin-3 display increased antimicrobial activity via lipid oxidation. J. Med. Chem. 2017, 60, 10047–10055. [Google Scholar] [CrossRef] [PubMed]
- Gusman, H.; Lendenmann, U.; Grogan, J.; Troxler, R.F.; Oppenheim, F.G. Is salivary histatin 5 a metallopeptide? Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 2001, 1545, 86–95. [Google Scholar] [CrossRef]
- Conklin, S.E.; Bridgman, E.C.; Su, Q.; Riggs-Gelasco, P.; Haas, K.L.; Franz, K.J. Specific histidine residues confer histatin peptides with copper-dependent activity against Candida albicans. Biochemistry 2017, 56, 4244–4255. [Google Scholar] [CrossRef] [PubMed]
- Kulprachakarn, K.; Chen, Y.-L.; Kong, X.; Arno, M.C.; Hider, R.C.; Srichairatanakool, S.; Bansal, S.S. Copper (II) binding properties of hepcidin. JBIC J. Biol. Inorg. Chem. 2016, 21, 329–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Barry, B.A. Ultraviolet resonance Raman microprobe spectroscopy of photosystem II. Photochem. Photobiol. 2008, 84, 815–818. [Google Scholar] [CrossRef]
- Abbas, I.M.; Vranic, M.; Hoffmann, H.; El-Khatib, A.H.; Montes-Bayón, M.; Möller, H.M.; Weller, M.G. Investigations of the Copper Peptide Hepcidin-25 by LC-MS/MS and NMR. Int. J. Mol. Sci. 2018, 19, 2271. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, P.; Bossak, K.; Stefaniak, E.; Hureau, C.; Raibaut, L.; Bal, W.; Faller, P. N-Terminal Cu-Binding Motifs (Xxx-Zzz-His, Xxx-His) and Their Derivatives: Chemistry, Biology and Medicinal Applications. Chem. A Eur. J. 2018, 24, 8029–8041. [Google Scholar] [CrossRef]
- Libardo, M.D.; Cervantes, J.L.; Salazar, J.C.; Angeles-Boza, A.M. Improved Bioactivity of Antimicrobial Peptides by Addition of Amino-Terminal Copper and Nickel (ATCUN) Binding Motifs. ChemMedChem 2014, 9, 1892–1901. [Google Scholar] [CrossRef] [Green Version]
- Park, C.B.; Yi, K.-S.; Matsuzaki, K.; Kim, M.S.; Kim, S.C. Structure-activity analysis of buforin II, a histone H2A-derived antimicrobial peptide: The proline hinge is responsible for the cell-penetrating ability of buforin II. Proc. Natl. Acad. Sci. USA 2000, 97, 8245–8250. [Google Scholar] [CrossRef] [Green Version]
- Hilpert, K.; Volkmer-Engert, R.; Walter, T.; Hancock, R.E. High-throughput generation of small antibacterial peptides with improved activity. Nat. Biotechnol. 2005, 23, 1008–1012. [Google Scholar] [CrossRef]
- Alexander, J.L.; Thompson, Z.; Yu, Z.; Cowan, J. Cu-ATCUN derivatives of Sub5 exhibit enhanced antimicrobial activity via multiple modes of action. ACS Chem. Biol. 2019, 14, 449–458. [Google Scholar] [CrossRef]
- Janicka-Kłos, A.; Janek, T.; Burger, J.; Czapor-Irzabek, H. Human salivary MUC7 mucin fragment and its analogues. Coordination and biological studies. J. Inorg. Biochem. 2020, 203, 110923. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Li, X.; Sun, G.; Zhang, Y.; Su, J.; Ye, J. Heavy Metal Induced Antibiotic Resistance in Bacterium LSJC7. Int. J. Mol. Sci. 2015, 16, 23390–23404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economou, N.J.; Cocklin, S.; Loll, P.J. High-resolution crystal structure reveals molecular details of target recognition by bacitracin. Proc. Natl. Acad. Sci. USA 2013, 110, 14207–14212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, K.W.; Ramesh, K.; Chew, F.T.; Mok, Y.K. Structural basis for the bacterial membrane insertion of dermcidin peptide, DCD-1L. Sci. Rep. UK 2017, 7, 1–11. [Google Scholar]
- Alexander, J.L.; Thompson, Z.; Cowan, J.A. Antimicrobial Metallopeptides. ACS Chem. Biol. 2018, 13, 844–853. [Google Scholar] [CrossRef]
- Paulmann, M.; Arnold, T.; Linke, D.; Özdirekcan, S.; Kopp, A.; Gutsmann, T.; Kalbacher, H.; Wanke, I.; Schuenemann, V.J.; Habeck, M. Structure-activity analysis of the dermcidin-derived peptide DCD-1L, an anionic antimicrobial peptide present in human sweat. J. Biol. Chem. 2012, 287, 8434–8443. [Google Scholar] [CrossRef] [Green Version]
- Juliano, S.A.; Pierce, S.; DeMayo, J.A.; Balunas, M.J.; Angeles-Boza, A.M. Exploration of the innate immune system of Styela clava: Zn2+ binding enhances the antimicrobial activity of the tunicate peptide clavanin A. Biochemistry 2017, 56, 1403–1414. [Google Scholar] [CrossRef]
- Hood, M.I.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2012, 10, 525–537. [Google Scholar] [CrossRef]
- Silva, F.D.; Rossi, D.C.; Martinez, L.R.; Frases, S.; Fonseca, F.L.; Campos, C.B.L.; Rodrigues, M.L.; Nosanchuk, J.D.; Daffre, S. Effects of microplusin, a copper-chelating antimicrobial peptide, against Cryptococcus neoformans. FEMS Microbiol. Lett. 2011, 324, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, D.M.; Brophy, M.B.; Bowman, S.E.; Stich, T.A.; Drennan, C.L.; Britt, R.D.; Nolan, E.M. Manganese binding properties of human calprotectin under conditions of high and low calcium: X-ray crystallographic and advanced electron paramagnetic resonance spectroscopic analysis. J. Am. Chem. Soc. 2015, 137, 3004–3016. [Google Scholar] [CrossRef] [PubMed]
- Rajput, S.; McLean, K.J.; Poddar, H.; Selvam, I.R.; Nagalingam, G.; Triccas, J.A.; Levy, C.W.; Munro, A.W.; Hutton, C.A. Structure-Activity Relationships of cyclo(l-Tyrosyl-l-tyrosine) Derivatives Binding to Mycobacterium tuberculosis CYP121: Iodinated Analogues Promote Shift to High-Spin Adduct. J. Med. Chem. 2019, 62, 9792–9805. [Google Scholar] [CrossRef] [PubMed]
- De Aguiar, I.; Tavares, A.; Roveda, A.C., Jr.; Da Silva, A.C.; Marino, L.B.; Lopes, É.O.; Pavan, F.R.; Lopes, L.G.; Franco, D.W. Antitubercular activity of Ru (II) isoniazid complexes. Eur. J. Pharm. Sci. 2015, 70, 45–54. [Google Scholar] [CrossRef] [PubMed]
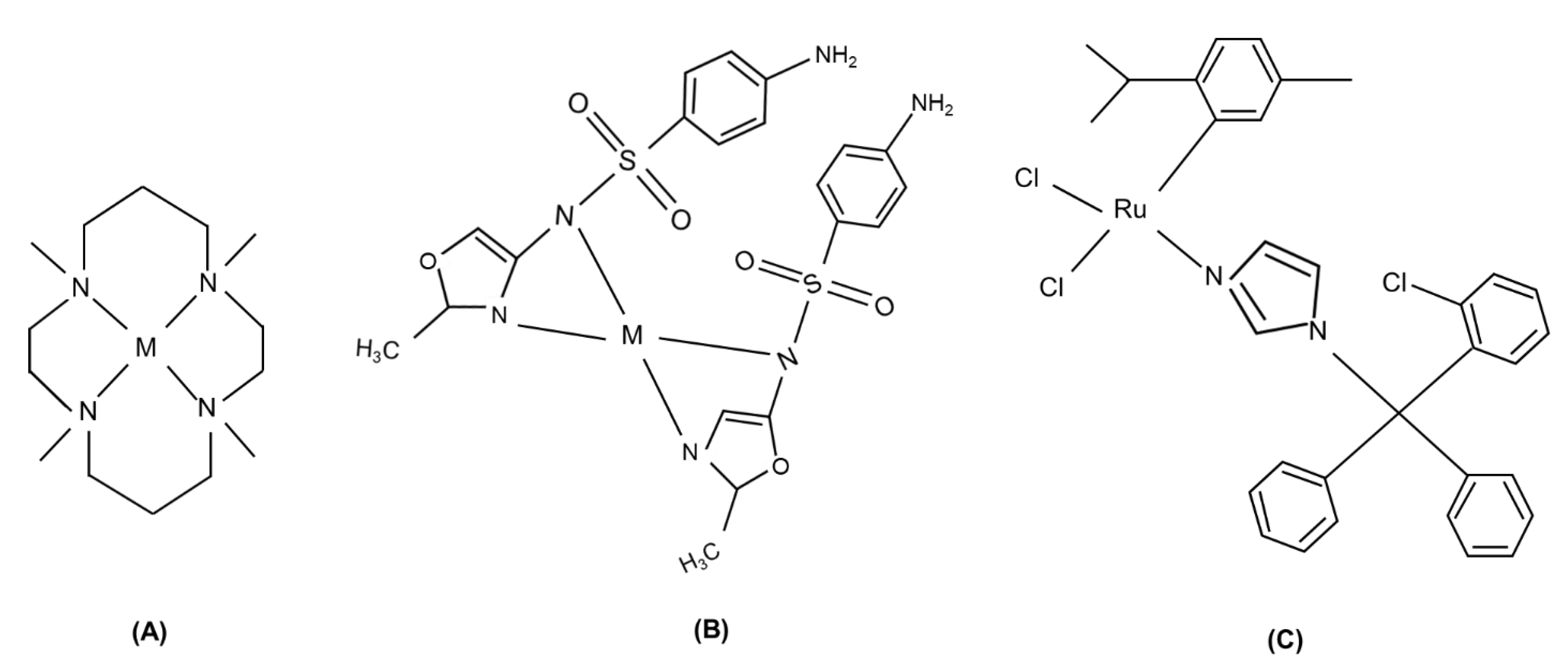
- Yu, M.; Nagalingam, G.; Ellis, S.; Martinez, E.; Sintchenko, V.; Spain, M.; Rutledge, P.J.; Todd, M.H.; Triccas, J.A. Nontoxic metal-cyclam complexes, a new class of compounds with potency against drug-resistant Mycobacterium tuberculosis. J. Med. Chem. 2016, 59, 5917–5921. [Google Scholar] [CrossRef] [PubMed]
- Frost, S.; McKenna, R. Carbonic Anhydrase: Mechanism, Regulation, Links to Disease, and Industrial Applications; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Chohan, Z.H.; Shad, H.A.; Youssoufi, M.H.; Hadda, T.B. Some new biologically active metal-based sulfonamide. Eur. J. Med. Chem. 2010, 45, 2893–2901. [Google Scholar] [CrossRef]
- Pavan, F.R.; Von Poelhsitz, G.; Do Nascimento, F.B.; Leite, S.R.; Batista, A.A.; Deflon, V.M.; Sato, D.N.; Franzblau, S.G.; Leite, C.Q. Ruthenium (II) phosphine/picolinate complexes as antimycobacterial agents. Eur. J. Med. Chem. 2010, 45, 598–601. [Google Scholar] [CrossRef]
- Frei, A. Metal Complexes, an untapped Source of Antibiotic Potential? Antibiotics 2020, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Chellan, P.; Avery, V.M.; Duffy, S.; Triccas, J.A.; Nagalingam, G.; Tam, C.; Cheng, L.W.; Liu, J.; Land, K.M.; Clarkson, G.J.; et al. Organometallic Conjugates of the Drug Sulfadoxine for Combatting Antimicrobial Resistance. Chemistry 2018, 24, 10078–10090. [Google Scholar] [CrossRef] [Green Version]
- Nakahata, D.H.; De Paiva, R.E.; Lustri, W.R.; Ribeiro, C.M.; Pavan, F.R.; Da Silva, G.G.; Ruiz, A.L.; De Carvalho, J.E.; Corbi, P.P. Sulfonamide-containing copper (II) metallonucleases: Correlations with in vitro antimycobacterial and antiproliferative activities. J. Inorg. Biochem. 2018, 187, 85–96. [Google Scholar] [CrossRef] [Green Version]
- Forgacs, P.; Wengenack, N.L.; Hall, L.; Zimmerman, S.K.; Silverman, M.L.; Roberts, G.D. Tuberculosis and trimethoprim-sulfamethoxazole. Antimicrob. Agents Chemother. 2009, 53, 4789–4793. [Google Scholar] [CrossRef] [Green Version]
- Corcoran, J.; Lange, A.; Cumming, R.I.; Owen, S.F.; Ball, J.S.; Tyler, C.R.; Winter, M.J. Bioavailability of the imidazole antifungal agent clotrimazole and its effects on key biotransformation genes in the common carp (Cyprinus carpio). Aquat. Toxicol. 2014, 152, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Abid, M.; Shamsi, F.; Azam, A. Ruthenium complexes: An emerging ground to the development of metallopharmaceuticals for cancer therapy. Mini Rev. Med. Chem. 2016, 16, 772–786. [Google Scholar] [CrossRef] [PubMed]
- Colina-Vegas, L.; Dutra, J.L.; Villarreal, W.; Neto, J.H.d.A.; Cominetti, M.R.; Pavan, F.; Navarro, M.; Batista, A.A. Ru (II)/clotrimazole/diphenylphosphine/bipyridine complexes: Interaction with DNA, BSA and biological potential against tumor cell lines and Mycobacterium tuberculosis. J. Inorg. Biochem. 2016, 162, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva, J.P.; Silva, I.C.; Pavan, F.R.; Back, D.F.; De Araujo, M.P. Bis (diphenylphosphino) amines-containing ruthenium cymene complexes as potential anti-Mycobacterium tuberculosis agents. J. Inorg. Biochem. 2017, 173, 134–140. [Google Scholar] [CrossRef]
- Paixão, D.A.; Marzano, I.M.; Jaimes, E.H.; Pivatto, M.; Campos, D.L.; Pavan, F.R.; Deflon, V.M.; Maia, P.I.d.S.; Ferreira, A.M.D.C.; Uehara, I.A. Novel copper (II) complexes with hydrazides and heterocyclic bases: Synthesis, structure and biological studies. J. Inorg. Biochem. 2017, 172, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, A.E.; DeStefano, M.; Shoen, C.; Gopinath, K.; Warner, D.F.; Cynamon, M.; Doyle, R.P. Co (II) and Cu (II) pyrophosphate complexes have selectivity and potency against Mycobacteria including Mycobacterium tuberculosis. Eur. J. Med. Chem. 2013, 70, 589–593. [Google Scholar] [CrossRef] [PubMed]
- Do Couto Almeida, J.; Marzano, I.M.; Pivatto, M.; Lopes, N.P.; Ferreira, A.M.D.C.; Pavan, F.; Silva, I.; Pereira-Maia, E.C.; Von Poelhsitz, G.; Guerra, W. Synthesis, cytotoxic and antitubercular activities of copper (II) complexes with heterocyclic bases and 3-hydroxypicolinic acid. Inorg. Chim. Acta 2016, 446, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Shah, S.; Dalecki, A.G.; Malalasekera, A.P.; Crawford, C.L.; Michalek, S.M.; Kutsch, O.; Sun, J.; Bossmann, S.H.; Wolschendorf, F. 8-Hydroxyquinolines are boosting agents of copper-related toxicity in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2016, 60, 5765–5776. [Google Scholar] [CrossRef] [Green Version]
- Jeżowska-Bojczuk, M.; Stokowa-Sołtys, K. Peptides having antimicrobial activity and their complexes with transition metal ions. Eur. J. Med. Chem. 2018, 143, 997–1009. [Google Scholar] [CrossRef]
- Stokowa-Sołtys, K.; Barbosa, N.A.; Kasprowicz, A.; Wieczorek, R.; Gaggelli, N.; Gaggelli, E.; Valensin, G.; Wrzesiński, J.; Ciesiołka, J.; Kuliński, T. Studies of viomycin, an anti-tuberculosis antibiotic: Copper (ii) coordination, DNA degradation and the impact on delta ribozyme cleavage activity. Dalton Trans. 2016, 45, 8645–8658. [Google Scholar] [CrossRef] [Green Version]
- Smieja, M.; Marchetti, C.; Cook, D.; Smaill, F.M. Isoniazid for preventing tuberculosis in non-HIV infected persons. Cochrane Database Syst. Rev. 1999, 2, CD001363. [Google Scholar] [CrossRef] [PubMed]
- Firmino, G.S.; De Souza, M.V.; Pessoa, C.; Lourenco, M.C.; Resende, J.A.; Lessa, J.A. Synthesis and evaluation of copper (II) complexes with isoniazid-derived hydrazones as anticancer and antitubercular agents. BioMetals 2016, 29, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Rooney, D.; McCann, M.; Devereux, M.; Twamley, B.; Galdino, A.C.M.; Sangenito, L.S.; Souza, L.O.P.; Lourenço, M.C.; Gomes, K. Synthesis and antimicrobial activity of a phenanthroline-isoniazid hybrid ligand and its Ag+ and Mn2+ complexes. BioMetals 2019, 32, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Coelho, T.S.; Halicki, P.C.B.; Silva, L., Jr.; De Menezes Vicenti, J.R.; Goncalves, B.L.; Almeida da Silva, P.E.; Ramos, D.F. Metal-based antimicrobial strategies against intramacrophage Mycobacterium tuberculosis. Lett. Appl. Microbiol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Giovagnoli, S.; Marenzoni, M.L.; Nocchetti, M.; Santi, C.; Blasi, P.; Schoubben, A.; Ricci, M. Synthesis, characterization and in vitro extracellular and intracellular activity against M ycobacterium tuberculosis infection of new second-line antitubercular drug-palladium complexes. J. Pharm. Pharmacol. 2014, 66, 106–121. [Google Scholar] [CrossRef]
- Singh, A.; Lumb, I.; Mehra, V.; Kumar, V. Ferrocene-appended pharmacophores: An exciting approach for modulating the biological potential of organic scaffolds. Dalton Trans. 2019, 48, 2840–2860. [Google Scholar] [CrossRef]
- Zhang, J.; Bond, A.M. Conditions required to achieve the apparent equivalence of adhered solid-and solution-phase voltammetry for ferrocene and other redox-active solids in ionic liquids. Anal. Chem. 2003, 75, 2694–2702. [Google Scholar] [CrossRef]
- Stringer, T.; Seldon, R.; Liu, N.; Warner, D.F.; Tam, C.; Cheng, L.W.; Land, K.M.; Smith, P.J.; Chibale, K.; Smith, G.S. Antimicrobial activity of organometallic isonicotinyl and pyrazinyl ferrocenyl-derived complexes. Dalton Trans. 2017, 46, 9875–9885. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.C.; Carbone, C.; Sousa, M.C.; Espina, M.; Garcia, M.L.; Sanchez-Lopez, E.; Souto, E.B. Nanomedicines for the Delivery of Antimicrobial Peptides (AMPs). Nanomaterials 2020, 10, 560. [Google Scholar] [CrossRef] [Green Version]
- Makvandi, P.; Wang, C.Y.; Zare, E.N.; Borzacchiello, A.; Niu, L.N.; Tay, F.R. Metal–Based Nanomaterials in Biomedical Applications: Antimicrobial Activity and Cytotoxicity Aspects. Adv. Funct. Mater. 2020, 30, 1910021. [Google Scholar] [CrossRef]
- Manning, T.; Mikula, R.; Lee, H.; Calvin, A.; Darrah, J.; Wylie, G.; Phillips, D.; Bythell, B.J. The copper (II) ion as a carrier for the antibiotic capreomycin against Mycobacterium tuberculosis. Bioorg. Med. Chem. Lett. 2014, 24, 976–982. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bonilla, A.; Fernández-García, M. Polymeric materials with antimicrobial activity. Prog. Polym. Sci. 2012, 37, 281–339. [Google Scholar] [CrossRef]
- Wang, C.-Y.; Makvandi, P.; Zare, E.N.; Tay, F.R.; Niu, L.-N. Advances in Antimicrobial Organic and Inorganic Nanocompounds in Biomedicine. Adv. Ther. 2020, in press. [Google Scholar] [CrossRef]
- Makvandi, P.; Gu, J.T.; Zare, E.N.; Ashtari, B.; Moeini, A.; Tay, F.R.; Niu, L.-n. Polymeric and inorganic nanoscopical antimicrobial fillers in dentistry. Acta Biomater. 2020, 101, 69–101. [Google Scholar] [CrossRef] [PubMed]
- Martin-Serrano, Á.; Gómez, R.; Ortega, P.; De la Mata, F.J. Nanosystems as vehicles for the delivery of antimicrobial peptides (AMPs). Pharmaceutics 2019, 11, 448. [Google Scholar] [CrossRef] [Green Version]
- Jamaledin, R.; Di Natale, C.; Onesto, V.; Taraghdari, Z.B.; Zare, E.N.; Makvandi, P.; Vecchione, R.; Netti, P.A. Progress in Microneedle-Mediated Protein Delivery. J. Clin. Med. 2020, 9, 542. [Google Scholar] [CrossRef] [Green Version]
- De Almeida Pachioni-Vasconcelos, J.; Lopes, A.M.; Apolinário, A.C.; Valenzuela-Oses, J.K.; Costa, J.S.R.; De Oliveira Nascimento, L.; Pessoa, A.; Barbosa, L.R.S.; De Oliveira Rangel-Yagui, C. Nanostructures for protein drug delivery. Biomater. Sci. 2016, 4, 205–218. [Google Scholar] [CrossRef]
- Garcia-Orue, I.; Gainza, G.; Girbau, C.; Alonso, R.; Aguirre, J.J.; Pedraz, J.L.; Igartua, M.; Hernandez, R.M. LL37 loaded nanostructured lipid carriers (NLC): A new strategy for the topical treatment of chronic wounds. Eur. J. Pharm. Biopharm. 2016, 108, 310–316. [Google Scholar] [CrossRef]
- Chen, G.; Zhou, M.; Chen, S.; Lv, G.; Yao, J. Nanolayer biofilm coated on magnetic nanoparticles by using a dielectric barrier discharge glow plasma fluidized bed for immobilizing an antimicrobial peptide. Nanotechnology 2009, 20, 465706. [Google Scholar] [CrossRef]
- Dror, Y.; Ophir, C.; Freeman, A. Silver–enzyme hybrids as wide-spectrum antimicrobial agents. In Innovations and Emerging Technologies in Wound Care; Elsevier: Amsterdam, The Netherlands, 2020; pp. 293–307. [Google Scholar]
- Chen, W.Y.; Chang, H.Y.; Lu, J.K.; Huang, Y.C.; Harroun, S.G.; Tseng, Y.T.; Li, Y.J.; Huang, C.C.; Chang, H.T. Self-assembly of antimicrobial peptides on gold nanodots: Against multidrug-resistant bacteria and wound-healing application. Adv. Funct. Mater. 2015, 25, 7189–7199. [Google Scholar] [CrossRef]
- Zheng, K.; Setyawati, M.I.; Lim, T.-P.; Leong, D.T.; Xie, J. Antimicrobial cluster bombs: Silver nanoclusters packed with daptomycin. ACS Nano 2016, 10, 7934–7942. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Hu, C.; Shao, L. The antimicrobial activity of nanoparticles: Present situation and prospects for the future. Int. J. Nanomed. 2017, 12, 1227–1249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camila Micheletti, A.; Kika Honda, N.; R Pavan, F.; QF Leite, C.; De Fatima Cepa Matos, M.; Trentin Perdomo, R.; Bogo, D.; Braz Alcantara, G.; Beatriz, A. Increment of antimycobaterial activity on lichexanthone derivatives. Med. Chem. 2013, 9, 904–910. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, E.R.; Mondelli, M.A.; Pozzi, L.V.; Corrêa, R.S.; Salistre-de-Araújo, H.S.; Pavan, F.R.; Leite, C.Q.; Ellena, J.; Malta, V.R.; Machado, S.P. New ruthenium (II)/phosphines/diimines complexes: Promising antitumor (human breast cancer) and Mycobacterium tuberculosis fighting agents. Polyhedron 2013, 51, 292–297. [Google Scholar] [CrossRef]
- Santos, J.A.d.; Corrales, R.C.N.R.; Pavan, F.R.; Leite, C.Q.F.; Dias, R.M.d.P.; Silva, A.D.d. Synthesis and antitubercular evaluation of new 1, 2, 3-triazole derivatives of carbohydrates. Mediterr. J. Chem. 2012, 1, 281–288. [Google Scholar] [CrossRef]
- Pereira, G.A.; Massabni, A.C.; Castellano, E.E.; Costa, L.A.S.; Leite, C.Q.F.; Pavan, F.R.; Cuin, A. A broad study of two new promising antimycobacterial drugs: Ag (I) and Au (I) complexes with 2-(2-thienyl) benzothiazole. Polyhedron 2012, 38, 291–296. [Google Scholar] [CrossRef]
- Mondelli, M.; Pavan, F.; De Souza, P.C.; Leite, C.Q.; Ellena, J.; Nascimento, O.R.; Facchin, G.; Torre, M.H. Study of a series of cobalt (II) sulfonamide complexes: Synthesis, spectroscopic characterization, and microbiological evaluation against M. tuberculosis. Crystal structure of [Co (sulfamethoxazole) 2 (H2O) 2]·H2O. J. Mol. Struct. 2013, 1036, 180–187. [Google Scholar] [CrossRef]
- Pavan, F.R.; Poelhsitz, G.V.; Da Cunha, L.V.; Barbosa, M.I.; Leite, S.R.; Batista, A.A.; Cho, S.H.; Franzblau, S.G.; De Camargo, M.S.; Resende, F.A. In vitro and in vivo activities of ruthenium (II) phosphine/diimine/picolinate complexes (SCAR) against Mycobacterium tuberculosis. PLoS ONE 2013, 8, 642428. [Google Scholar] [CrossRef]
- Pavan, F.R.; Poelhsitz, G.V.; Barbosa, M.I.; Leite, S.R.; Batista, A.A.; Ellena, J.; Sato, L.S.; Franzblau, S.G.; Moreno, V.; Gambino, D. Ruthenium (II) phosphine/diimine/picolinate complexes: Inorganic compounds as agents against tuberculosis. Eur. J. Med. Chem. 2011, 46, 5099–5107. [Google Scholar] [CrossRef]
- Mehta, S.; Kaur, G.; Bhasin, K. Tween-embedded microemulsions–Physicochemical and spectroscopic analysis for antitubercular drugs. AAPS PharmSciTech 2010, 11, 143–153. [Google Scholar] [CrossRef]
- Jeetah, R.; Bhaw-Luximon, A.; Jhurry, D. Dual encapsulation and controlled delivery of anti-TB drugs from PEG-block-poly (ester-ether) nanomicelles. J. Nanopharm. Drug Deliv. 2013, 1, 240–257. [Google Scholar] [CrossRef]
- Choi, S.-R.; Britigan, B.E.; Moran, D.M.; Narayanasamy, P. Gallium nanoparticles facilitate phagosome maturation and inhibit growth of virulent Mycobacterium tuberculosis in macrophages. PLoS ONE 2017, 12, e0177987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Praphakar, R.A.; Jeyaraj, M.; Ahmed, M.; Kumar, S.S.; Rajan, M. Silver nanoparticle functionalized CS-g-(CA-MA-PZA) carrier for sustainable anti-tuberculosis drug delivery. Int. J. Biol. Macromol. 2018, 118, 1627–1638. [Google Scholar] [CrossRef] [PubMed]
- Zare, E.N.; Jamaledin, R.; Naserzadeh, P.; Afjeh-Dana, E.; Ashtari, B.; Hosseinzadeh, M.; Vecchione, R.; Wu, A.; Tay, F.R.; Borzacchiello, A. Metal-Based Nanostructures/PLGA Nanocomposites: Antimicrobial Activity, Cytotoxicity, and Their Biomedical Applications. ACS Appl. Mater. Interfaces 2020, 12, 3279–3300. [Google Scholar] [CrossRef] [PubMed]
- Pereira, E.D.; Souza, F.G., Jr.; Pinto, J.C.C.; Cerruti, R.; Santana, C. Synthesis, Characterization and Drug Delivery Profile of Magnetic PLGA-PEG-PLGA/Maghemite Nanocomposite; Macromolecular Symposia; Wiley Online Library: Hoboken, NJ, USA, 2014; pp. 18–25. [Google Scholar]
- Gajendiran, M.; Yousuf, S.M.J.; Elangovan, V.; Balasubramanian, S. Gold nanoparticle conjugated PLGA-PEG-SA-PEG-PLGA multiblock copolymer nanoparticles: Synthesis, characterization, in vivo release of rifampicin. J. Mater. Chem. B 2014, 2, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Manning, T.; Mikula, R.; Wylie, G.; Phillips, D.; Jarvis, J.; Zhang, F. Structural measurements and cell line studies of the copper-PEG-Rifampicin complex against Mycobacterium tuberculosis. Bioorg. Med. Chem. Lett. 2015, 25, 451–458. [Google Scholar] [CrossRef]
- Ellis, T.; Chiappi, M.; García-Trenco, A.; Al-Ejji, M.; Sarkar, S.; Georgiou, T.K.; Shaffer, M.S.; Tetley, T.D.; Schwander, S.; Ryan, M.P. Multimetallic microparticles increase the potency of rifampicin against intracellular Mycobacterium tuberculosis. ACS Nano 2018, 12, 5228–5240. [Google Scholar] [CrossRef]
- Jahagirdar, P.S.; Gupta, P.K.; Kulkarni, S.P.; Devarajan, P.V. Intramacrophage delivery of dual drug loaded nanoparticles for effective clearance of Mycobacterium tuberculosis. J. Pharm. Sci. 2020, 20, 30177-5. [Google Scholar] [CrossRef]
- Zhao, C.; Liu, X.; Zhang, X.; Yan, H.; Qian, Z.; Li, X.; Ma, Z.; Han, Q.; Pei, C. A facile one-step method for preparation of Fe3O4/CS/INH nanoparticles as a targeted drug delivery for tuberculosis. Mater. Sci. Eng. C 2017, 77, 1182–1188. [Google Scholar] [CrossRef]
- Battisti, M.; Vecchione, R.; Casale, C.; Pennacchio, F.A.; Lettera, V.; Jamaledin, R.; Profeta, M.; Di Natale, C.; Imparato, G.; Urciuolo, F. Non-invasive production of multi-compartmental biodegradable polymer microneedles for controlled intradermal drug release of labile molecules. Front. Bioeng. Biotechnol. 2019, 7, 296. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Cheng, Z.; Liu, H.; Shan, W.; Cheng, Z.; Dai, X.; Xue, Y.; Chen, F. Enhancement of Ag85B DNA vaccine immunogenicity against tuberculosis by dissolving microneedles in mice. Vaccine 2018, 36, 4471–4476. [Google Scholar] [CrossRef] [PubMed]
- Di Natale, C.; La Manna, S.; Malfitano, A.M.; Di Somma, S.; Florio, D.; Scognamiglio, P.L.; Novellino, E.; Netti, P.A.; Marasco, D. Structural insights into amyloid structures of the C-terminal region of nucleophosmin 1 in type A mutation of acute myeloid leukemia. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2019, 1867, 637–644. [Google Scholar] [CrossRef] [PubMed]
- La Manna, S.; Scognamiglio, P.L.; Roviello, V.; Borbone, F.; Florio, D.; Di Natale, C.; Bigi, A.; Cecchi, C.; Cascella, R.; Giannini, C. The acute myeloid leukemia-associated Nucleophosmin 1 gene mutations dictate amyloidogenicity of the C-terminal domain. FEBS J. 2019, 286, 2311–2328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Natale, C.; Scognamiglio, P.L.; Cascella, R.; Cecchi, C.; Russo, A.; Leone, M.; Penco, A.; Relini, A.; Federici, L.; Di Matteo, A. Nucleophosmin contains amyloidogenic regions that are able to form toxic aggregates under physiological conditions. FASEB J. 2015, 29, 3689–3701. [Google Scholar] [CrossRef] [Green Version]
- La Manna, S.; Roviello, V.; Scognamiglio, P.L.; Diaferia, C.; Giannini, C.; Sibillano, T.; Morelli, G.; Novellino, E.; Marasco, D. Amyloid fibers deriving from the aromatic core of C-terminal domain of nucleophosmin 1. Int. J. Biol. Macromol. 2019, 122, 517–525. [Google Scholar] [CrossRef]
- Russo, A.; Diaferia, C.; La Manna, S.; Giannini, C.; Sibillano, T.; Accardo, A.; Morelli, G.; Novellino, E.; Marasco, D. Insights into amyloid-like aggregation of H2 region of the C-terminal domain of nucleophosmin. Biochim. Biophys. Acta (BBA)-Proteins Proteom. 2017, 1865, 176–185. [Google Scholar] [CrossRef]
- Scognamiglio, P.L.; Di Natale, C.; Leone, M.; Cascella, R.; Cecchi, C.; Lirussi, L.; Antoniali, G.; Riccardi, D.; Morelli, G.; Tell, G. Destabilisation, aggregation, toxicity and cytosolic mislocalisation of nucleophosmin regions associated with acute myeloid leukemia. Oncotarget 2016, 7, 59129. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhang, X.; Zheng, J.; Liu, D. Self-assembled peptide nanotubes as potential nanocarriers for drug delivery. RSC Adv. 2014, 4, 25461–25469. [Google Scholar] [CrossRef]
- Brandelli, A. Nanostructures as promising tools for delivery of antimicrobial peptides. Mini Rev. Med. Chem. 2012, 12, 731–741. [Google Scholar] [CrossRef]
- Hissam, R.; Farmer, B.L.; Pandey, R.B. Scaffolding of an antimicrobial peptide (KSL) by a scale-down coarse-grained approach. Phys. Chem. Chem. Phys. 2011, 13, 21262–21272. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M.; De Melo Carrasco, L.D. Novel formulations for antimicrobial peptides. Int. J. Mol. Sci. 2014, 15, 18040–18083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selim, A.; Elhaig, M.; Taha, S.; Nasr, E. Antibacterial activity of silver nanoparticles against field and reference strains of Mycobacterium tuberculosis, Mycobacterium bovis and multiple-drugresistant tuberculosis strains. Rev-Off Int. Epizoot. 2018, 37, 823–830. [Google Scholar] [PubMed] [Green Version]
- Harrison, C. Drug researchers pursue new lines of attack against COVID-19. Nat. Biotechnol. 2020, 38, 659–662. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Natural Peptides | ||||
|---|---|---|---|---|
| AMP | Chemical Structure | Source | Activity | Mechanism of Action (MOA) |
| Pantocin wh-1 [22] | Cycle | Pantoea dispersa W18 | Mycobacterium smegmatis mc2 155, H37Ra mice | Unknown |
| Lassomycin [29] | Cycle | Lentzea kentuckyensis | Mtb and Mycobacterium avium subsp. paratuberculosis. | ATP-protease ClpC1P1P2 |
| Bacteriocin AS-48 [30] | Cycle | Enterococcus faecalis | H37Rv, H37Ra, BCG Pasteur 1173, Mt103, CDC1551, GC 1237, H37Rv phoP, SS18b, M. smegmatis mc2 155 | Disruption of cell membranes |
| Micrococcin P1 [35] | Thio-cycle | Staphylococci | H37Rv | Inhibition of protein synthesis |
| Teixobactin [36] | Iso-cycle | Eleftheria terrae (β-proteobacterium) | H37Rv | Inhibition of cell wall synthesis |
| Ecumicin [37] | Cycle | Nonomuraea spp. MJM5123 | H37Rv | Inhibition of ClpC1 |
| Rufomycin I/Ilamycin A [38] | Cycle | Streptomyces sp. (MJM3502), Streptomyces atratus (NRRL B-16927) | Mtb and Mycobacterium abscessus | Inhibition of ClpC1 |
| Synthetic Peptides | ||||
| Several cationic antimicrobial peptides (CAMPs) [23] | Cationic peptide rich in W and R | Peptide libraries | M. smegmatis mc2 6, H37Rv and Mycobacterium tuberculosis lux strain | Pore formation |
| Cinnamic acid (Cin+) CAMP1, 2, 3, 5, 7 [24] | N-terminally modified protein | Five de novo proteins designed from CAMPs [23] | H37Rv, Multi drug resistant Tubercolosis (MDR-Tb) | Pore formation |
| hBD consensus hBD10 [25] | Disulfide bonds | Human β-defensin (hBD) | Multi drug resistant (MDR) strains, H37Rv | Possible activity on Kv channels |
| NZX [26] | Cysteine-rich protein | Plectasin | BCG, H37Rv | Unknown |
| Vaejovis punctatus Antimicrobial peptide (VpAmp1.0, 2.0) [27] | Cycle, disulfide bond | Mexican scorpion Vaejovis punctatus | MDR strains, H37Rv | Cell lysis |
| Cyclohexyl griselimycin (CGM) [28] | Cycle | H37Rv, M. smegmatis mc2 155, | Inhibition of dnaN | |
| Vesicle associated membrane proteins (VAMP) α1, α2, α4 (VapBC30) [39] | α-helix | Fragments of VapBC30 | H37Rv | Inhibition of VapB30/VapC30 |
| d-LAK 120 [40] | D-amino acid derivative, α-helix | MDR strains |
| |
| LL37-analogous peptide(LLAP) [41] | LL-37 | M. smegmatis | Inhibition of ATPase | |
| Synthetic AMPs (SAMPs-Dma) [42] | Dimethylamination and imidazolation | De novo designed | M. smegmatis mc2 155 |
|
| Innate defense regulators ((IDR)-1002, -HH2, IDR-1018) [43,44,45] | Macrophage chemotactic protein-1 (MCP-1) | H37Rv, MDR strains |
| |
| RNAse (RN3) (1–45) (RN6) (1–45) (RN7) (1–45) [46,47] | RNAse (RN)N-terminus | Mycobacterium vaccae; Mycobacterium aurum; M. smegmatis mc2 155; Mycobacterium bovis; bacillus Calmette-Guérin (BCG) |
| |
| Peptide B (Pep-B) [48] | hBD-1 (H β-Defensin-1) | H37Rv |
| |
| Synthetic cyclomarin A [49,50] | Cycle | Cyclomarin A | M. smegmatis, H37Rv | ClpC1 activity inhibition |
| Pandinin-2 (Pin2) based [51] | Short helix | Pandinin-2 | H37Rv | Disruption of cell membranes |
| Cargo System | Antimicrobial Compound | Bacteria |
|---|---|---|
| Liposomes | LL-37 | Escherichia coli [141], Pseudomonas aeruginosa [176] |
| Ni NPs | LL-37 | General Bacteria [142] |
| Au NPs | Surfactin, lactoferrin | Staphylococcus aureus and E. coli [98,144] |
| Ag NPs | Silver | Mycobacterium bovis, Mtb [146,177] |
| Micro and nano-emulsions | Ru complexes | Mtb [147,148,149,150,151] |
| Folate/mannose-conjugated NPs | Ga3* complexes, Rf | Mtb [156] |
| Polymeric NPs | CTZ, Rf | Mtb [159,160] |
| Dissolvable microneedles | Ag85B | Mtb [166] |
| Self-assembly nanoparticles | KSL peptide | Staphylococcus epidermidis, aureus |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Natale, C.; De Benedictis, I.; De Benedictis, A.; Marasco, D. Metal–Peptide Complexes as Promising Antibiotics to Fight Emerging Drug Resistance: New Perspectives in Tuberculosis. Antibiotics 2020, 9, 337. https://doi.org/10.3390/antibiotics9060337
Di Natale C, De Benedictis I, De Benedictis A, Marasco D. Metal–Peptide Complexes as Promising Antibiotics to Fight Emerging Drug Resistance: New Perspectives in Tuberculosis. Antibiotics. 2020; 9(6):337. https://doi.org/10.3390/antibiotics9060337
Chicago/Turabian StyleDi Natale, Concetta, Ilaria De Benedictis, Arianna De Benedictis, and Daniela Marasco. 2020. "Metal–Peptide Complexes as Promising Antibiotics to Fight Emerging Drug Resistance: New Perspectives in Tuberculosis" Antibiotics 9, no. 6: 337. https://doi.org/10.3390/antibiotics9060337
APA StyleDi Natale, C., De Benedictis, I., De Benedictis, A., & Marasco, D. (2020). Metal–Peptide Complexes as Promising Antibiotics to Fight Emerging Drug Resistance: New Perspectives in Tuberculosis. Antibiotics, 9(6), 337. https://doi.org/10.3390/antibiotics9060337