Mutated Human P-Selectin Glycoprotein Ligand-1 and Viral Protein-1 of Enterovirus 71 Interactions on Au Nanoplasmonic Substrate for Specific Recognition by Surface-Enhanced Raman Spectroscopy



 and
and 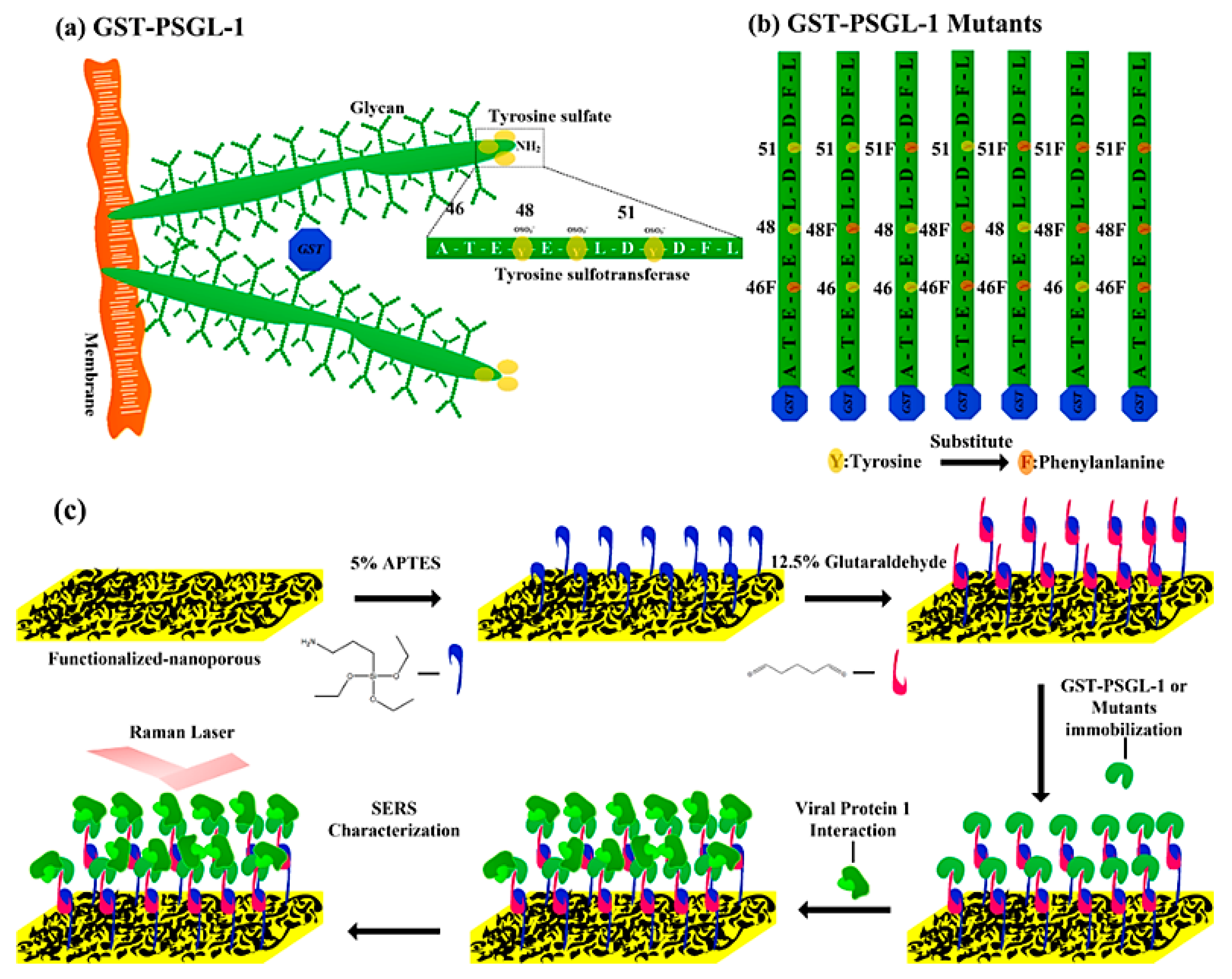{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Experimental Section
2.1. Fabrication of the Au Nanoporous SERS Substrate
2.2. Molecular Probe Detection
2.3. Protein Immobilization on Plasmonic Au Nanoporous Substrate
2.3.1. Protein Preparation
2.3.2. Self-Assembled Monolayers of Proteins on Au Nanoporous Substrate
3. Results and Discussion
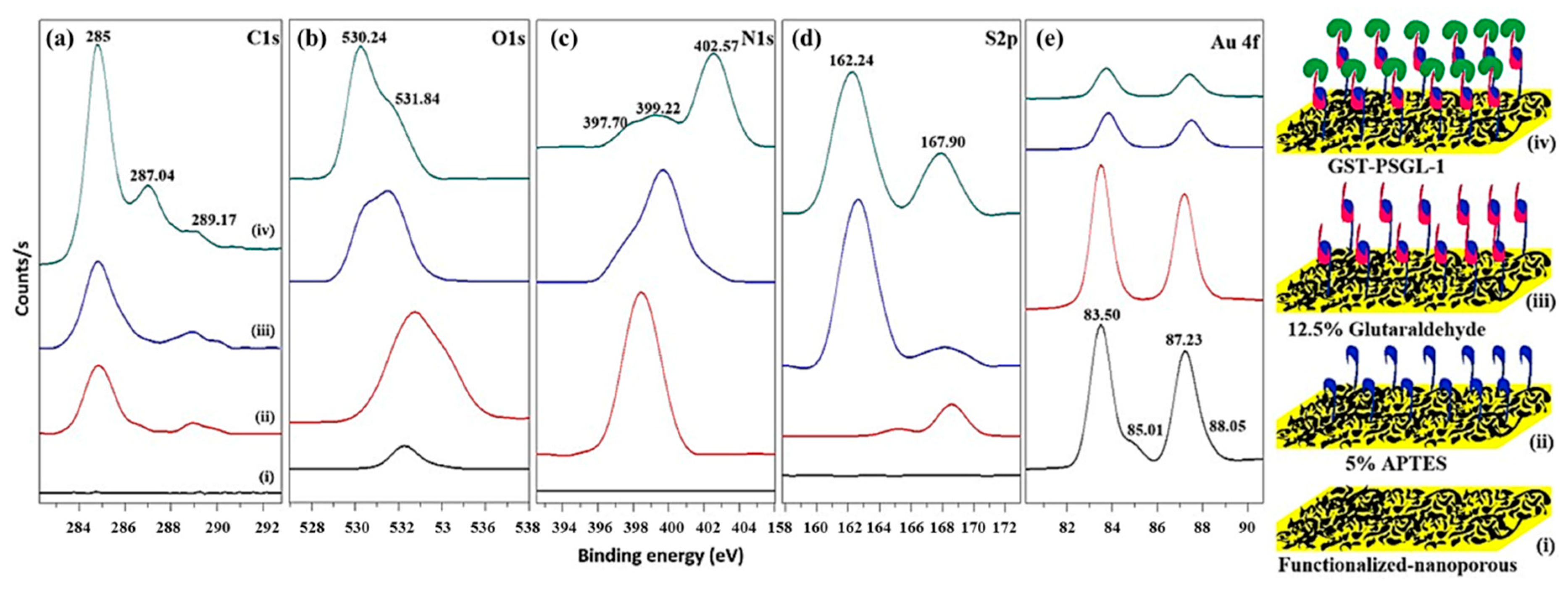
3.1. Characterization of the Plasmonic Au Nanoporous Substrate
3.2. Chemisorption of Protein-Immobilized Plasmonic Au Nanoporous Substrate
3.3. Protein-Protein Interactions on the Plasmonic Au Nanoporous Substrate
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bettelheim, F.R. Tyrosine-O-sulfate in a peptide from fibrinogen. J. Am. Chem. Soc. 1954, 76, 2838–2839. [Google Scholar] [CrossRef]
- Moore, K.L. The biology and enzymology of protein tyrosine O-sulfation. J. Biol. Chem. 2003, 278, 24243–24246. [Google Scholar] [CrossRef] [Green Version]
- Kehoe, J.W.; Bertozzi, C.R. Tyrosine sulfation: A modulator of extracellular protein–protein interactions. Chem. Biol. 2000, 7, R57–R61. [Google Scholar] [CrossRef] [Green Version]
- Huang, B.Y.; Chen, P.C.; Chen, B.H.; Wang, C.C.; Liu, H.F.; Chen, Y.Z.; Chen, C.S.; Yang, Y.S. High-throughput screening of sulfated proteins by using a genome-wide proteome microarray and protein tyrosine sulfation system. Anal. Chem. 2017, 89, 3278–3284. [Google Scholar] [CrossRef]
- Monigatti, F.; Hekking, B.; Steen, H. Protein sulfation analysis-a primer. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2006, 1764, 1904–1913. [Google Scholar] [CrossRef]
- Seibert, C.; Sakmar, T.P. Toward a framework for sulfoproteomics: Synthesis and characterization of sulfotyrosine-containing peptides. Pept. Sci. 2008, 90, 459–477. [Google Scholar] [CrossRef] [Green Version]
- Seibert, C.; Cadene, M.; Sanfiz, A.; Chait, B.T.; Sakmar, T.P. Tyrosine sulfation of CCR5 N-terminal peptide by tyrosylprotein sulfotransferases 1 and 2 follows a discrete pattern and temporal sequence. Proc. Natl. Acad. Sci. USA 2002, 99, 11031–11036. [Google Scholar] [CrossRef] [Green Version]
- Angiari, S.; Constantin, G. Regulation of T cell trafficking by the T cell immunoglobulin and mucin domain 1 glycoprotein. Trends Mol. Med. 2014, 20, 675–684. [Google Scholar] [CrossRef]
- Dong, J.F.; Ye, P.; Schade, A.J.; Gao, S.; Romo, G.M.; Turner, N.T.; López, J.A. Tyrosine sulfation of glycoprotein Ibα role of electrostatic interactions in von willebrand factor binding. J. Biol. Chem. 2011, 276, 16690–16694. [Google Scholar] [CrossRef] [Green Version]
- Farzan, M.; Mirzabekov, T.; Kolchinsky, P.; Wyatt, R.; Cayabyab, M.; Gerard, N.P.; Choe, H. Tyrosine sulfation of the amino terminus of CCR5 facilitates HIV-1 entry. Cell 1999, 96, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Hsu, W.; Rosenquist, G.L.; Ansari, A.A.; Gershwin, M.E. Autoimmunity and tyrosine sulfation. Autoimmun. Rev. 2005, 4, 429–435. [Google Scholar] [CrossRef]
- Braun, P.; Gingras, A.C. History of protein–protein interactions: From egg-white to complex networks. Proteomics 2012, 12, 1478–1498. [Google Scholar] [CrossRef]
- Huang, C.C.; Venturi, M.; Majeed, S.; Moore, M.J.; Phogat, S.; Zhang, M.Y.; Wyatt, R. Structural basis of tyrosine sulfation and VH-gene usage in antibodies that recognize the HIV type 1 coreceptor-binding site on gp120. Proc. Natl. Acad. Sci. USA 2004, 101, 2706–2711. [Google Scholar] [CrossRef] [Green Version]
- Farzan, M.; Babcock, G.J.; Vasilieva, N.; Wright, P.L.; Kiprilov, E.; Mirzabekov, T.; Choe, H. The role of post-translational modifications of the CXCR4 amino terminus in stromal-derived factor 1α association and HIV-1 entry. J. Biol. Chem. 2002, 277, 29484–29489. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, Y.; Shimojima, M.; Tano, Y.; Miyamura, T.; Wakita, T.; Shimizu, H. Human P-selectin glycoprotein ligand-1 is a functional receptor for enterovirus 71. Nat. Med. 2009, 15, 794. [Google Scholar] [CrossRef]
- Nishimura, Y.; Wakita, T.; Shimizu, H. Tyrosine sulfation of the amino terminus of PSGL-1 is critical for enterovirus 71 infection. PLoS Pathog. 2010, 6, 1174. [Google Scholar] [CrossRef] [Green Version]
- Chen, B.H.; Wang, C.C.; Lu, L.Y.; Hung, K.S.; Yang, Y.S. Fluorescence assay for protein post-translational tyrosine sulfation. Anal. Bioanal. Chem. 2013, 405, 1425–1429. [Google Scholar] [CrossRef]
- Goldberg, M.E.; Djavadi-Ohaniance, L. Methods for measurement of antibody/antigen affinity based on ELISA and RIA. Curr. Opin. Immunol. 1993, 5, 278–281. [Google Scholar] [CrossRef]
- Robinson, M.R.; Moore, K.L.; Brodbelt, J.S. Direct identification of tyrosine sulfation by using ultraviolet photodissociation mass spectrometry. J. Am. Soc. Mass Spectrom. 2014, 25, 1461–1471. [Google Scholar] [CrossRef] [Green Version]
- Itkonen, O.; Helin, J.; Saarinen, J.; Kalkkinen, N.; Ivanov, K.I.; Stenman, U.H.; Valmu, L. Mass spectrometric detection of tyrosine sulfation in human pancreatic trypsinogens, but not in tumor-associated trypsinogen. FEBS J. 2008, 275, 289–301. [Google Scholar] [CrossRef]
- Wang, C.C.; Sivashanmugan, K.; Chen, C.K.; Hong, J.R.; Sung, W.I.; Liao, J.D.; Yang, Y.S. Specific unbinding forces between mutated human P-selectin glycoprotein ligand-1 and viral protein-1 measured using force spectroscopy. J. Phys. Chem. Lett. 2017, 8, 5290–5295. [Google Scholar] [CrossRef]
- Pence, I.; Mahadevan-Jansen, A. Clinical instrumentation and applications of Raman spectroscopy. Chem. Soc. Rev. 2016, 45, 1958–1979. [Google Scholar] [CrossRef] [Green Version]
- Sato, H.; Maeda, Y.; Ishigaki, M.; Andriana, B.B. Biomedical Applications of Raman Spectroscopy. In Encyclopedia of Analytical Chemistry: Applications, Theory and Instrumentation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 1–12. [Google Scholar]
- Luo, S.C.; Sivashanmugan, K.; Liao, J.D.; Yao, C.K.; Peng, H.C. Nanofabricated SERS-active substrates for single-molecule to virus detection in vitro: A review. Biosens. Bioelectron. 2014, 61, 232–240. [Google Scholar] [CrossRef]
- Sivashanmugan, K.; Huang, W.L.; Lin, C.H.; Liao, J.D.; Lin, C.C.; Su, W.C.; Wen, T.C. Bimetallic nanoplasmonic gap-mode SERS substrate for lung normal and cancer-derived exosomes detection. J. Taiwan Inst. Chem. Eng. 2017, 80, 149–155. [Google Scholar] [CrossRef]
- Sivashanmugan, K.; Liao, J.D.; You, J.W.; Wu, C.L. Focused-ion-beam-fabricated Au/Ag multilayered nanorod array as SERS-active substrate for virus strain detection. Sensors Actuat. B Chem. 2013, 181, 361–367. [Google Scholar] [CrossRef]
- Sivashanmugan, K.; Liu, P.C.; Tsai, K.W.; Chou, Y.N.; Lin, C.H.; Chang, Y.; Wen, T.C. An anti-fouling nanoplasmonic SERS substrate for trapping and releasing a cationic fluorescent tag from human blood solution. Nanoscale 2017, 9, 2865–2874. [Google Scholar] [CrossRef]
- Pallaoro, A.; Mirsafavi, R.Y.; Culp, W.T.; Braun, G.B.; Meinhart, C.D.; Moskovits, M. Screening for canine transitional cell carcinoma (TCC) by SERS-based quantitative urine cytology. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1279–1287. [Google Scholar] [CrossRef]
- Wang, H.N.; Fales, A.M.; Vo-Dinh, T. Plasmonics-based SERS nanobiosensor for homogeneous nucleic acid detection. Nanomed. Nanotechnol. Biol. Med. 2015, 11, 811–814. [Google Scholar] [CrossRef]
- Kumar, S.; Kumar, A.; Kim, G.H.; Rhim, W.K.; Hartman, K.L.; Nam, J.M. Myoglobin and polydopamine-engineered raman nanoprobes for detecting, imaging, and monitoring reactive oxygen species in biological samples and living cells. Small 2017, 13, 1701584. [Google Scholar] [CrossRef]
- Shin, S.; Lee, J.; Lee, S.; Kim, H.; Seo, J.; Kim, D.; Lee, T. A droplet-based high-throughput SERS platform on a droplet-guiding-track-engraved superhydrophobic substrate. Small 2017, 13, 1602865. [Google Scholar] [CrossRef]
- Xue, L.; Xie, W.; Driessen, L.; Domke, K.F.; Wang, Y.; Schlücker, S.; Steinhart, M. Advanced SERS sensor based on capillarity-assisted preconcentration through gold nanoparticle-decorated porous nanorods. Small 2017, 13, 1603947. [Google Scholar] [CrossRef] [Green Version]
- Arnob, M.M.P.; Zhao, F.; Zeng, J.; Santos, G.M.; Li, M.; Shih, W.C. Laser rapid thermal annealing enables tunable plasmonics in nanoporous gold nanoparticles. Nanoscale 2014, 6, 12470–12475. [Google Scholar] [CrossRef]
- Seker, E.; Reed, M.L.; Begley, M.R. Nanoporous gold: Fabrication, characterization, and applications. Materials 2009, 2, 2188–2215. [Google Scholar] [CrossRef] [Green Version]
- Polat, O.; Seker, E. Effect of surface–molecule interactions on molecular loading capacity of nanoporous gold thin films. J. Phys. Chem. C 2016, 120, 19189–19194. [Google Scholar] [CrossRef] [Green Version]
- Ke, X.; Xu, Y.; Yu, C.; Zhao, J.; Cui, G.; Higgins, D.; Wu, G. Pd-decorated three-dimensional nanoporous Au/Ni foam composite electrodes for H2O2 reduction. J. Mater. Chem. A 2014, 2, 16474–16479. [Google Scholar] [CrossRef]
- Dorofeeva, T.S.; Seker, E. In situ electrical modulation and monitoring of nanoporous gold morphology. Nanoscale 2016, 8, 19551–19556. [Google Scholar] [CrossRef] [Green Version]
- Ishida, S.; Anno, Y.; Takeuchi, M.; Matsuoka, M.; Takei, K.; Arie, T.; Akita, S. Highly photosensitive graphene field-effect transistor with optical memory function. Sci. Rep. 2015, 5, 15491. [Google Scholar] [CrossRef] [Green Version]
- Kim, W.K.; Lee, J.L. Effect of oxygen plasma treatment on reduction of contact resistivity at pentacene/Au interface. Appl. Phys. Lett. 2006, 88, 262102. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Sun, D.; Fu, J.; Yuan, R.; Li, Z. Assembly of evenly distributed Au nanoparticles on thiolated reduced graphene oxide as an active and robust catalyst for hydrogenation of 4-nitroarenes. RSC Adv. 2014, 4, 11003–11011. [Google Scholar] [CrossRef]
- Pasternack, R.M.; Rivillon, A.S.; Chabal, Y.J. Attachment of 3-(aminopropyl) triethoxysilane on silicon oxide surfaces: Dependence on solution temperature. Langmuir 2008, 24, 12963–12971. [Google Scholar] [CrossRef]
- Jastrzebska, M.; Wrzalik, R.; Kocot, A.; Zalewska-Rejdak, J.; Cwalina, B. Raman spectroscopic study of glutaraldehyde-stabilized collagen and pericardium tissue. J. Biomater. Sci. Polym. Ed. 2003, 14, 185–197. [Google Scholar] [CrossRef]
- Satheeshkumar, E.; Karuppaiya, P.; Sivashanmugan, K.; Chao, W.T.; Tsay, H.S.; Yoshimura, M. Biocompatible 3D SERS substrate for trace detection of amino acids and melamine. Pectroc. Acta Pt. A-Molec. Biomolec. Spectr. 2017, 181, 91–97. [Google Scholar] [CrossRef]
- Madzharova, F.; Heiner, Z.; Kneipp, J. Surface enhanced hyper-Raman scattering of the amino acids tryptophan, Histidine, Phenylalanine, and Tyrosine. J. Phys. Chem. C 2017, 121, 1235–1242. [Google Scholar] [CrossRef]
- Hernández, B.; Coïc, Y.M.; Pflüger, F.; Kruglik, S.G.; Ghomi, M. All characteristic Raman markers of tyrosine and tyrosinate originate from phenol ring fundamental vibrations. J. Raman Spectrosc. 2016, 47, 210–220. [Google Scholar] [CrossRef]
- Fischer, W.B.; Eysel, H.H. Polarized Raman spectra and intensities of aromatic amino acids phenylalanine, tyrosine and tryptophan. Spectroc. Acta Pt. A-Molec. Biomolec. Spectr. 1992, 48, 725–732. [Google Scholar] [CrossRef]
- Bugaev, K.O.; Zelenina, A.A.; Volodin, V.A. Vibrational spectroscopy of chemical species in silicon and silicon-rich nitride thin films. J. Spectrosc. 2012, 2012, 281851. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.T.; Li, D.W.; Cao, Y.; Long, Y.T. Label-free in-situ monitoring of protein tyrosine nitration in blood by surface-enhanced Raman spectroscopy. Biosens. Bioelectron. 2015, 69, 1–7. [Google Scholar] [CrossRef]
- Brewster, V.L.; Ashton, L.; Goodacre, R. Monitoring the glycosylation status of proteins using Raman spectroscopy. Anal. Chem. 2011, 83, 6074–6081. [Google Scholar] [CrossRef]
- Davies, H.S.; Singh, P.; Deckert-Gaudig, T.; Deckert, V.; Rousseau, K.; Ridley, C.E.; Blanch, E.W. Secondary structure and glycosylation of mucus glycoproteins by Raman spectroscopies. Anal. Chem. 2016, 88, 11609–11615. [Google Scholar] [CrossRef] [Green Version]
- Ravikumar, B.; Rajaram, R.K.; Ramakrishnan, V. Raman and IR spectral studies of L-phenylalanine L-phenylalaninium dihydrogenphosphate and DL-phenylalaninium dihydrogenphosphate. J. Raman Spectrosc. 2006, 37, 597–605. [Google Scholar] [CrossRef]
- Arp, Z.; Autrey, D.; Laane, J.; Overman, S.A.; Thomas, G.J. Tyrosine Raman signatures of the filamentous virus Ff are diagnostic of non-hydrogen-bonded phenoxyls: Demonstration by Raman and infrared spectroscopy of p-cresol vapor. Biochemistry 2001, 40, 2522–2529. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Martin, S.J.; Chinoy, Z.S.; Liu, L.; Rittgers, B.; Dluhy, R.A.; Boons, G.J. Label-free detection of glycan–protein interactions for array development by surface-enhanced Raman spectroscopy (SERS). Chem.-Eur. J. 2016, 22, 11180–11185. [Google Scholar] [CrossRef] [PubMed] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sivashanmugan, K.; Lee, H.; Liao, J.-D.; Wang, C.-C.; Lin, C.-H.; Yang, Y.-S.; Sitjar, J. Mutated Human P-Selectin Glycoprotein Ligand-1 and Viral Protein-1 of Enterovirus 71 Interactions on Au Nanoplasmonic Substrate for Specific Recognition by Surface-Enhanced Raman Spectroscopy. Coatings 2020, 10, 403. https://doi.org/10.3390/coatings10040403
Sivashanmugan K, Lee H, Liao J-D, Wang C-C, Lin C-H, Yang Y-S, Sitjar J. Mutated Human P-Selectin Glycoprotein Ligand-1 and Viral Protein-1 of Enterovirus 71 Interactions on Au Nanoplasmonic Substrate for Specific Recognition by Surface-Enhanced Raman Spectroscopy. Coatings. 2020; 10(4):403. https://doi.org/10.3390/coatings10040403
Chicago/Turabian StyleSivashanmugan, Kundan, Han Lee, Jiunn-Der Liao, Chen-Chu Wang, Chen-Hsueh Lin, Yuh-Shyong Yang, and Jaya Sitjar. 2020. "Mutated Human P-Selectin Glycoprotein Ligand-1 and Viral Protein-1 of Enterovirus 71 Interactions on Au Nanoplasmonic Substrate for Specific Recognition by Surface-Enhanced Raman Spectroscopy" Coatings 10, no. 4: 403. https://doi.org/10.3390/coatings10040403
APA StyleSivashanmugan, K., Lee, H., Liao, J. -D., Wang, C. -C., Lin, C. -H., Yang, Y. -S., & Sitjar, J. (2020). Mutated Human P-Selectin Glycoprotein Ligand-1 and Viral Protein-1 of Enterovirus 71 Interactions on Au Nanoplasmonic Substrate for Specific Recognition by Surface-Enhanced Raman Spectroscopy. Coatings, 10(4), 403. https://doi.org/10.3390/coatings10040403