Using GFP-Tagged Escherichia coli to Investigate the Persistence of Fecal Bacteria in Vegetated Wetlands: An Experimental Approach



Abstract
:1. Introduction
2. Results
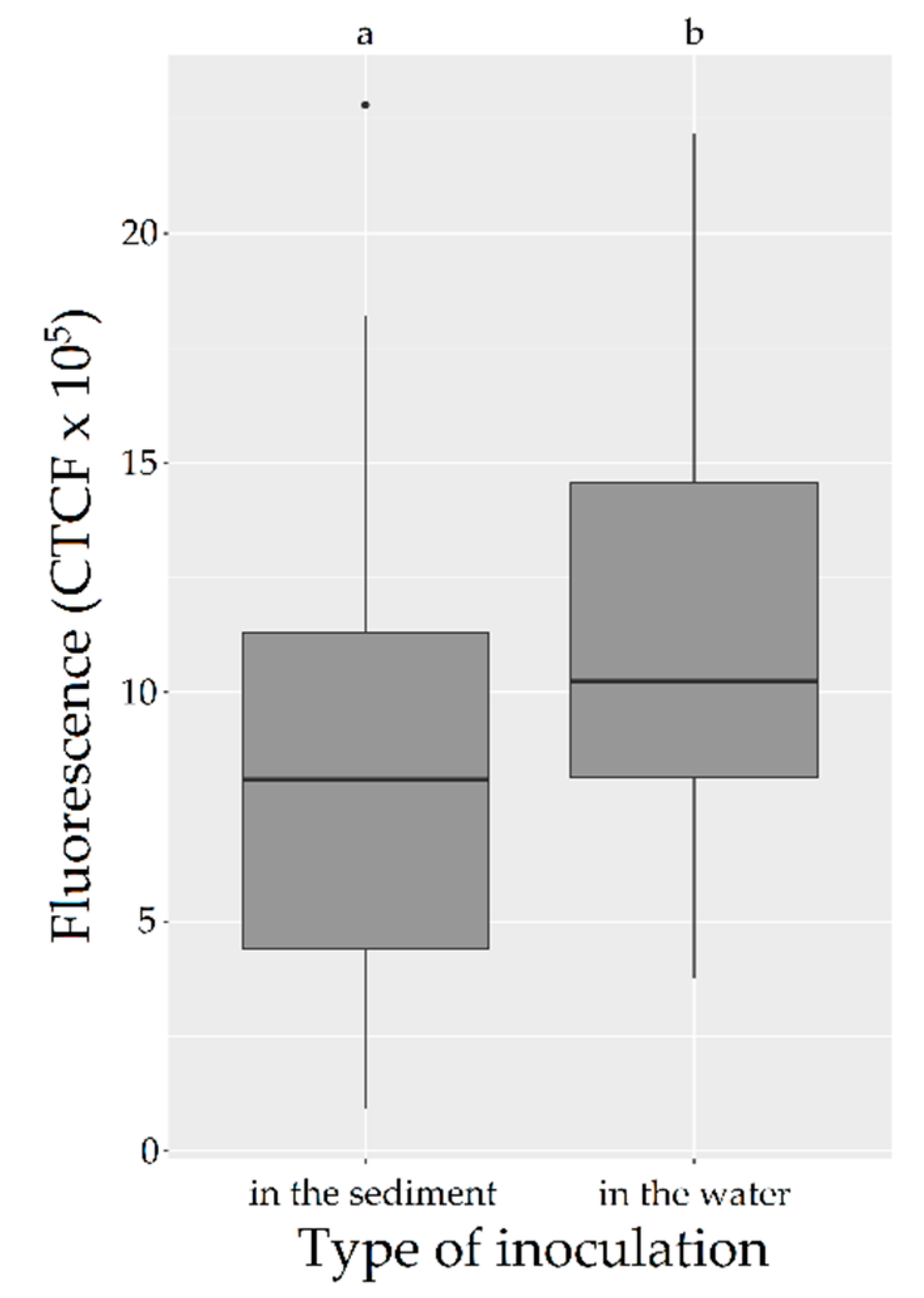
2.1. Influence of the Type of Inoculation in Bacterial Survival
2.2. The Role of Vegetation and Plant Species in Bacterial Survival
3. Discussion
4. Materials and Methods
4.1. Ecosystem and Conditions of Reference
4.2. Experiment Set up
4.2.1. Rhizotrons
4.2.2. Inoculation with GFP-E. coli
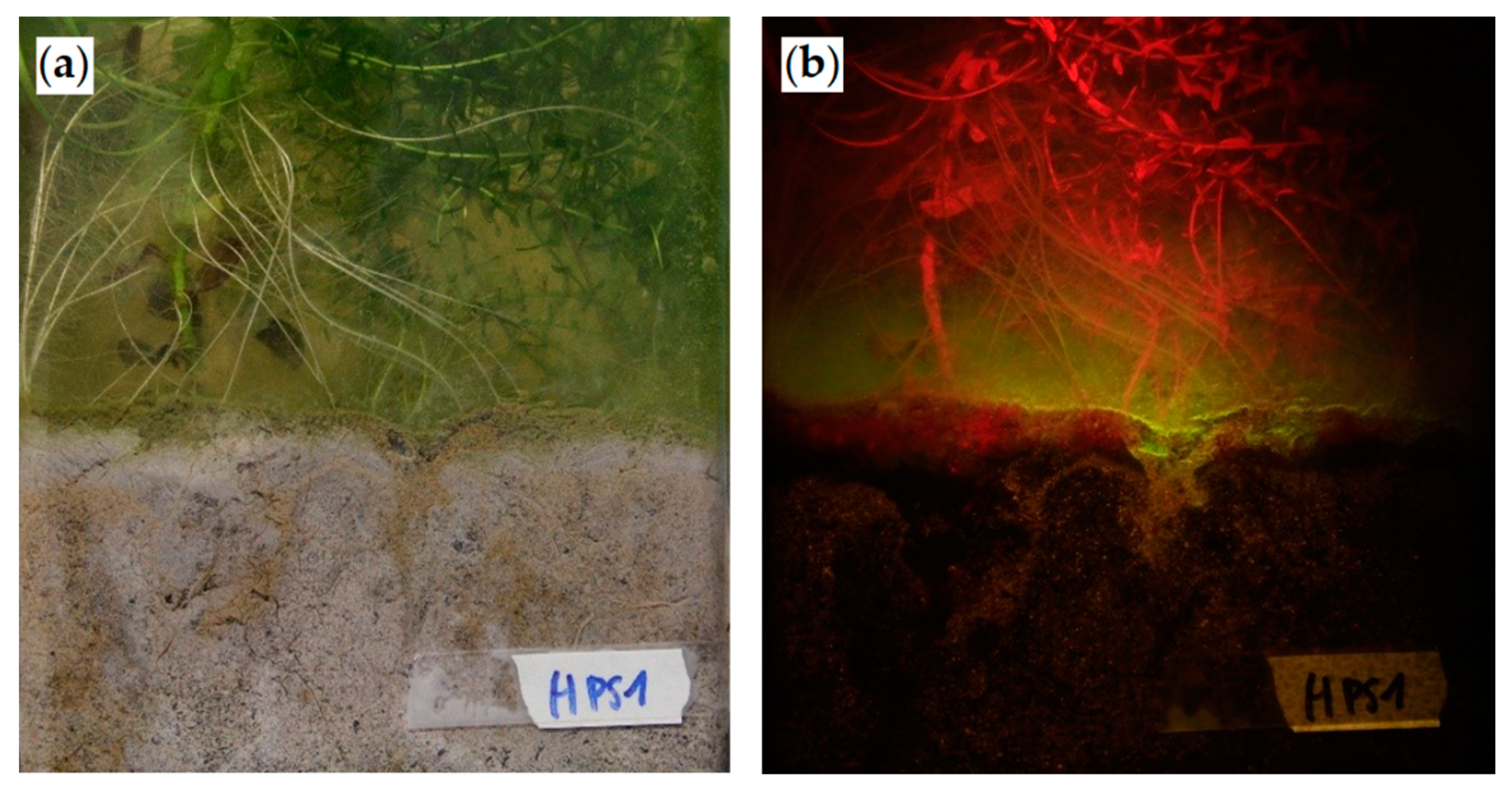
4.3. Image Analysis
4.3.1. Camera and Settings
4.3.2. Image Analysis
4.3.3. Calibration Curve
4.4. Statistics
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Pandey, P.; Kass, P.H.; Soupir, M.L.; Biswas, S.; Singh, V.P. Contamination of water resources by pathogenic bacteria. AMB Express 2014, 4, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Price, R.G.; Wildeboer, D.E. Coli as an indicator of contamination and health risk in environmental waters. In Escherichia Coli—Recent Advances on Physiology, Pathogenesis and Biotechnological Applications; Samie, A., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- World Health Organization. Global Action Plan on Antimicrobial Resistance; WHO Library Cataloguing-in-Publication: Geneva, Switzerland, 2015; ISBN 978 92 4 150976 3. [Google Scholar]
- Rizzo, L.; Manaia, C.M.; Merlin, C.; Schwartz, T.; Dagot, C.; Ploy, M.; Michael, I.; Fatta-Kassinos, D. Urban wastewater treatment plants as hotspots for antibiotic resistant bacteria and genes spread into the environment: A review. Sci. Total Environ. 2013, 447, 345–360. [Google Scholar] [CrossRef]
- Hocquet, D.; Müller, A.; Bertrand, X. What happens in hospitals does not stay in hospitals: Antibiotic-resistant bacteria in hospital wastewater systems. J. Hosp. Infect. 2016, 93, 395–402. [Google Scholar] [CrossRef] [PubMed]
- Manyi-Loh, C.E.; Mamphweli, S.N.; Meyer, E.; Makaka, G.; Simon, M.; Okoh, A.I. An overview of the control of bacterial pathogens in cattle manure. Int. J. Environ. Res. Public Health 2016, 13, 843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.-L.; An, X.; Li, H.; Su, J.; Ma, Y.; Zhu, Y.-G. Long-term field application of sewage sludge increases the abundance of antibiotic resistance genes in soil. Environ. Int. 2016, 92, 1–10. [Google Scholar] [CrossRef] [PubMed]
- McKinney, C.W.; Dungan, R.S.; Moore, A.; Leytem, A.B. Occurrence and abundance of antibiotic resistance genes in agricultural soil receiving dairy manure. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Pachepsky, Y.; Shelton, D.; Jeong, J.; Whelan, G. Survival of manure-borne escherichia coli and fecal coliforms in soil: Temperature dependence as affected by site-specific factors. J. Environ. Qual. 2016, 45, 949–957. [Google Scholar] [CrossRef]
- Heuer, H.; Schmitt, H.; Smalla, K. Antibiotic resistance gene spread due to manure application on agricultural fields. Curr. Opin. Microbiol. 2011, 14, 236–243. [Google Scholar] [CrossRef]
- Ishii, S.; Sadowsky, M.J. Escherichia coli in the environment: Implications for water quality and human health. Microbes Environ. 2008, 23, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Odonkor, S.T.; Ampofo, J.K. Escherichia coli as an indicator of bacteriological quality of water: An overview. Microbiol. Res. 2013, 4, 2. [Google Scholar] [CrossRef]
- Price, J.; Ledford, S.H.; Ryan, M.O.; Toran, L.; Sales, C.M. Wastewater treatment plant effluent introduces recoverable shifts in microbial community composition in receiving streams. Sci. Total Environ. 2018, 613, 1104–1116. [Google Scholar] [CrossRef] [PubMed]
- Henriot, C.P.; Martak, D.; Cuenot, Q.; Loup, C.; Masclaux, H.; Gillet, F.; Bertrand, X.; Hocquet, D.; Bornette, G. Occurrence and ecological determinants of the contamination of floodplain wetlands with Klebsiella pneumoniae and pathogenic or antibiotic-resistant Escherichia coli. FEMS Microbiol. Ecol. 2019, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zedler, J.B. Wetlands at your service: Reducing impacts of agriculture at the watershed scale. Front. Ecol. Environ. 2003, 1, 65–72. [Google Scholar] [CrossRef]
- Vymazal, J. Emergent plants used in free water surface constructed wetlands: A review. Ecol. Eng. 2013, 61, 582–592. [Google Scholar] [CrossRef]
- Rehman, F.; Pervez, A.; Khattak, B.N.; Ahmad, R. Constructed wetlands: Perspectives of the oxygen released in the rhizosphere of macrophytes. CLEAN Soil Air Water 2016, 45. [Google Scholar] [CrossRef]
- Gross, E.M.; Hilt, S.; Lombardo, P.; Mulderij, G. Searching for allelopathic effects of submerged macrophytes on phytoplankton—State of the art and open questions. Hydrobiologia 2007, 584, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, Z.A. Macrophytes-cyanobacteria allelopathic interactions and their implications for water resources management—A review. Limnologica 2017, 63, 122–132. [Google Scholar] [CrossRef]
- Brix, H. Do macrophytes play a role in constructed treatment wetlands? Water Sci. Technol. 1997, 35, 11–17. [Google Scholar] [CrossRef]
- Torrens, A.; Molle, P.; Boutin, C.; Salgot, M. Removal of bacterial and viral indicator in vertical flow constructed wetlands and intermittent sand filters. Desalination 2009, 246, 169–178. [Google Scholar] [CrossRef]
- Vacca, G.; Wand, H.; Nikolausz, M.; Kuschk, P.; Kästner, M. Effect of plants and filter materials on bacteria removal in pilot-scale constructed wetlands. Water Res. 2005, 39, 1361–1373. [Google Scholar] [CrossRef]
- Jousset, A.; Scheu, S.; Bonkowski, M. Secondary metabolite production facilitates establishment of rhizobacteria by reducing both protozoan predation and the competitive effects of indigenous bacteria. Funct. Ecol. 2008, 22, 714–719. [Google Scholar] [CrossRef]
- Sleytr, K.; Tietz, A.; Langergraber, G.; Haberl, R. Investigation of bacterial removal during the filtration process in constructed wetlands. Sci. Total Environ. 2007, 380, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Mburu, N.; Thumbi, G.M.; Mayabi, A.O. Removal of bacterial pathogens from domestic wastewater in a tropical subsurface horizontal flow constructed wetland. In Phytoremediation: Management of Environmental Contaminants; Springer International Publishing: Basel, Switzerland, 2008; Volume 4, pp. 327–346. [Google Scholar]
- Knox, A.K.; Dahlgren, R.A.; Tate, K.W.; Atwill, E.R. Efficacy of natural wetlands to retain nutrient, sediment and microbial pollutants. J. Environ. Qual. 2008, 37, 1837–1846. [Google Scholar] [CrossRef]
- Tailleur, J.; Cates, M.E. Sedimentation, trapping, and rectification of dilute bacteria. Europhys. Lett. Assoc. 2009, 86, 60002. [Google Scholar] [CrossRef] [Green Version]
- Cervantes, A.A. Resuspension of E. coli Under Controlled Flows and Stream Bottom Sediments; Master, Iowa State University: Ames, IA, USA, 2018. [Google Scholar]
- Dunn, A.M.; Silliman, S.E.; Dhamwichukorn, S.; Kulpa, C.F. Demonstration of microbial transport into the capillary fringe via advection from below the water table. J. Hydrol. 2005, 306, 50–58. [Google Scholar] [CrossRef]
- Brennan, F.; Moynihan, E.; Griffiths, B.; Hillier, S.; Owen, J.; Pendlowski, H.; Avery, L.M. Clay mineral type effect on bacterial enteropathogen survival in soil. Sci. Total Environ. 2014, 468–469, 302–305. [Google Scholar] [CrossRef]
- Leff, L.G.; Leff, A.A. Use of green fluorescent protein to monitor survival of genetically engineered bacteria in aquatic environments. Appl. Environ. Microbiol. 1996, 62, 3486–3488. [Google Scholar] [CrossRef] [Green Version]
- Heß, S.; Berendonk, T.U.; Kneis, D. Antibiotic resistant bacteria and resistance genes in the bottom sediment of a small stream and the potential impact of remobilization. FEMS Microbiol. Ecol. 2018, 94. [Google Scholar] [CrossRef] [Green Version]
- Faulwetter, J.L.; Gagnon, V.; Sundberg, C.; Chazarenc, F.; Burr, M.D.; Brisson, J.; Camper, A.K.; Stein, O.R. Microbial processes influencing performance of treatment wetlands: A review. Ecol. Eng. 2009, 35, 987–1004. [Google Scholar] [CrossRef]
- Berg, G.; Eberl, L.; Hartmann, A. The rhizosphere as a reservoir for opportunistic human pathogenic bacteria. Environ. Microbiol. 2005, 7, 1673–1685. [Google Scholar] [CrossRef]
- Bertin, C.; Yang, X.; Weston, L.A. The role of root exudates and allelochemicals in the rhizosphere. Plant Soil 2003, 256, 67–83. [Google Scholar] [CrossRef]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Boil. 2006, 57, 233–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Özbay, H.; Alim, A. Antimicrobial activity of some water plants from the northeastern anatolian region of Turkey. Molecules 2009, 14, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Kozlowski, G.; Jones, R.A.; Nicholls-Vuille, F.-L. Biological flora of central Europe: Baldellia ranunculoides (Alismataceae). Perspect. Plant Ecol. Evol. Syst. 2008, 10, 109–142. [Google Scholar] [CrossRef] [Green Version]
- Harborne, J.B.; Williams, C.A. Occurrence of sulphated flavones and caffeic acid esters in members of the fluviales. Biochem. Syst. Ecol. 1976, 4, 37–41. [Google Scholar] [CrossRef]
- Hilt, S.; Gross, E.M. Can allelopathically active submerged macrophytes stabilise clear-water states in shallow lakes? Basic Appl. Ecol. 2008, 9, 422–432. [Google Scholar] [CrossRef] [Green Version]
- Mues, R. Species specific flavone glucuronides in Elodea species. Biochem. Syst. Ecol. 1983, 11, 261–265. [Google Scholar] [CrossRef]
- Basile, A.; Giordano, S.; López-Sáez, J.A.; Cobianchi, R.C. Antibacterial activity of pure flavonoids isolated from mosses. Phytochemistry 1999, 52, 1479–1482. [Google Scholar] [CrossRef]
- Avelar, F.F.; Matos, M.P.; Matos, M.P.; Borges, A.C. Coliform bacteria removal from sewage in constructed wetlands planted withMentha aquatica. Environ. Technol. 2014, 35, 2095–2103. [Google Scholar] [CrossRef]
- Getahun, Z.; Asres, K.; Mazumder, A.; Bucar, F. Essential oil composition, antibacterial and antioxidant activities of mentha aquatica growing in Ethiopia. Ethiop. Pharm. J. 2008, 26, 9–16. [Google Scholar] [CrossRef]
- Mimica-Dukic, N.; Bozin, B.; Sokovic, M.; Mihajlović, B.; Matavulj, M. Antimicrobial and antioxidant activities of three mentha species essential oils. Planta Med. 2003, 69, 413–419. [Google Scholar] [CrossRef] [PubMed]
- Shahbazi, Y. Chemical composition and in vitro antibacterial activity of mentha spicata essential oil against common food-borne pathogenic bacteria. J. Pathog. 2015, 2015, 1–5. [Google Scholar] [CrossRef]
- Rubinos, M.I.; Pablo, R. Luronium natans, a rare species in the iberian peninsula. Belg. J. Bot. 2004, 137, 85–90. [Google Scholar] [CrossRef]
- Jonsell, B.; Dahlgren, R.M.T.; Clifford, H.T.; Yeo, P.F. The families of the monocotyledons. Structure, evolution and taxonomy. Nord. J. Bot. 1987, 7, 254. [Google Scholar] [CrossRef]
- Brisson, J.; Rodriguez, M.; Martin, C.A.; Proulx, R. Plant diversity effect on water quality in wetlands: A meta-analysis based on experimental systems. Ecol. Appl. 2020, 30. [Google Scholar] [CrossRef]
- Agami, M.; Reddy, K. Competition for space between Eichhornia crassipes (Mart.) Solms and Pistia stratiotes L. cultured in nutrient-enriched water. Aquat. Bot. 1990, 38, 195–208. [Google Scholar] [CrossRef]
- Liang, M.-Q.; Zhang, C.-F.; Peng, C.-L.; Lai, Z.-L.; Chen, D.-F.; Chen, Z.-H. Plant growth, community structure, and nutrient removal in monoculture and mixed constructed wetlands. Ecol. Eng. 2011, 37, 309–316. [Google Scholar] [CrossRef]
- Smith, J.; Edwards, J.; Hilger, H.; Steck, T.R. Sediment can be a reservoir for coliform bacteria released into streams. J. Gen. Appl. Microbiol. 2008, 54, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Garzio-Hadzick, A.; Shelton, D.; Hill, R.; Pachepsky, Y.; Guber, A.K.; Rowland, R. Survival of manure-borne E. coli in streambed sediment: Effects of temperature and sediment properties. Water Res. 2010, 44, 2753–2762. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The rhizosphere microbiome: Significance of plant beneficial, plant pathogenic, and human pathogenic microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Bourceret, A.; Leyval, C.; Thomas, F.; Cébron, A. Rhizosphere effect is stronger than PAH concentration on shaping spatial bacterial assemblages along centimetre-scale depth gradients. Can. J. Microbiol. 2017, 63, 881–893. [Google Scholar] [CrossRef] [PubMed]
- Miranda, G.E.; Prado, F.B.; Delwing, F.; Junior, E.D. Analysis of the fluorescence of body fluids on different surfaces and times. Sci. Justice 2014, 54, 427–431. [Google Scholar] [CrossRef] [PubMed]
- Koenig, M.H.; Yi, E.P.; Sandridge, M.J.; Mathew, A.S.; Demas, J.N. “Open-Box” approach to measuring fluorescence quenching using an iPad screen and digital SLR camera. J. Chem. Educ. 2014, 92, 310–316. [Google Scholar] [CrossRef]
- Cochrane, N.; Walker, G.; Manton, D.J.; Reynolds, E.C. Comparison of quantitative light-induced fluorescence, digital photography and transverse microradiography for quantification of enamel remineralization. Aust. Dent. J. 2012, 57, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Wenzel, M.; Vischer, N.; Strahl, H.; Hamoen, L.W. Assessing membrane fluidity and visualizing fluid membrane domains in bacteria using fluorescent membrane dyes. Bio-Protocol 2018, 8. [Google Scholar] [CrossRef] [Green Version]
- McCloy, R.A.; Rogers, S.; Caldon, C.E.; Lorca, T.; Castro, A.; Burgess, A. Partial inhibition of Cdk1 in G2 phase overrides the SAC and decouples mitotic events. Cell Cycle 2014, 13, 1400–1412. [Google Scholar] [CrossRef] [Green Version]
- Ang, L.Y.E.; Too, H.K.I.; Tan, E.L.; Chow, T.-K.V.; Shek, L.P.-C.; Tham, E.H.; Alonso, S.; Shek, P.-C.L. Antiviral activity of Lactobacillus reuteri Protectis against Coxsackievirus A and Enterovirus 71 infection in human skeletal muscle and colon cell lines. Virol. J. 2016, 13, 111. [Google Scholar] [CrossRef] [Green Version]
- R Development Core Team. R: A Language and Environment for Statistical Computing: Reference Index. Vienna: R Foundation for Statistical Computing 2010. Available online: http://www.polsci.wvu.edu/duval/PS603/Notes/R/fullrefman.pdf (accessed on 22 October 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rhizotron Type | Plant Species | Type of Inoculation with GFP-E. coli | ||
|---|---|---|---|---|
| Water | Sediment | Control 1 | ||
| Rizotrons planted with the four species together (n = 7) | Baldellia ranunculoides, Elodea. canadensis, Mentha aquatica, Sparganiumemersum together | n = 3 | n = 3 | n = 1 |
| Rhizotrons without vegetation (n = 7) | Ø 2 | n = 3 | n = 3 | n = 1 |
| Rhizotrons planted with one single plant species (n = 16) | Baldellia ranunculoides | n = 3 | Ø 3 | n = 1 |
| Elodea canadensis | n = 3 | Ø 3 | n = 1 | |
| Mentha aquatica | n = 3 | Ø 3 | n = 1 | |
| Sparganium emersum | n = 3 | Ø 3 | n = 1 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiapponi, E.; Henriot, C.P.; Bertrand, X.; Hocquet, D.; Bornette, G. Using GFP-Tagged Escherichia coli to Investigate the Persistence of Fecal Bacteria in Vegetated Wetlands: An Experimental Approach. Antibiotics 2020, 9, 335. https://doi.org/10.3390/antibiotics9060335
Chiapponi E, Henriot CP, Bertrand X, Hocquet D, Bornette G. Using GFP-Tagged Escherichia coli to Investigate the Persistence of Fecal Bacteria in Vegetated Wetlands: An Experimental Approach. Antibiotics. 2020; 9(6):335. https://doi.org/10.3390/antibiotics9060335
Chicago/Turabian StyleChiapponi, Emilia, Charles P. Henriot, Xavier Bertrand, Didier Hocquet, and Gudrun Bornette. 2020. "Using GFP-Tagged Escherichia coli to Investigate the Persistence of Fecal Bacteria in Vegetated Wetlands: An Experimental Approach" Antibiotics 9, no. 6: 335. https://doi.org/10.3390/antibiotics9060335
APA StyleChiapponi, E., Henriot, C. P., Bertrand, X., Hocquet, D., & Bornette, G. (2020). Using GFP-Tagged Escherichia coli to Investigate the Persistence of Fecal Bacteria in Vegetated Wetlands: An Experimental Approach. Antibiotics, 9(6), 335. https://doi.org/10.3390/antibiotics9060335