N-Nonyloxypentyl-l-Deoxynojirimycin Inhibits Growth, Biofilm Formation and Virulence Factors Expression of Staphylococcus aureus

 ,
,  ,
,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
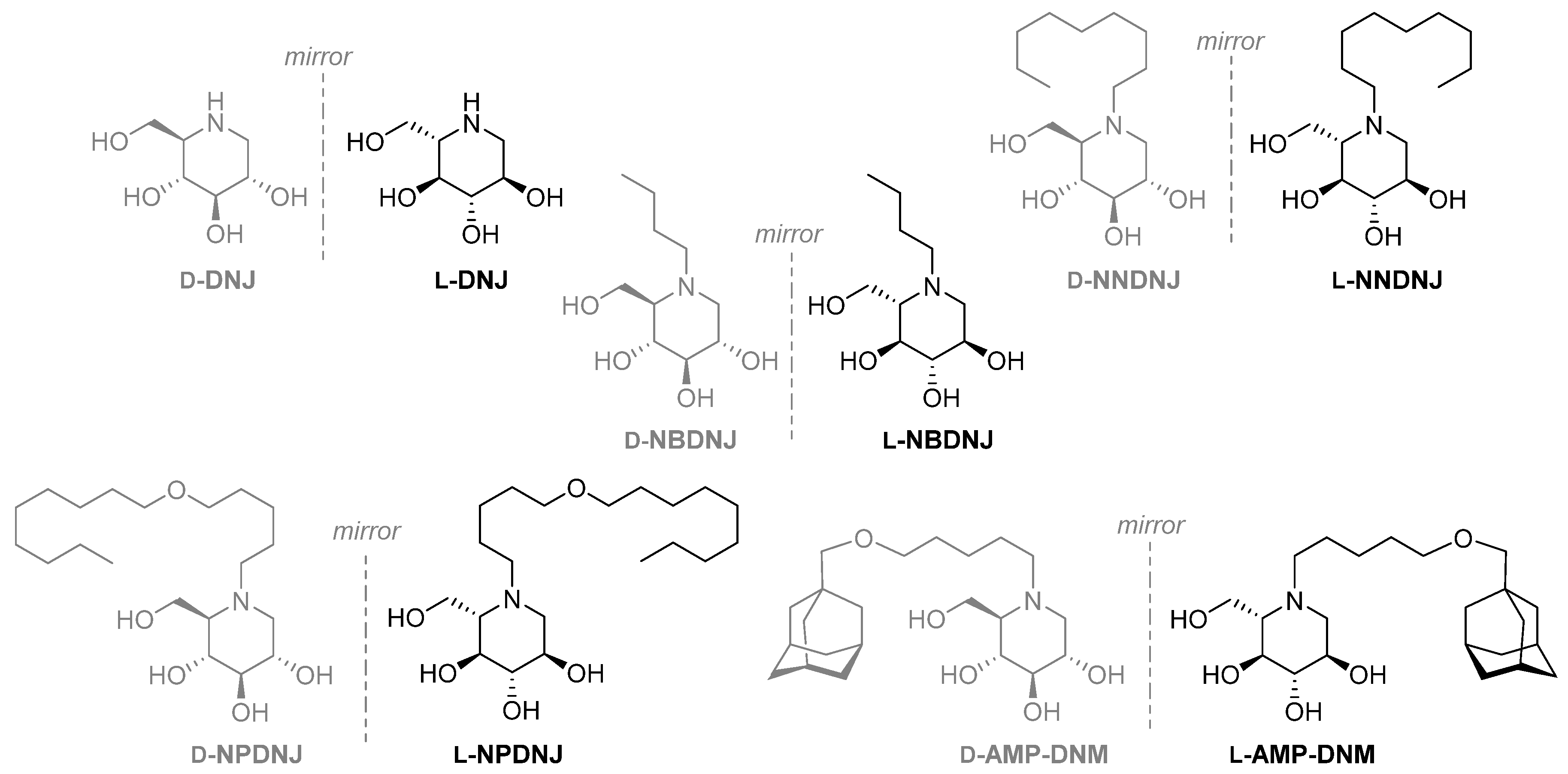
2.1. Antimicrobial Activity of D- and L-DNJ and of A Library of Their N-Alkyl Derivatives
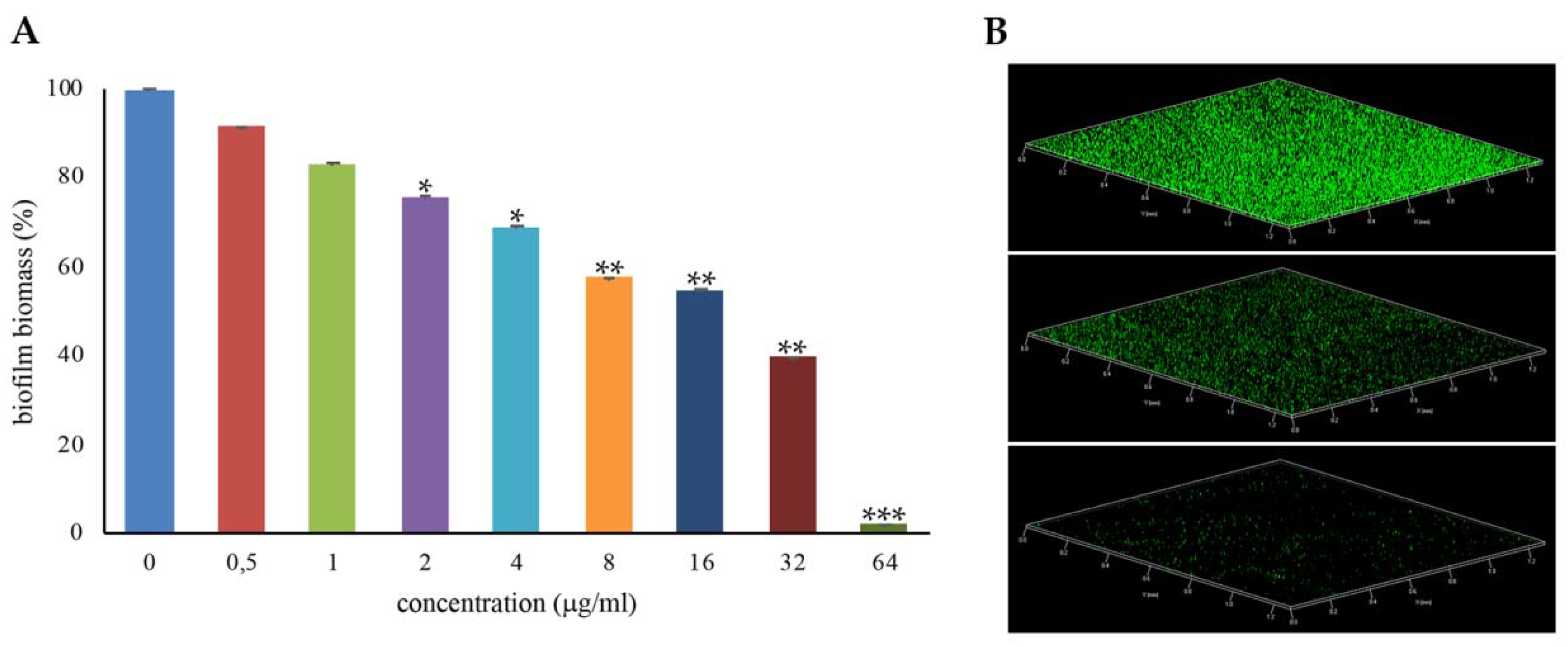
2.2. Effects of l-NPDNJ on Formation of S. Aureus Biofilm
2.3. Antivirulence Activity of l-NPDNJ in S. Aureus ATCC 29213
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Bacterial Strains and Growth Conditions
3.3. Determination of Minimum Inhibitory Concentration (MIC) and Minimum Bactericidal Concentration (MBC)
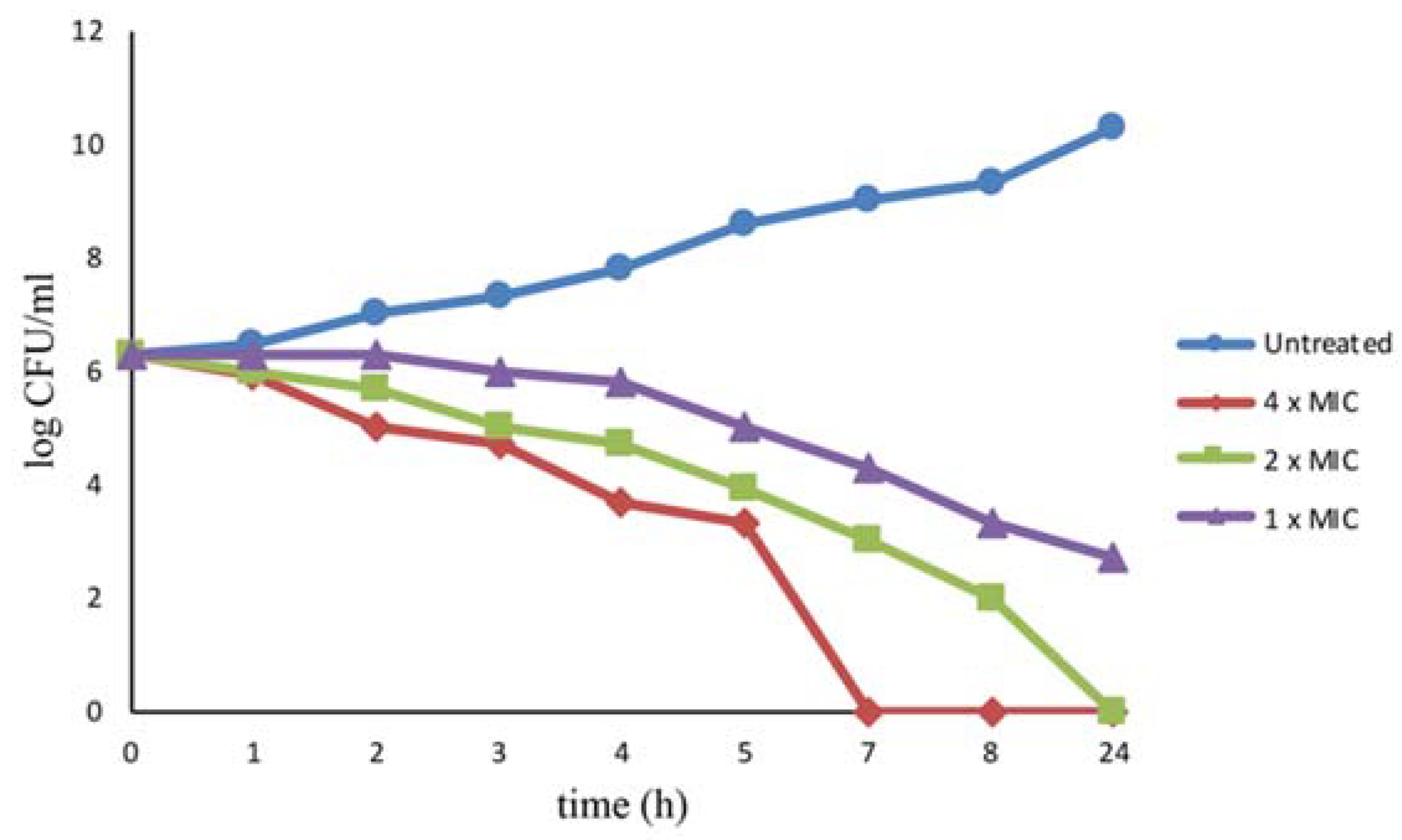
3.4. Time Killing Assay
3.5. Checkerboard Assay
3.6. Biofilm Assay
3.7. RNA Purification and Real-Time RT-PCR
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- David, M.Z.; Daum, R.S. Treatment of Staphylococcus aureus infections. Rev. Microbiol. Immunol. 2017, 409, 325–383. [Google Scholar]
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-resistant Staphylococcus aureus: An overview of basic and clinical research. Nat. Rev. Genet. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, M.; Frees, D.; Ingmer, H. Antibiotic resistance and the MRSA problem. Microbiol. Spectr. 2019, 7, 747–765. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, A.M.S.; Ferreira, F.; Beltrame, C.O.; Côrtes, M.F. The role of biofilms in persistent infections and factors involved in ica-independent biofilm development and gene regulation in Staphylococcus aureus. Crit. Rev. Microbiol. 2017, 2, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Otto, M. Staphylococcal infections: Mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity. Annu. Rev. Med. 2013, 64, 175–188. [Google Scholar] [CrossRef]
- Paharik, A.E.; Horswill, A.R. The staphylococcal biofilm: Adhesins, regulation, and host response. Microbiol. Spectr. 2016, 4, 529–566. [Google Scholar] [CrossRef] [Green Version]
- Jenul, C.; Horswill, A.R. Regulation of Staphylococcus aureus virulence. Microbiol. Spectr. 2018, 6, 669–686. [Google Scholar] [CrossRef]
- Tam, K.; Torres, V.J. Staphylococcus aureus secreted toxins and extracellular enzymes. Microbiol. Spectr. 2019, 7, 640–668. [Google Scholar] [CrossRef]
- Peschel, A.; Otto, M. Phenol-soluble modulins and staphylococcal infection. Nat. Rev. Genet. 2013, 11, 667–673. [Google Scholar] [CrossRef]
- Totsika, M. Disarming pathogens: Benefits and challenges of antimicrobials that target bacterial virulence instead of growth and viability. Futur. Med. Chem. 2017, 9, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Kane, T.L.; Carothers, K.E.; Lee, S.W. Virulence factor targeting of the bacterial pathogen Staphylococcus aureus for vaccine and therapeutics. Curr. Drug Targets 2018, 19, 111–127. [Google Scholar] [CrossRef] [PubMed]
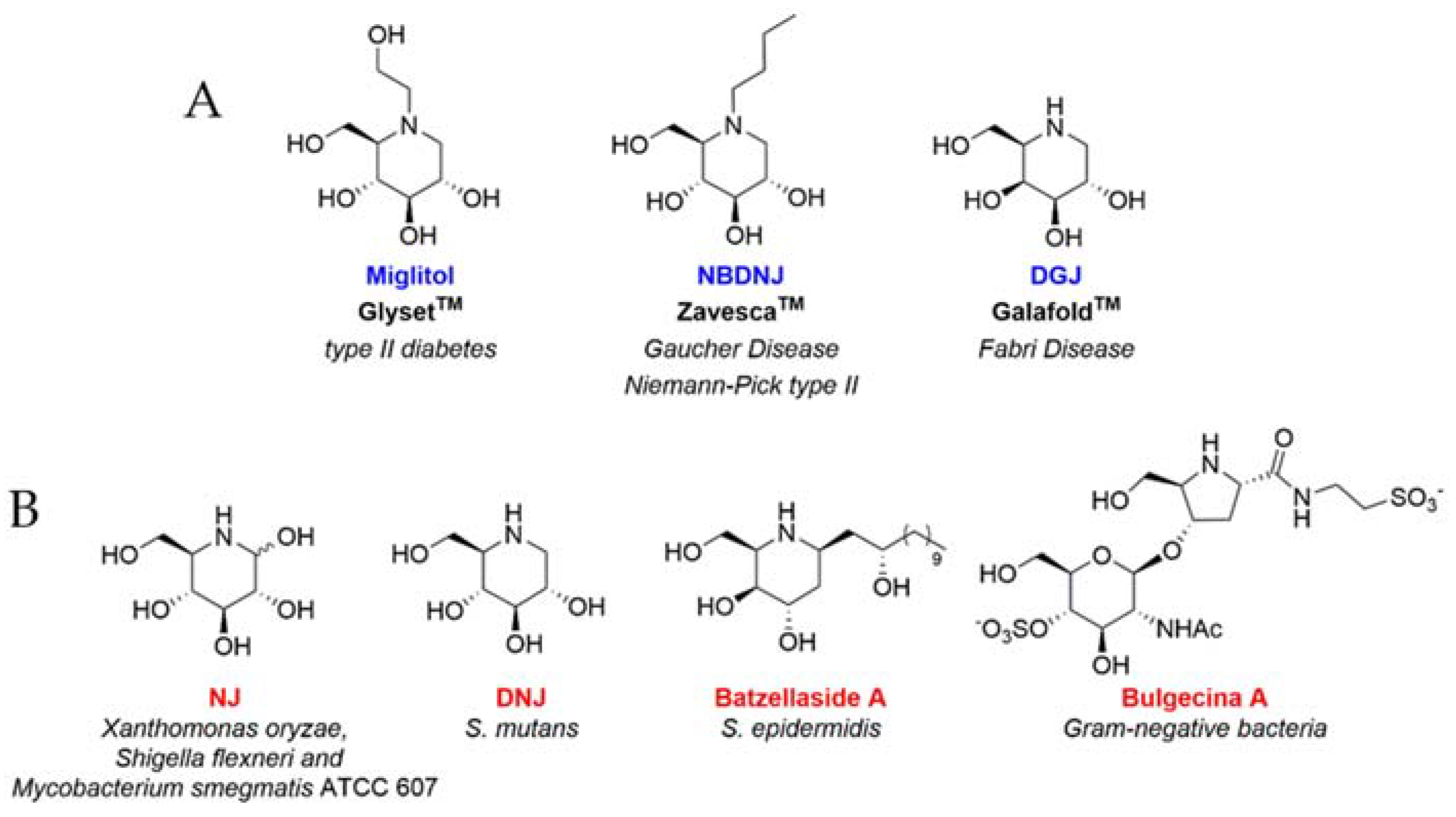
- Compain, P.; Martin, O.R. Iminosugars: From Synthesis to Therapeutic Applications; John Wiley & Sons, Ltd.: Chichester, UK, 2007; ISBN 9780470517437. [Google Scholar]
- Wang, H.; Shen, Y.; Zhao, L.; Ye, Y. 1-Deoxynojirimycin and its derivatives: A mini review of the literature. Curr. Med. Chem. 2020, 27, 1. [Google Scholar] [CrossRef] [PubMed]
- Nash, R.J.; Kato, A.; Yu, C.-Y.; Fleet, G.W. Iminosugars as therapeutic agents: Recent advances and promising trends. Futur. Med. Chem. 2011, 3, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Platt, F.M.; d’Azzo, A.; Davidson, B.L.; Neufeld, E.F.; Tifft, C.J. Lysosomal storage diseases. Nat. Rev. Dis. Prim. 2018, 4, 1–25. [Google Scholar] [CrossRef]
- Fiaux, H.; Popowycz, F.; Favre, S.; Schütz, C.; Vogel, P.; Gerber-Lemaire, S.; Juillerat-Jeanneret, L. Functionalized pyrrolidines inhibit α-mannosidase activity and growth of human glioblastoma and melanoma cells. J. Med. Chem. 2005, 48, 4237–4246. [Google Scholar] [CrossRef] [Green Version]
- Evans, G.B.; Tyler, P.C.; Schramm, V.L. Immucillins in infectious diseases. ACS Infect. Dis. 2017, 4, 107–117. [Google Scholar] [CrossRef]
- Alonzi, D.S.; Scott, K.; Dwek, R.A.; Zitzmann, N. Iminosugar antivirals: The therapeutic sweet spot. Biochem. Soc. Trans. 2017, 45, 571–582. [Google Scholar] [CrossRef] [Green Version]
- Dechecchi, M.C.; Nicolis, E.; Norez, C.; Bezzerri, V.; Borgatti, M.; Mancini, I.; Rizzotti, P.; Ribeiro, C.M.; Gambari, R.; Becq, F.; et al. Anti-inflammatory effect of miglustat in bronchial epithelial cells. J. Cyst. Fibros. 2008, 7, 555–565. [Google Scholar] [CrossRef]
- Best, D.; Jenkinson, S.F.; Saville, A.W.; Alonzi, D.S.; Wormald, M.; Butters, T.; Norez, C.; Becq, F.; Blériot, Y.; Adachi, I.; et al. Cystic fibrosis and diabetes: IsoLAB and isoDAB, enantiomeric carbon-branched pyrrolidine iminosugars. Tetrahedron Lett. 2010, 51, 4170–4174. [Google Scholar] [CrossRef]
- De Fenza, M.; D’Alonzo, D.; Esposito, A.; Munari, S.; Loberto, N.; Santangelo, A.; Lampronti, I.; Tamanini, A.; Rossi, A.; Ranucci, S.; et al. Exploring the effect of chirality on the therapeutic potential of N-alkyl-deoxyiminosugars: Anti-inflammatory response to Pseudomonas aeruginosa infections for application in CF lung disease. Eur. J. Med. Chem. 2019, 175, 63–71. [Google Scholar] [CrossRef]
- Esposito, A.; D’Alonzo, D.; De Fenza, M.; De Gregorio, E.; Tamanini, A.; Lippi, G.; Dechecchi, M.C.; Guaragna, A. Synthesis and therapeutic applications of iminosugars in cystic fibrosis. Int. J. Mol. Sci. 2020, 21, 3353. [Google Scholar] [CrossRef] [PubMed]
- Greimel, P.; Spreitz, J.; Stutz, A.; Wrodnigg, T. Iminosugars and relatives as antiviral and potential anti-infective agents. Curr. Top. Med. Chem. 2003, 3, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Islam, B.; Khan, S.N.; Haque, I.; Alam, M.; Mushfiq, M.; Khan, A.U. Novel anti-adherence activity of mulberry leaves: Inhibition of Streptococcus mutans biofilm by 1-deoxynojirimycin isolated from Morus alba. J. Antimicrob. Chemother. 2008, 62, 751–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hasan, S.; Singh, K.; Danisuddin, M.; Verma, P.K.; Khan, A.U. Inhibition of major virulence pathways of streptococcus mutans by quercitrin and deoxynojirimycin: A synergistic approach of infection control. PLoS ONE 2014, 9, e91736. [Google Scholar] [CrossRef] [Green Version]
- Yoo, Y.; Seo, D.-H.; Lee, H.; Cho, E.-S.; Song, N.-E.; Nam, T.G.; Nam, Y.-D.; Seo, M.-J. Inhibitory effect of Bacillus velezensis on biofilm formation by Streptococcus mutans. J. Biotechnol. 2019, 298, 57–63. [Google Scholar] [CrossRef]
- Strus, M.; Mikołajczyk, D.; Machul, A.; Heczko, P.; Chronowska, A.; Stochel, G.; Gallienne, E.; Nicolas, C.; Martin, O.R.; Kyzioł, A. Effects of the selected iminosugar derivatives on pseudomonas aeruginosa biofilm formation. Microb. Drug Resist. 2016, 22, 638–645. [Google Scholar] [CrossRef]
- Horne, G.; Wilson, F.X.; Tinsley, J.; Williams, D.H.; Storer, R. Iminosugars past, present and future: Medicines for tomorrow. Drug Discov. Today 2011, 16, 107–118. [Google Scholar] [CrossRef]
- D’Alonzo, D.; Guaragna, A.; Palumbo, G. Glycomimetics at the mirror: Medicinal chemistry of L-iminosugars. Curr. Med. Chem. 2009, 16, 473–505. [Google Scholar] [CrossRef]
- Ghisaidoobe, A.; Bikker, P.; De Bruijn, A.C.J.; Godschalk, F.D.; Rogaar, E.; Guijt, M.C.; Hagens, P.; Halma, J.M.; Hart, S.M.V.; Luitjens, S.B.; et al. Identification of potent and selective glucosylceramide synthase inhibitors from a library of N-alkylated iminosugars. ACS Med. Chem. Lett. 2010, 2, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Asano, N. Iminosugars: The potential of carbohydrate analogs. Carbohydr. Chem. State Art Chall. Drug Dev. 2015, 279–301. [Google Scholar] [CrossRef]
- Jenkinson, S.F.; Fleet, G.; Nash, R.J.; Koike, Y.; Adachi, I.; Yoshihara, A.; Morimoto, K.; Izumori, K.; Kato, A. Looking-glass synergistic pharmacological chaperones: DGJ and L-DGJ from the enantiomers of tagatose. Org. Lett. 2011, 13, 4064–4067. [Google Scholar] [CrossRef] [PubMed]
- Rountree, J.S.S.; Butters, T.; Wormald, M.; Boomkamp, S.D.; Dwek, R.A.; Asano, N.; Ikeda, K.; Evinson, E.L.; Nash, R.J.; Fleet, G. Design, synthesis, and biological evaluation of enantiomeric β-N-acetylhexosaminidase inhibitors LABNAc and DABNAc as potential agents against Tay-Sachs and Sandhoff disease. Chem. Med. Chem. 2009, 4, 378–392. [Google Scholar] [CrossRef] [PubMed]
- D’Alonzo, D.; De Fenza, M.; Porto, C.; Iacono, R.; Huebecker, M.; Cobucci-Ponzano, B.; Priestman, D.A.; Platt, F.M.; Parenti, G.; Moracci, M.; et al. N-Butyl-l-deoxynojirimycin (l-NBDNJ): Synthesis of an allosteric enhancer of α-glucosidase activity for the treatment of pompe disease. J. Med. Chem. 2017, 60, 9462–9469. [Google Scholar] [CrossRef] [PubMed]
- Caputo, R.; Ciriello, U.; Festa, P.; Guaragna, A.; Palumbo, G.; Pedatella, S. Stereoselective synthesis of fully protected (S)-1,7-dioxaspiro[5,5]undec-4-ene derivatives of sugars. Eur. J. Org. Chem. 2003, 2617–2621. [Google Scholar] [CrossRef]
- D’Alonzo, D.; Guaragna, A.; Van Aerschot, A.; Herdewijn, P.; Palumbo, G. Toward L-homo-DNA: Stereoselective de novo synthesis of β-L-erythro-hexopyranosyl nucleosides. J. Org. Chem. 2010, 75, 6402–6410. [Google Scholar] [CrossRef]
- Levison, M.E. Pharmacodynamics of antimicrobial drugs. Infect. Dis. Clin. 2004, 18, 451–465. [Google Scholar] [CrossRef]
- Hall, M.; Middleton, R.; Westmacott, D. The fractional inhibitory concentration (FIC) index as a measure of synergy. J. Antimicrob. Chemother. 1983, 11, 427–433. [Google Scholar] [CrossRef]
- Vollaro, A.; Esposito, A.; Antonaki, E.; Iula, V.D.; D’Alonzo, D.; Guaragna, A.; De Gregorio, E. Steroid derivatives as potential antimicrobial agents against Staphylococcus aureus planktonic cells. Microorganisms 2020, 8, 468. [Google Scholar] [CrossRef] [Green Version]
- Bouton, J.; Van Hecke, K.; Rasooly, R.; Van Calenbergh, S. Synthesis of pyrrolidine-based hamamelitannin analogues as quorum sensing inhibitors in Staphylococcus aureus. Beilstein J. Org. Chem. 2018, 14, 2822–2828. [Google Scholar] [CrossRef] [Green Version]
- Von Hoven, G.; Qin, Q.; Neukirch, C.; Husmann, M.; Hellmann, N. Staphylococcus aureus α-toxin: Small pore, large consequences. Biol. Chem. 2019, 400, 1261–1276. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Huang, H.; Rao, X.; Chen, W.; Wang, Z.; Hu, X. Phenol-soluble modulins: Novel virulence-associated peptides of staphylococci. Futur. Microbiol. 2014, 9, 203–216. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J.; Lin, Y.-C.; Gillman, A.; Parks, P.J.; Schlievert, P.M.; Peterson, M.L. Alpha-toxin promotes Staphylococcus aureus mucosal biofilm formation. Front. Microbiol. 2012, 2, 64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huseby, M.J.; Kruse, A.C.; Digre, J.; Kohler, P.L.; Vocke, J.A.; Mann, E.E.; Bayles, K.W.; Bohach, G.A.; Schlievert, P.M.; Ohlendorf, U.H.; et al. Beta toxin catalyzes formation of nucleoprotein matrix in staphylococcal biofilms. Proc. Natl. Acad. Sci. USA 2010, 107, 14407–14412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, C.; Neoh, H.-M.; Nathan, S. Targeting Staphylococcus aureus toxins: A potential form of anti-virulence therapy. Toxins 2016, 8, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung, G.Y.C.; Otto, M. The potential use of toxin antibodies as a strategy for controlling acute Staphylococcus aureus infections. Expert Opin. Ther. Targets 2012, 16, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Xiong, Y.Q.; Willard, J.; Yeaman, M.R.; Cheung, A.L.; Bayer, A.S. Regulation of Staphylococcus aureus α-toxin gene (hla) expression by agr, sarA, and sae in vitro and in experimental infective endocarditis. J. Infect. Dis. 2006, 194, 1267–1275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caiazza, N.C.; O’Toole, G.A. Alpha-toxin is required for biofilm formation by Staphylococcus aureus. J. Bacteriol. 2003, 185, 3214–3217. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.-C.; Liu, F.; Zhu, K.; Shen, J.-Z. Natural products that target virulence factors in antibiotic-resistant Staphylococcus aureus. J. Agric. Food Chem. 2019, 67, 13195–13211. [Google Scholar] [CrossRef]
- Betley, M.; Mekalanos, J. Staphylococcal enterotoxin A is encoded by phage. Science 1985, 229, 185–187. [Google Scholar] [CrossRef]
- Schlievert, P.M.; Bohach, G. Staphylococcal and streptococcal superantigens: An update. In Superantigens: Molecular Basis for the Role in Human Diseases; Fraser, J.D., Kobt, M., Eds.; ASM Press: Washington, DC, USA, 2007; pp. 21–36. [Google Scholar]
- Chen, Y.; Liu, T.; Wang, K.; Hou, C.; Cai, S.; Huang, Y.; Du, Z.; Huang, H.; Kong, J.; Chen, Y. Baicalein inhibits Staphylococcus aureus biofilm formation and the quorum sensing system in vitro. PLoS ONE 2016, 11, e0153468. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Wang, D.; Xiang, H.; Feng, H.; Jiang, Y.; Xia, L.; Dong, J.; Lu, J.; Yu, L.; Deng, X. Subinhibitory concentrations of thymol reduce enterotoxins A and B and α-hemolysin production in Staphylococcus aureus isolates. PLoS ONE 2010, 5, e9736. [Google Scholar] [CrossRef] [Green Version]
- Periasamy, S.; Chatterjee, S.S.; Cheung, G.Y.C.; Otto, M. Phenol-soluble modulins in staphylococci: What are they originally for? Commun. Integr. Biol. 2012, 5, 275–277. [Google Scholar] [CrossRef] [PubMed]
- Schilcher, K.; Andreoni, F.; Haunreiter, V.D.; Seidl, K.; Hasse, B.; Zinkernagel, A.S. Modulation of Staphylococcus aureus biofilm matrix by subinhibitory concentrations of clindamycin. Antimicrob. Agents Chemother. 2016, 60, 5957–5967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Nong, X.-H.; Zhang, X.-Y.; Xu, X.-Y.; Amin, M.; Qi, S.-H. Screening of anti-biofilm compounds from marine-derived fungi and the effects of secalonic acid D on Staphylococcus aureus biofilm. J. Microbiol. Biotechnol. 2017, 27, 1078–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerke, C.; Kraft, A.; Süßmuth, R.; Schweitzer, O.; Götz, F. Characterization of the N-acetylglucosaminyltransferase activity involved in the biosynthesis of the Staphylococcus epidermidis polysaccharide intercellular adhesin. J. Biol. Chem. 1998, 273, 18586–18593. [Google Scholar] [CrossRef] [Green Version]
- Atkins, K.L.; Burman, J.D.; Chamberlain, E.S.; Cooper, J.E.; Poutrel, B.; Bagby, S.; Jenkins, A.T.A.; Feil, E.; Elsen, J.M.V.D.; Jenkins, A.T.A. S. aureus IgG-binding proteins SpA and Sbi: Host specificity and mechanisms of immune complex formation. Mol. Immunol. 2008, 45, 1600–1611. [Google Scholar] [CrossRef]
- Merino, N.; Toledo-Arana, A.; Vergara-Irigaray, M.; Valle, J.; Solano, C.; Calvo, E.; Lopez, J.A.; Foster, T.J.; Penadés, J.R.; Lasa, I. Protein A-mediated multicellular behavior in Staphylococcus aureus. J. Bacteriol. 2008, 191, 832–843. [Google Scholar] [CrossRef] [Green Version]
- Kong, C.; Chee, C.F.; Richter, K.; Thomas, N.; Rahman, N.A.; Nathan, S. Suppression of Staphylococcus aureus biofilm formation and virulence by a benzimidazole derivative, UM-C162. Sci. Rep. 2018, 8, 2758. [Google Scholar] [CrossRef]
- Shrestha, L.; Kayama, S.; Sasaki, M.; Kato, F.; Hisatsune, J.; Tsuruda, K.; Koizumi, K.; Tatsukawa, N.; Yu, L.; Takeda, K.; et al. Inhibitory effects of antibiofilm compound 1 against Staphylococcus aureus biofilms. Microbiol. Immunol. 2016, 60, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Dunman, P.M.; Murphy, E.; Haney, S.; Palacios, D.; Tucker-Kellogg, G.; Wu, S.; Brown, E.L.; Zagursky, R.J.; Shlaes, D.; Projan, S.J. Transcription profiling-based identification ofStaphylococcus aureus genes regulated by the agrand/or sarA Loci. J. Bacteriol. 2001, 183, 7341–7353. [Google Scholar] [CrossRef] [Green Version]
- Goerke, C.; Fluckiger, U.; Steinhuber, A.; Zimmerli, W.; Wolz, C. Impact of the regulatory loci agr, sarA and sae of Staphylococcus aureus on the induction of alpha-toxin during device-related infection resolved by direct quantitative transcript analysis. Mol. Microbiol. 2001, 40, 1439–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coelho, L.; Souza, R.R.; Ferreira, F.; Guimarães, M.A.; Ferreira-Carvalho, B.T.; Figueiredo, A.M.S.; Nozaki, S.; Ogawa, T. Agr RNAIII divergently regulates glucose-induced biofilm formation in clinical isolates of Staphylococcus aureus. Microbiology 2008, 154, 3480–3490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gustafsson, E.; Nilsson, P.; Karlsson, S.; Arvidson, S. Characterizing the dynamics of the quorum-sensing system in Staphylococcus aureus. J. Mol. Microbiol. Biotechnol. 2004, 8, 232–242. [Google Scholar] [CrossRef] [PubMed]
- Queck, S.Y.; Jameson-Lee, M.; Villaruz, A.E.; Bach, T.-H.L.; Khan, B.; Sturdevant, D.E.; Ricklefs, S.M.; Li, M.; Otto, M. RNAIII-independent target gene control by the agr quorum-sensing system: Insight into the evolution of virulence regulation in Staphylococcus aureus. Mol. Cell 2008, 32, 150–158. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Jin, Z.; Sun, B. MgrA negatively regulates biofilm formation and detachment by repressing the expression of psm operons in Staphylococcus aureus. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [Green Version]
- Zapf, R.; Wiemels, R.E.; Keogh, R.; Holzschu, D.L.; Howell, K.M.; Trzeciak, E.; Caillet, A.R.; King, K.A.; Selhorst, S.A.; Naldrett, M.J.; et al. The small RNA Teg41 regulates expression of the alpha phenol-soluble modulins and is required for virulence in Staphylococcus aureus. MBio 2019, 10, e02484-18. [Google Scholar] [CrossRef] [Green Version]
- Geiger, T.; Goerke, C.; Mainiero, M.; Kraus, D.; Wolz, C. The virulence regulator sae of Staphylococcus aureus: Promoter activities and response to phagocytosis-related signals. J. Bacteriol. 2008, 190, 3419–3428. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yeo, W.; Bae, T. The SaeRS two-component system of Staphylococcus aureus. Genes 2016, 7, 81. [Google Scholar] [CrossRef] [Green Version]
- Mainiero, M.; Goerke, C.; Geiger, T.; Gonser, C.; Herbert, S.; Wolz, C. Differential target gene activation by the Staphylococcus aureus two-component system saeRS. J. Bacteriol. 2009, 192, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Voyich, J.M.; Vuong, C.; Dewald, M.; Nygaard, T.K.; Kocianova, S.; Griffith, S.; Jones, J.; Iverson, C.; Sturdevant, D.E.; Braughton, K.R.; et al. The SaeR/S gene regulatory system is essential for innate immune evasion by Staphylococcus aureus. J. Infect. Dis. 2009, 199, 1698–1706. [Google Scholar] [CrossRef] [Green Version]
- Flack, C.E.; Zurek, O.W.; Meishery, D.D.; Pallister, K.B.; Malone, C.L.; Horswill, A.R.; Voyich, J.M. Differential regulation of staphylococcal virulence by the sensor kinase SaeS in response to neutrophil-derived stimuli. Proc. Natl. Acad. Sci. USA 2014, 111, E2037–E2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Gregorio, E.; Roscetto, E.; Iula, V.D.; Martinucci, M.; Zarrilli, R.; Di Nocera, P.P.; Catania, M.R. Development of a real-time PCR assay for the rapid detection of Acinetobacter baumannii from whole blood samples. New Microbiol. 2015, 38, 251–257. [Google Scholar] [PubMed]
- Clinical and Laboratory Standards Institute. CLSI document M07-A9. In Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically: Approved Standard-Ninth Edition; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012. [Google Scholar]
- Esposito, A.; De Gregorio, E.; De Fenza, M.; D’Alonzo, D.; Satawani, A.; Guaragna, A. Expeditious synthesis and preliminary antimicrobial activity of deflazacort and its precursors. RSC Adv. 2019, 9, 21519–21524. [Google Scholar] [CrossRef] [Green Version]
- Esposito, A.; Vollaro, A.; Esposito, E.P.; D’Alonzo, D.; Guaragna, A.; Zarrilli, R.; De Gregorio, E. Antibacterial and antivirulence activity of glucocorticoid PYED-1 against Stenotrophomonas maltophilia. Antibiotics 2020, 9, 105. [Google Scholar] [CrossRef] [Green Version]
- European Committee for Antimicrobial Susceptibility Testing (EUCAST) of the European Society of Clinical Microbiology and Infectious Diseases (ESCMID). Terminology relating to methods for the determination of susceptibility of bacteria to antimicrobial agents. Clin. Microbiol. Infect. 2000, 6, 503–508. [Google Scholar] [CrossRef] [Green Version]
- Vollaro, A.; Esposito, A.; Esposito, E.P.; Zarrilli, R.; Guaragna, A.; De Gregorio, E. PYED-1 inhibits biofilm formation and disrupts the preformed biofilm of Staphylococcus aureus. Antibiotics 2020, 9, 240. [Google Scholar] [CrossRef]
- De Gregorio, E.; Esposito, E.P.; Zarrilli, R.; Di Nocera, P.P. Contact-dependent growth inhibition proteins in Acinetobacter baylyi ADP1. Curr. Microbiol. 2018, 75, 1434–1440. [Google Scholar] [CrossRef] [Green Version]
- Martinucci, M.; Roscetto, E.; Iula, V.D.; Votsi, A.; Catania, M.R.; De Gregorio, E. Accurate identification of members of the Burkholderia cepacia complex in cystic fibrosis sputum. Lett. Appl. Microbiol. 2016, 62, 221–229. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entry | Compound | MIC | MBC |
|---|---|---|---|
| 1 | d-DNJ | >1000 | >1000 |
| 2 | L-DNJ | >1000 | >1000 |
| 3 | d-NBDNJ | >1000 | >1000 |
| 4 | l-NBDNJ | >1000 | >1000 |
| 5 | d-NNDNJ | 1000 | >1000 |
| 6 | l-NNDNJ | 1000 | >1000 |
| 7 | d-NPDNJ | 256 | 1000 |
| 8 | l-NPDNJ | 128 | 256 |
| 9 | d-AMP-DNM | 256 | >1000 |
| 10 | l-AMP-DNM | 512 | >1000 |
| 11 | Gentamicin | 1 | 2 |
| Bacterial Strain | Combination | MICa (μg/mL) | MICc (μg/mL) | FICI |
|---|---|---|---|---|
| S. aureus 00717 | l-NPDNJ/oxacillin | 128/128 | 64/1 | 0.5078 |
| l-NPDNJ/gentamicin | 128/256 | 64/8 | 0.5312 |
| Gene | Forward Primer (5’-3’) | Reverse Primer (5’-3’) | Reference |
|---|---|---|---|
| agrA | TGCGAAGACGATCCAAAAC | TTTAGCTTGCTCAAGCACCTC | [39] |
| arlR | AGTTGCTGGGCTTGATTACG | ATCCTTTTGTGGCTGACGAC | this study |
| capC | CATCCAGAGCGGAATAAAGC | CGGAAATACCCGCTAATGAC | [39] |
| fnbA | AAGCACAAGGACCAATCGAG | ACGCCATAATTACCGTGACC | this study |
| fnbB | GAACATGGTCAAGCACAAGG | ACGCCATAATTACCGTGACC | [39] |
| hla | TCTTGGAACCCGGTATATGG | AGCGAAGTCTGGTGAAAACC | [39] |
| hlb | GTGCCAAAGCCGAATCTAAG | ATCAGCGCGTTTATATTGTCC | [39] |
| hld | AAGGAAGGAGTGATTTCAATGG | TTTGTTCACTGTGTCGATAATCC | [79] |
| icaA | ACGCAGCAGTAGTTCTTGTCG | TGACCATGTTGCGTAACCAC | this study |
| lukD | GTACTTAAGGCAGCCGGAAAC | CGCCCCAATAAAACTGTGAG | [39] |
| psmα | TCAAAAGCTTAATCGAACAATTCAC | AATGGCCCCCTTCAAATAAG | [79] |
| rpoB | ACAACCACTTGGCGGTAAAG | ATGCTTCAAGTGCCCATACC | [39] |
| saeR | CCAAGGGAACTCGTTTTACG | ACGCATAGGGACTTCGTGAC | [39] |
| sarA | TTGCTTTGAGTTGTTATCAATGG | CAATACAGCGAATTCTTCAAAGC | [79] |
| sea | ATTGCCCTAACGTGGACAAC | TGCTCCCTGCAATTCAGAC | [39] |
| sigB | TGATCGCGAACGAGAAATC | ATTGCCGTTCTCTGAAGTCG | [39] |
| spa | AGATGACCCAAGCCAAAGTG | CTTTCGGTGCTTGAGATTCATT | [39] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Gregorio, E.; Esposito, A.; Vollaro, A.; De Fenza, M.; D’Alonzo, D.; Migliaccio, A.; Iula, V.D.; Zarrilli, R.; Guaragna, A. N-Nonyloxypentyl-l-Deoxynojirimycin Inhibits Growth, Biofilm Formation and Virulence Factors Expression of Staphylococcus aureus. Antibiotics 2020, 9, 362. https://doi.org/10.3390/antibiotics9060362
De Gregorio E, Esposito A, Vollaro A, De Fenza M, D’Alonzo D, Migliaccio A, Iula VD, Zarrilli R, Guaragna A. N-Nonyloxypentyl-l-Deoxynojirimycin Inhibits Growth, Biofilm Formation and Virulence Factors Expression of Staphylococcus aureus. Antibiotics. 2020; 9(6):362. https://doi.org/10.3390/antibiotics9060362
Chicago/Turabian StyleDe Gregorio, Eliana, Anna Esposito, Adriana Vollaro, Maria De Fenza, Daniele D’Alonzo, Antonella Migliaccio, Vita Dora Iula, Raffaele Zarrilli, and Annalisa Guaragna. 2020. "N-Nonyloxypentyl-l-Deoxynojirimycin Inhibits Growth, Biofilm Formation and Virulence Factors Expression of Staphylococcus aureus" Antibiotics 9, no. 6: 362. https://doi.org/10.3390/antibiotics9060362
APA StyleDe Gregorio, E., Esposito, A., Vollaro, A., De Fenza, M., D’Alonzo, D., Migliaccio, A., Iula, V. D., Zarrilli, R., & Guaragna, A. (2020). N-Nonyloxypentyl-l-Deoxynojirimycin Inhibits Growth, Biofilm Formation and Virulence Factors Expression of Staphylococcus aureus. Antibiotics, 9(6), 362. https://doi.org/10.3390/antibiotics9060362