A Systematic Survey and Characterization of Enhancers that Regulate Sox3 in Neuro-Sensory Development in Comparison with Sox2 Enhancers

Abstract
:1. Introduction
2. Results
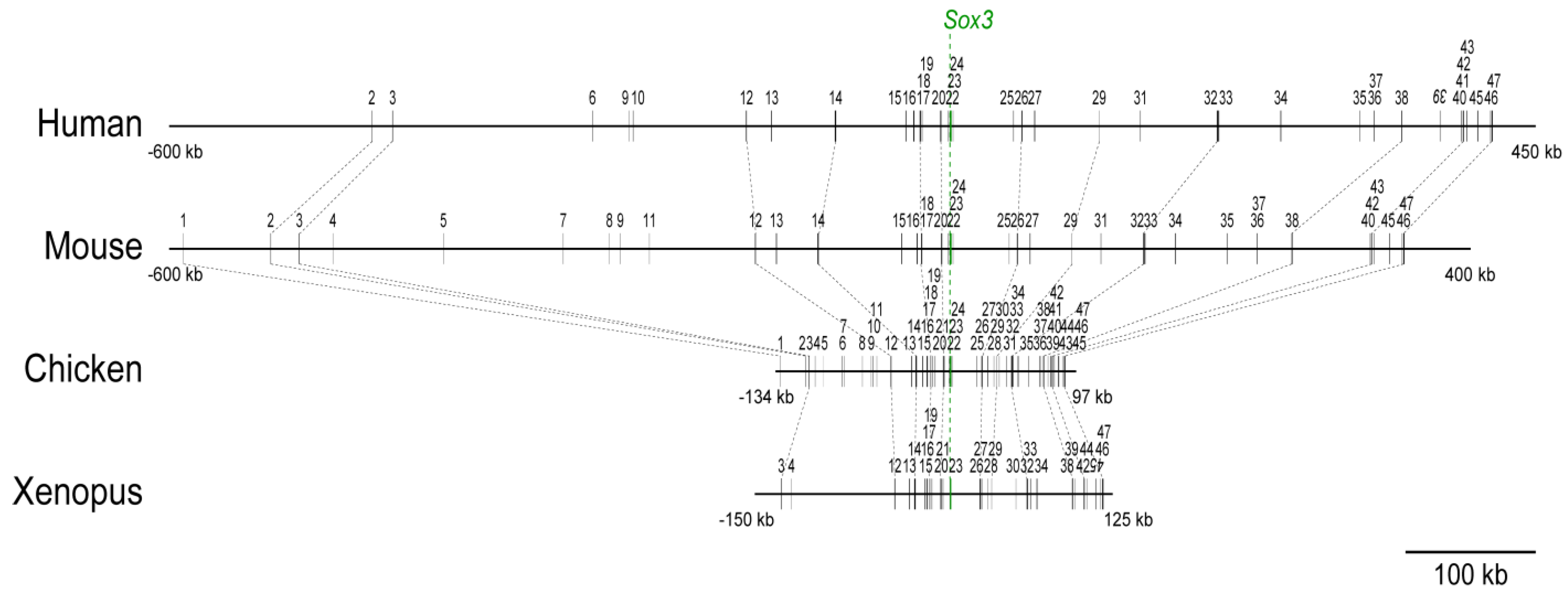
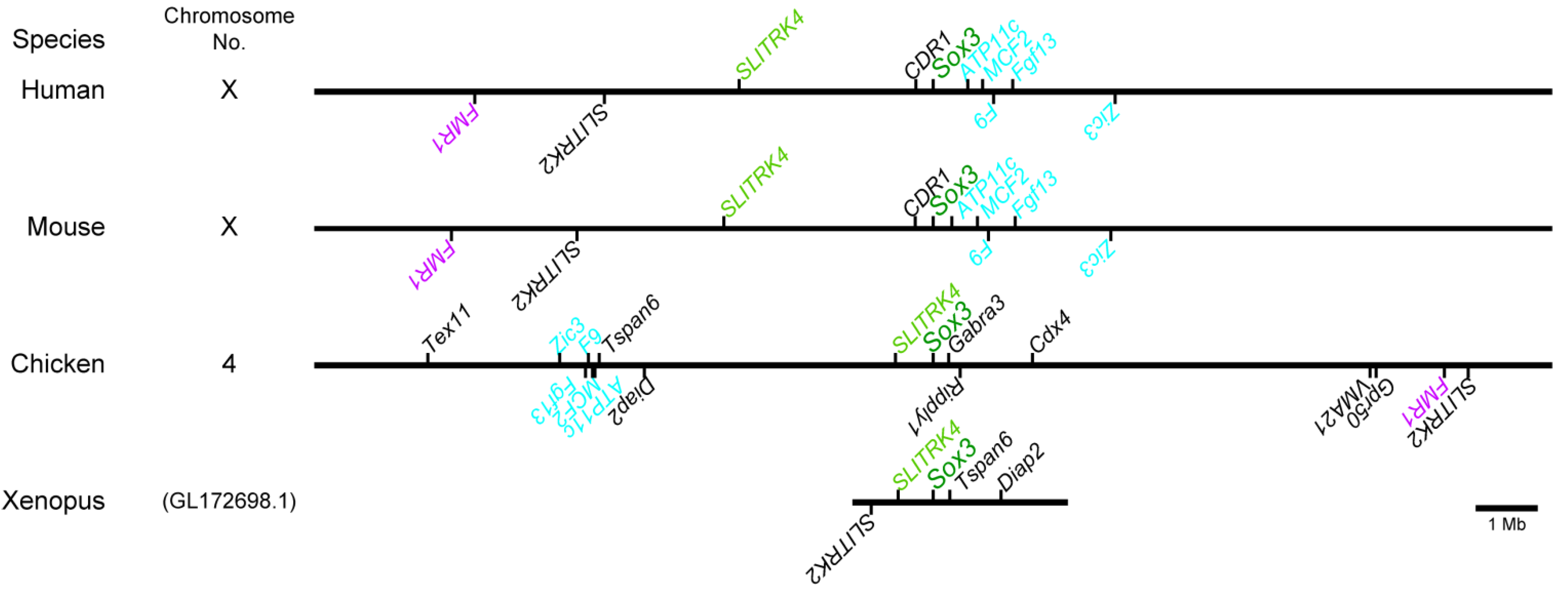
2.1. Distribution of the Conserved Sequence Blocks (CSBs) in the Region Surrounding the Sox3 Gene Locus in Various Vertebrate Species

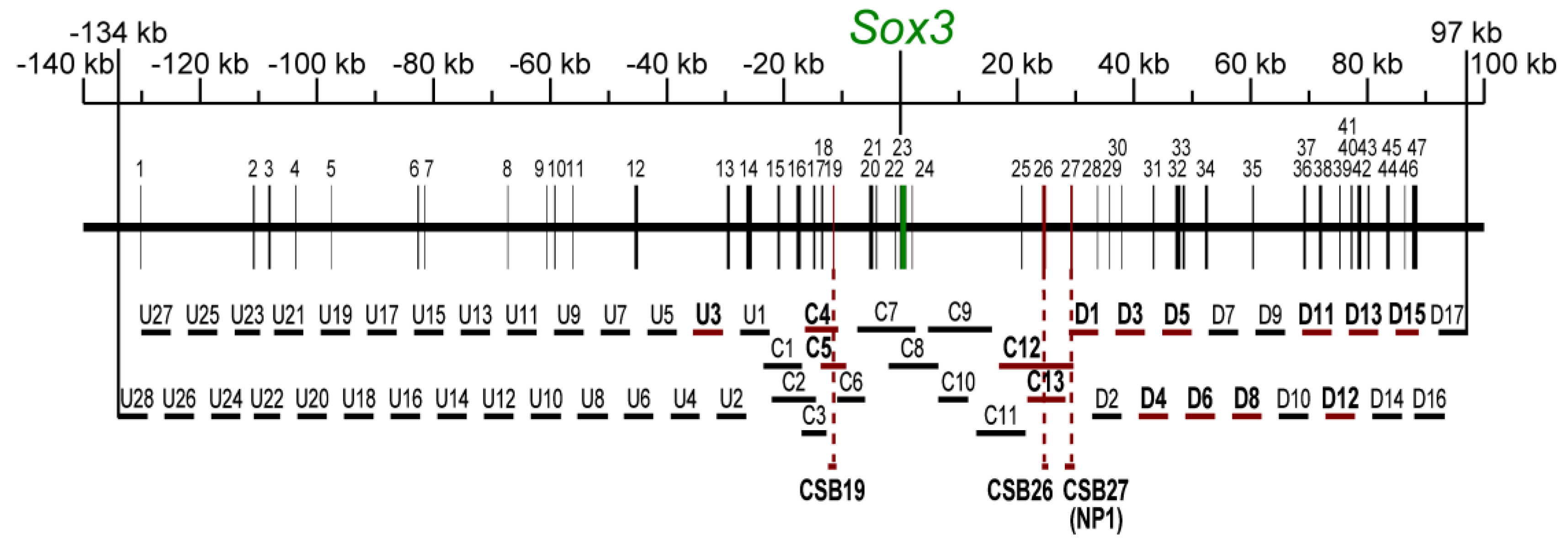
2.2. Screening for Enhancer Sequences and Their Characterization



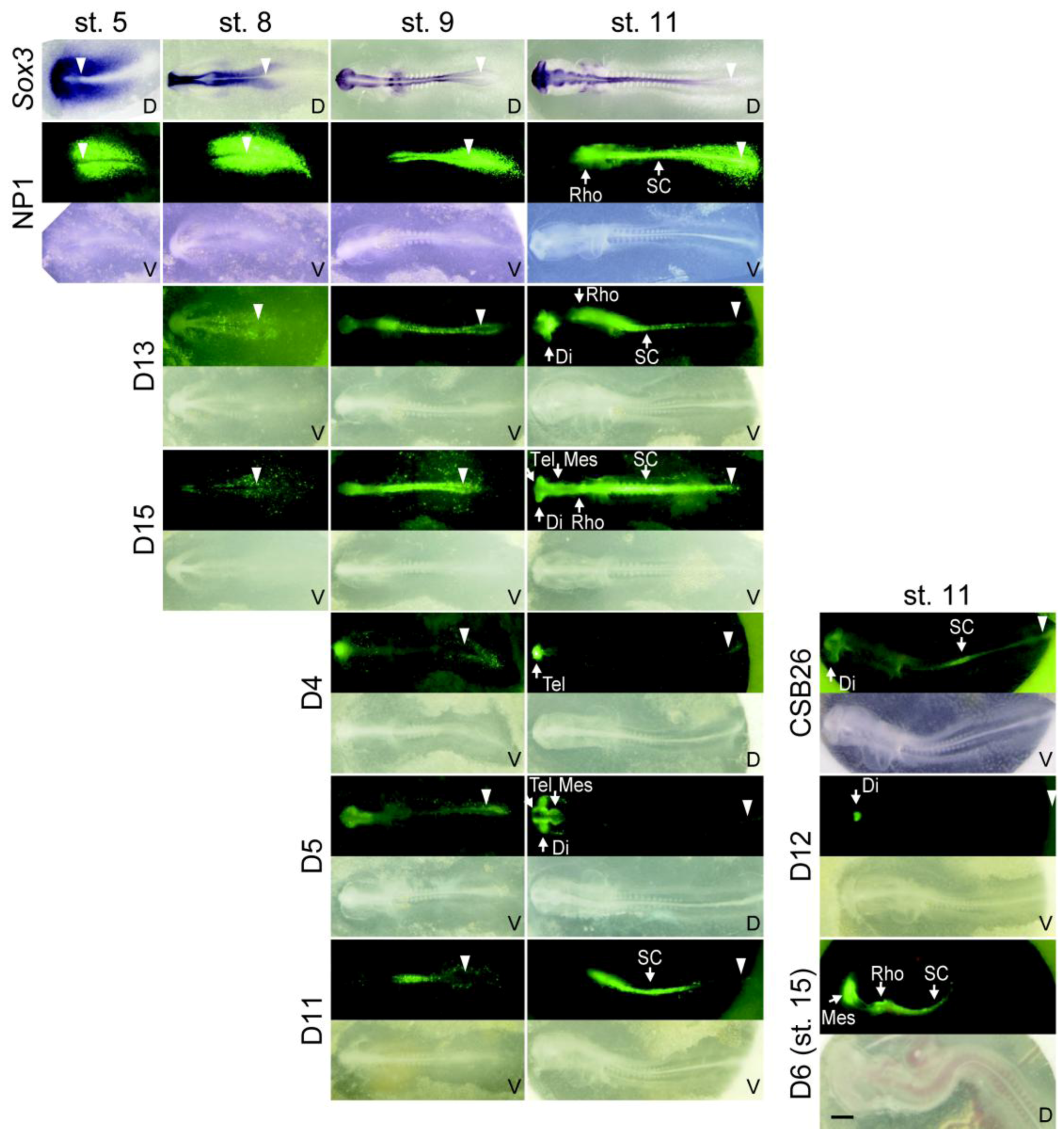
2.3. The Time Course of the Activation of Neural Enhancers
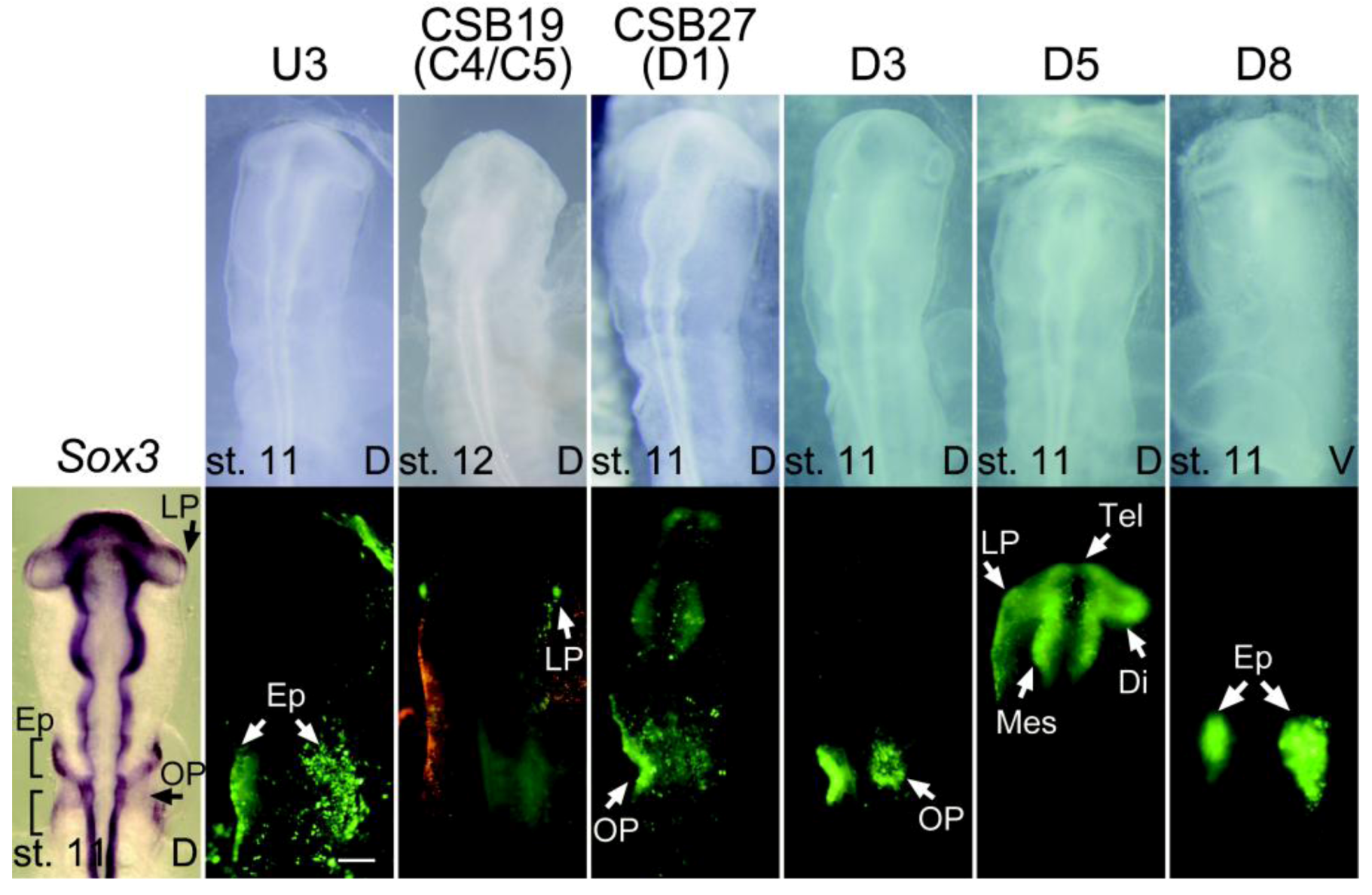
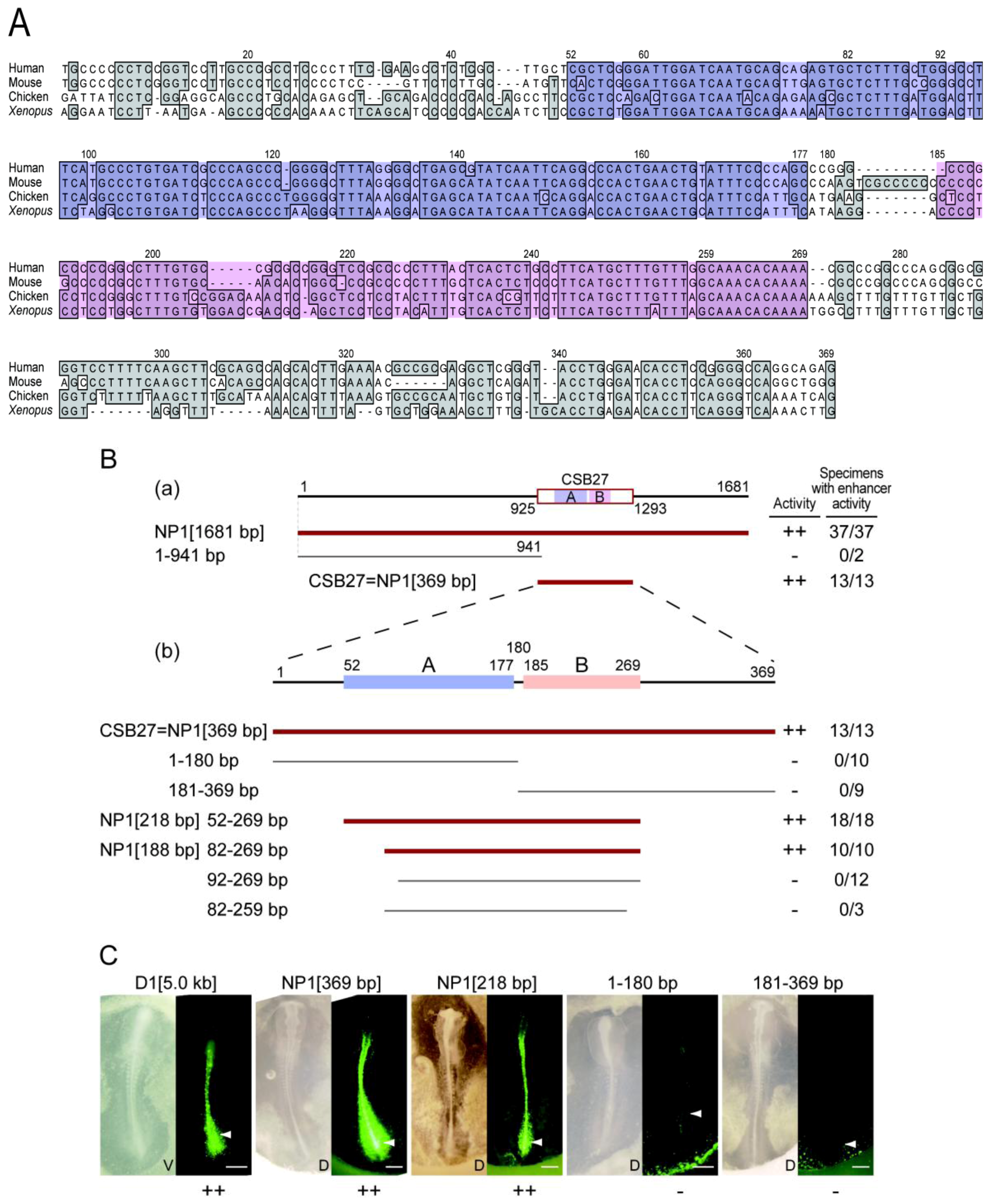
2.4. Functional Dissection of the NP1 Enhancer Region

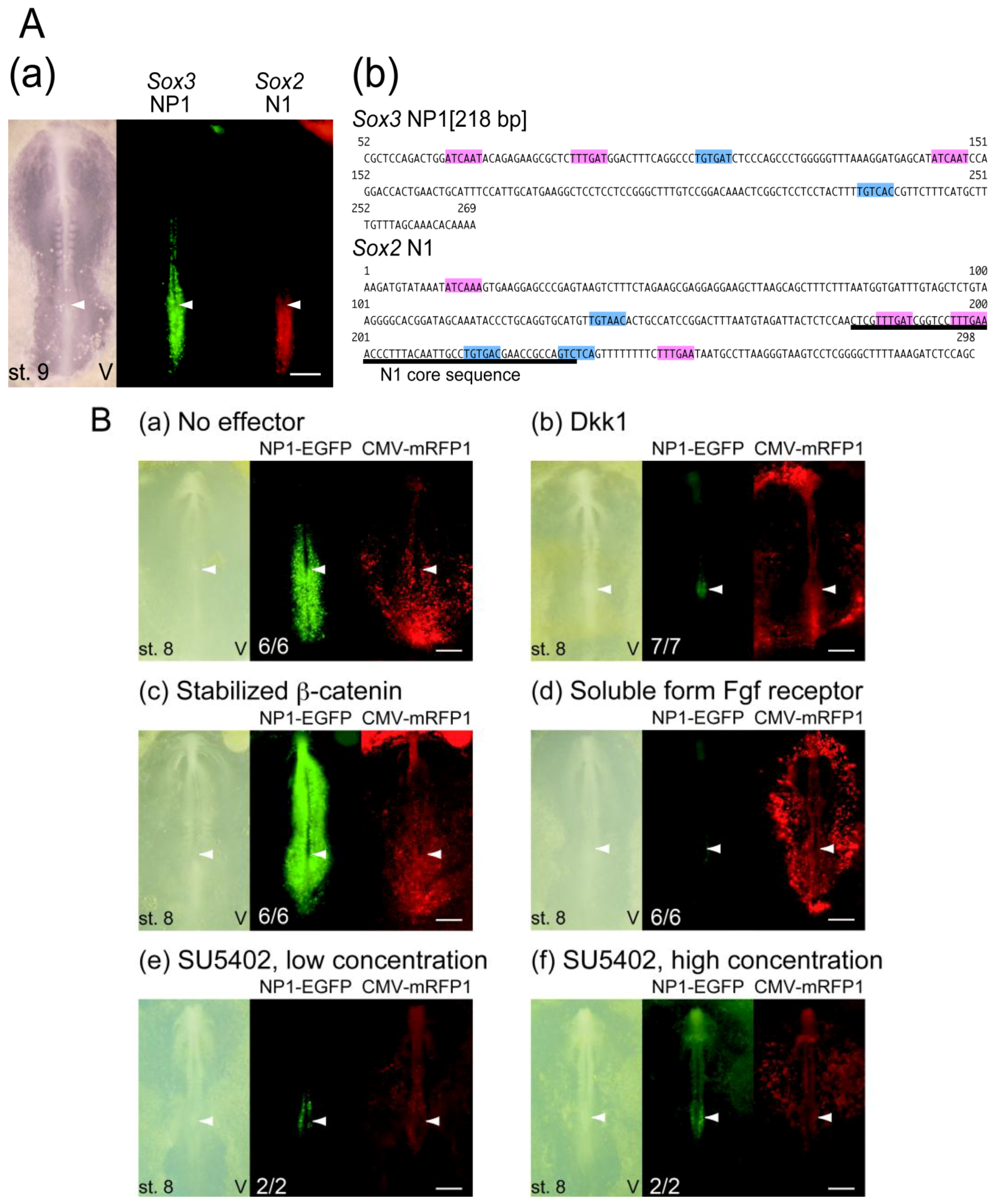
2.5. Regulation of the NP1 Enhancer by Wnt and Fgf Signals

3. Discussion
3.1. Regulation of the Sox3 Gene
3.2. Comparison of the Regulation of Sox3 and Sox2
3.3. Phylogenetic Conservation of Sox3 Regulation
4. Materials and methods
4.1. Genome Sequences of the Sox3 Locus
4.2. Enhancer Screening of the 233-kb Genomic Region and Characterization
4.3. Effector-Dependent Regulation of the NP1 Enhancer
Acknowledgements
References
- Kamachi, Y.; Uchikawa, M.; Collignon, J.; Lovell-Badge, R.; Kondoh, H. Involvement of Sox1, 2 and 3 in the early and subsequent molecular events of lens induction. Development 1998, 125, 2521–2532. [Google Scholar]
- Tanaka, S.; Kamachi, Y.; Tanouchi, A.; Hamada, H.; Jing, N.; Kondoh, H. Interplay of SOX and POU factors in regulation of the Nestin gene in neural primordial cells. Mol. Cell Biol. 2004, 24, 8834–8846. [Google Scholar]
- Inoue, M.; Kamachi, Y.; Matsunami, H.; Imada, K.; Uchikawa, M.; Kondoh, H. PAX6 and SOX2-dependent regulation of the Sox2 enhancer N-3 involved in embryonic visual system development. Genes Cells 2007, 12, 1049–1061. [Google Scholar] [CrossRef]
- Kamachi, Y.; Iwafuchi, M.; Okuda, Y.; Takemoto, T.; Uchikawa, M.; Kondoh, H. Evolution of non-coding regulatory sequences involved in the developmental process: reflection of differential employment of paralogous genes as highlighted by Sox2 and group B1 Sox genes. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2009, 85, 55–68. [Google Scholar] [CrossRef]
- Uchikawa, M.; Yoshida, M.; Iwafuchi-Doi, M.; Matsuda, K.; Ishida, Y.; Takemoto, T.; Kondoh, H. B1 and B2 Sox gene expression during neural plate development in chicken and mouse embryos: Universal versus species-dependent features. Dev. Growth Differ. 2011, 53, 761–771. [Google Scholar] [CrossRef]
- Collignon, J.; Sockanathan, S.; Hacker, A.; Cohen-Tannoudji, M.; Norris, D.; Rastan, S.; Stevanovic, M.; Goodfellow, P.N.; Lovell-Badge, R. A comparison of the properties of Sox-3 with Sry and two related genes, Sox-1 and Sox-2. Development 1996, 122, 509–520. [Google Scholar]
- Wood, H.B.; Episkopou, V. Comparative expression of the mouse Sox1, Sox2 and Sox3 genes from pre-gastrulation to early somite stages. Mech. Dev. 1999, 86, 197–201. [Google Scholar]
- Okuda, Y.; Yoda, H.; Uchikawa, M.; Furutani-Seiki, M.; Takeda, H.; Kondoh, H.; Kamachi, Y. Comparative genomic and expression analysis of group B1 sox genes in zebrafish indicates their diversification during vertebrate evolution. Dev. Dyn. 2006, 235, 811–825. [Google Scholar] [CrossRef]
- Penzel, R.; Oschwald, R.; Chen, Y.; Tacke, L.; Grunz, H. Characterization and early embryonic expression of a neural specific transcription factor xSOX3 in Xenopus laevis. Int. J. Dev. Biol. 1997, 41, 667–477. [Google Scholar]
- Rex, M.; Orme, A.; Uwanogho, D.; Tointon, K.; Wigmore, P.M.; Sharpe, P.T.; Scotting, P.J. Dynamic expression of chicken Sox2 and Sox3 genes in ectoderm induced to form neural tissue. Dev. Dyn. 1997, 209, 323–232. [Google Scholar] [CrossRef]
- Uwanogho, D.; Rex, M.; Cartwright, E.J.; Pearl, G.; Healy, C.; Scotting, P.J.; Sharpe, P.T. Embryonic expression of the chicken Sox2, Sox3 and Sox11 genes suggests an interactive role in neuronal development. Mech. Dev. 1995, 49, 23–36. [Google Scholar] [CrossRef]
- Rogers, C.D.; Archer, T.C.; Cunningham, D.D.; Grammer, T.C.; Casey, E.M. Sox3 expression is maintained by FGF signaling and restricted to the neural plate by Vent proteins in the Xenopus embryo. Dev. Biol. 2008, 313, 307–319. [Google Scholar] [CrossRef]
- Schlosser, G.; Ahrens, K. Molecular anatomy of placode development in Xenopus laevis. Dev. Biol. 2004, 271, 439–466. [Google Scholar] [CrossRef]
- Uchikawa, M.; Kamachi, Y.; Kondoh, H. Two distinct subgroups of Group B Sox genes for transcriptional activators and repressors: Their expression during embryonic organogenesis of the chicken. Mech. Dev. 1999, 84, 103–120. [Google Scholar]
- Nitta, K.R.; Takahashi, S.; Haramoto, Y.; Fukuda, M.; Onuma, Y.; Asashima, M. Expression of Sox1 during Xenopus early embryogenesis. Biochem. Biophys. Res. Commun. 2006, 351, 287–293. [Google Scholar] [CrossRef]
- Okuda, Y.; Ogura, E.; Kondoh, H.; Kamachi, Y. B1 SOX coordinate cell specification with patterning and morphogenesis in the early zebrafish embryo. PLoS Genet. 2010, 6. [Google Scholar] [CrossRef]
- Rizzoti, K.; Lovell-Badge, R. SOX3 activity during pharyngeal segmentation is required for craniofacial morphogenesis. Development 2007, 134, 3437–3448. [Google Scholar]
- Iwafuchi-Doi, M.; Yoshida, Y.; Onichtchouk, D.; Leichsenring, M.; Driever, W.; Takemoto, T.; Uchikawa, M.; Kamachi, Y.; Kondoh, H. The Pou5f1/Pou3f-dependent but SoxB-independent regulation of conserved enhancer N2 initiates Sox2 expression during epiblast to neural plate stages in vertebrates. Dev. Biol. 2011, 352, 354–366. [Google Scholar] [CrossRef]
- Nishiguchi, S.; Wood, H.; Kondoh, H.; Lovell-Badge, R.; Episkopou, V. Sox1 directly regulates the gamma-crystallin genes and is essential for lens development in mice. Genes Dev. 1998, 12, 776–781. [Google Scholar]
- Ekonomou, A.; Kazanis, I.; Malas, S.; Wood, H.; Alifragis, P.; Denaxa, M.; Karagogeos, D.; Constanti, A.; Lovell-Badge, R.; Episkopou, V. Neuronal migration and ventral subtype identity in the telencephalon depend on SOX1. PLoS Biol. 2005, 3. [Google Scholar] [CrossRef] [Green Version]
- Avilion, A.A.; Nicolis, S.K.; Pevny, L.H.; Perez, L.; Vivian, N.; Lovell-Badge, R. Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 2003, 17, 126–140. [Google Scholar] [CrossRef]
- Rizzoti, K.; Brunelli, S.; Carmignac, D.; Thomas, P.Q.; Robinson, I.C.; Lovell-Badge, R. SOX3 is required during the formation of the hypothalamo-pituitary axis. Nat. Genet. 2004, 36, 247–255. [Google Scholar] [CrossRef]
- Weiss, J.; Meeks, J.J.; Hurley, L.; Raverot, G.; Frassetto, A.; Jameson, J.L. Sox3 is required for gonadal function, but not sex determination, in males and females. Mol. Cell Biol. 2003, 23, 8084–8891. [Google Scholar] [CrossRef]
- Graham, V.; Khudyakov, J.; Ellis, P.; Pevny, L. SOX2 functions to maintain neural progenitor identity. Neuron 2003, 39, 749–765. [Google Scholar]
- Pevny, L.H.; Nicolis, S.K. Sox2 roles in neural stem cells. Int. J. Biochem. Cell Biol. 2010, 42, 421–424. [Google Scholar] [CrossRef]
- Dee, C.T.; Hirst, C.S.; Shih, Y.H.; Tripathi, V.B.; Patient, R.K.; Scotting, P.J. Sox3 regulates both neural fate and differentiation in the zebrafish ectoderm. Dev. Biol. 2008, 320, 289–301. [Google Scholar] [CrossRef]
- Uchikawa, M.; Ishida, Y.; Takemoto, T.; Kamachi, Y.; Kondoh, H. Functional analysis of chicken Sox2 enhancers highlights an array of diverse regulatory elements that are conserved in mammals. Dev. Cell 2003, 4, 509–519. [Google Scholar] [CrossRef]
- Okamoto, R.; Uchikawa, M.; Kondoh, H. Twenty-seven enhancers and additional enhancer candidates for Sox2 regulation are distributed within a 200 kb region of the chicken genome. Nucleic Acids Res. 2012. [Google Scholar]
- Takemoto, T.; Uchikawa, M.; Kamachi, Y.; Kondoh, H. Convergence of Wnt and FGF signals in the genesis of posterior neural plate through activation of the Sox2 enhancer N-1. Development 2006, 133, 297–306. [Google Scholar]
- Saigou, Y.; Kamimura, Y.; Inoue, M.; Kondoh, H.; Uchikawa, M. Regulation of Sox2 in the pre-placodal cephalic ectoderm and central nervous system by enhancer N-4. Dev. Growth Differ. 2010, 52, 397–408. [Google Scholar]
- Iwafuchi-Doi, M.; Matsuda, K.; Murakami, K.; Niwa, H.; Tesar, P.; Aruga, J.; Matsuo, I.; Kondoh, H. Transcriptional regulatory networks in epiblast cells and during anterior neural plate development as modeled in epiblast stem cells. Development 2012, 139, 3926–3937. [Google Scholar] [CrossRef]
- Kondoh, H.; Takemoto, T. Axial stem cells deriving both posterior neural and mesodermal tissues during gastrulation. Curr. Opin. Genet. Dev. 2012, 22, 374–380. [Google Scholar] [CrossRef]
- Takemoto, T.; Uchikawa, M.; Yoshida, M.; Bell, D.M.; Lovell-Badge, R.; Papaioannou, V.E.; Kondoh, H. Tbx6-dependent Sox2 regulation determines neural or mesodermal fate in axial stem cells. Nature 2011, 470, 394–398. [Google Scholar] [CrossRef]
- van Rooijen, C.; Simmini, S.; Bialecka, M.; Neijts, R.; van de Ven, C.; Beck, F.; Deschamps, J. Evolutionarily conserved requirement of Cdx for post-occipital tissue emergence. Development 2012, 139, 2576–2583. [Google Scholar] [CrossRef]
- Brunelli, S.; Silva Casey, E.; Bell, D.; Harland, R.; Lovell-Badge, R. Expression of Sox3 throughout the developing central nervous system is dependent on the combined action of discrete, evolutionarily conserved regulatory elements. Genesis 2003, 36, 12–24. [Google Scholar] [CrossRef]
- Kondoh, H.; Uchikawa, M. Dissection of chick genomic regulatory regions. Meth. Cell Biol. 2008, 87, 313–336. [Google Scholar]
- Kovacevic Grujicic, N.; Mojsin, M.; Krstic, A.; Stevanovic, M. Functional characterization of the human SOX3 promoter: identification of transcription factors implicated in basal promoter activity. Gene 2005, 344, 287–297. [Google Scholar] [CrossRef]
- Krstic, A.; Mojsin, M.; Stevanovic, M. Regulation of SOX3 gene expression is driven by multiple NF-Y binding elements. Arch. Biochem. Biophys. 2007, 467, 163–173. [Google Scholar] [CrossRef]
- Mojsin, M.; Stevanovic, M. PBX1 and MEIS1 up-regulate SOX3 gene expression by direct interaction with a consensus binding site within the basal promoter region. Biochem. J. 2009, 425, 107–116. [Google Scholar]
- Sawicki, J.A.; Morris, R.J.; Monks, B.; Sakai, K.; Miyazaki, J. A composite CMV-IE enhancer/beta-actin promoter is ubiquitously expressed in mouse cutaneous epithelium. Exp. Cell Res. 1998, 244, 367–369. [Google Scholar] [CrossRef]
Supplementary Materials


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chicken Sox3 locus | Human Sox3 locus | Mouse Sox3 locus | Xenopus Sox3 locus | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| CSB | Positionsa | Length (bp) | Positionsa | Length (bp) | Human vs. chicken Identity (%) | Positionsa | Length (bp) | Mouse vs. chicken Identity (%) | Positionsa | Length (bp) | Xenopus vs. chicken Identity (%) |
| 1 | -130257 ~ -130129 | 129 | -970154 ~ -970026 | 129 | 84c | -589181 ~ -589053 | 129 | 82 | Absent | ||
| 2 | -110940 ~ -110700 | 241 | -444400 ~ -444161 | 240 | 73 | -522083 ~ -521837 | 247 | 74 | Absent | ||
| 3 | -108306 ~ -107937 | 370 | -428513 ~ -428153 | 361 | 71 | -500213 ~ -499841 | 373 | 69 | -129767 ~ -129393 | 375 | 62 |
| 4 | -103688 ~ -103557 | 132 | Absent | -474215 ~ -474091 | 125 | 60 | -121856 ~ -121723 | 134 | 64 | ||
| 5 | -97501 ~ -97400 | 102 | Absent | -388001 ~ -387900 | 102 | 62 | Absent | ||||
| 6 | -82740 ~ -82535 | 206 | -274707 ~ -274502 | 206 | 66 | -286133 ~ -285930 | 204 | 53d | Absent | ||
| 7 | -81558 ~ -81458 | 101 | Absent | -297179 ~ -297065 | 115 | 60 | Absent | ||||
| 8 | -67333 ~ -67221 | 113 | -252300 ~ -252193 | 108 | 57d | -261781 ~ -261669 | 113 | 63 | Absent | ||
| 9 | -60650 ~ -60531 | 120 | -246870 ~ -246750 | 121 | 64 | -253372 ~ -253247 | 126 | 64 | Absent | ||
| 10 | -59303 ~ -59126 | 178 | -243592 ~ -243410 | 183 | 63 | Absent | Absent | ||||
| 11 | -56180 ~ -56079 | 102 | Absent | -230997 ~ -230897 | 101 | 61 | Absent | ||||
| 12 | -45556 ~ -44996 | 561 | -156675 ~ -156165 | 511 | 75 | -149792 ~ -149283 | 510 | 75 | -42634 ~ -42145 | 490 | 62 |
| 13 | -29736 ~ -29259 | 478 | -137259 ~ -136775 | 485 | 79 | -133617 ~ -133151 | 467 | 80 | -31421 ~ -30964 | 458 | 60 |
| 14 | -26388 ~ -25483 | 906 | -88401 ~ -87521 | 881 | 78 | -101763 ~ -100874 | 890 | 79 | -27373 ~ -26479 | 895 | 67 |
| 15 | -21081 ~ -20662 | 420 | -34050 ~ -33637 | 414 | 77 | -37248 ~ -36842 | 407 | 77 | -19657 ~ -19236 | 422 | 65 |
| 16 | -17784 ~ -17128 | 657 | -28051 ~ -27389 | 663 | 82 | -25441 ~ -24780 | 662 | 81 | -18039 ~ -17396 | 644 | 61 |
| 17 | -14950 ~ -14611 | 340 | -23177 ~ -22843 | 335 | 76 | -22166 ~ -21840 | 327 | 69 | -15729 ~ -15389 | 341 | 60 |
| 18 | -13560 ~ -13273 | 288 | -22619 ~ -22337 | 283 | 65 | -21624 ~ -21340 | 285 | 63 | Absent | ||
| 19 | -11574 ~ -11339 | 236 | -21444 ~ -21219 | 226 | 63 | -20545 ~ -20316 | 230 | 51d | -13988 ~ -13757 | 232 | 71 |
| 20 | -5363 ~ -4690 | 674 | -7397 ~ -6739 | 659 | 77 | -6672 ~ -6014 | 659 | 76 | -7241 ~ -6571 | 671 | 60 |
| 21 | -4176 ~ -4004 | 173 | Absent | Absent | -5417 ~ -5246 | 172 | 64 | ||||
| 22 | -944 ~ -792 | 153 | -1258 ~ -1099 | 160 | 65 | -1253 ~ -1097 | 157 | 65 | Absent | ||
| 23 | -88 ~ 997 | 1086 | -87 ~ 1166 | 1254 | 67 | -87 ~ 1177 | 1265 | 67 | -88 ~ 969 | 1058 | 71 |
| Sox3 CDSb | -225 ~ 1116 | 1342 | -225 ~ 1128 | 1354 | |||||||
| Sox3 CDSa | 1 ~ 951 | 951 | 1 ~ 1116 | 1116 | 66 | 1 ~ 1128 | 1128 | 68 | 1 ~ 924 | 924 | 76 |
| 24 | 1982 ~ 2098 | 117 | 2253 ~ 2378 | 126 | 68 | 2247 ~ 2369 | 123 | 64 | Absent | ||
| 25 | 20709 ~ 20882 | 174 | 48743 ~ 48914 | 172 | 67 | 45466 ~ 45638 | 173 | 67 | Absent | ||
| 26 | 24291 ~ 24916 | 626 | 54918 ~ 55541 | 624 | 70 | 51461 ~ 52097 | 637 | 70 | 22831 ~ 23470 | 640 | 70 |
| 27 | 29103 ~ 29471 | 369 | 64927 ~ 65286 | 360 | 63 | 61426 ~ 61785 | 360 | 61 | 24237 ~ 24595 | 359 | 76 |
| 28 | 33725 ~ 33839 | 115 | Absent | Absent | 28864 ~ 28991 | 128 | 63 | ||||
| 29 | 35797 ~ 35915 | 119 | 114633 ~ 114750 | 118 | 61 | 93771 ~ 93888 | 118 | 61 | 31915 ~ 32036 | 122 | 61 |
| 30 | 37839 ~ 37943 | 105 | Absent | Absent | 50701 ~ 50801 | 101 | 69 | ||||
| 31 | 43240 ~ 43490 | 251 | 146259 ~ 146505 | 247 | 60 | 115905 ~ 116157 | 253 | 60 | Absent | ||
| 32 | 47105 ~ 47964 | 860 | 205211 ~ 206065 | 855 | 79 | 148542 ~ 149384 | 843 | 76 | 59011 ~ 59847 | 837 | 63 |
| 33 | 48341 ~ 48713 | 373 | 206395 ~ 206764 | 370 | 72 | 149726 ~ 150095 | 370 | 72 | 61798 ~ 62162 | 365 | 60 |
| 34 | 52194 ~ 52648 | 455 | 253981 ~ 254434 | 454 | 86 | 173018 ~ 173471 | 454 | 86 | 66646 ~ 67075 | 430 | 71 |
| 35 | 60282 ~ 60540 | 259 | 314971 ~ 315223 | 253 | 67 | 212815 ~ 213069 | 255 | 69 | 82061 ~ 82322 | 262 | 56d |
| 36 | 69025 ~ 69166 | 142 | 325798 ~ 325935 | 138 | 61 | 235698 ~ 235833 | 136 | 62 | Absent | ||
| 37 | 69205 ~ 69439 | 235 | 325967 ~ 326202 | 236 | 61 | 235872 ~ 236110 | 239 | 60 | Absent | ||
| 38 | 71699 ~ 72240 | 542 | 346769 ~ 347310 | 542 | 82 | 262559 ~ 263101 | 543 | 73 | 93894 ~ 94430 | 537 | 65 |
| 39 | 75210 ~ 75377 | 168 | 376498 ~ 376664 | 167 | 84c | Absent | 95795 ~ 95968 | 174 | 71 | ||
| 40 | 77117 ~ 77242 | 126 | 393019 ~ 393146 | 128 | 69 | 322436 ~ 322565 | 130 | 60 | Absent | ||
| 41 | 77293 ~ 77414 | 122 | 393172 ~ 393292 | 121 | 62 | 322599 ~ 322709 | 111 | 50d | Absent | ||
| Sequence number | Positions relative to the Sox3 start site (bp) | Sequence length (bp) | Enhancer activity in the CNS (HH stagesa) | Non-CNS enhancer activity (HH stagesa) | Analyzed specimens |
|---|---|---|---|---|---|
| U28 | -134079 ~ -129036 | 5044 | - | - | 3 |
| U27 | -130087 ~ -125081 | 5007 | - | - | 7 |
| U26 | -126128 ~ -121082 | 5047 | - | - | 8 |
| U25 | -122088 ~ -117083 | 5006 | - | - | 3 |
| U24 | -118083 ~ -113121 | 4963 | - | - | 4 |
| U23 | -114084 ~ -109748 | 4337 | - | - | 4 |
| U22 | -110784 ~ -106313 | 4472 | - | - | 5 |
| U21 | -107300 ~ -102342 | 4959 | - | - | 9 |
| U20 | -103342 ~ -98324 | 5019 | - | - | 5 |
| U19 | -99343 ~ -94344 | 5000 | - | - | 6 |
| U18 | -95344 ~ -90289 | 5056 | - | - | 4 |
| U17 | -91345 ~ -86346 | 5000 | - | - | 4 |
| U16 | -87346 ~ -82309 | 5038 | - | - | 6 |
| U15 | -83347 ~ -78332 | 5016 | - | - | 8 |
| U14 | -79348 ~ -74349 | 5000 | - | - | 2 |
| U13 | -75351 ~ -70345 | 5007 | - | - | 3 |
| U12 | -71350 ~ -66311 | 5040 | - | - | 3 |
| U11 | -67351 ~ -62352 | 5000 | - | - | 4 |
| U10 | -63400 ~ -58258 | 5143 | - | - | 5 |
| U9 | -59365 ~ -54312 | 5054 | - | - | 3 |
| U8 | -55353 ~ -50146 | 5208 | - | - | 6 |
| U7 | -51354 ~ -46353 | 5002 | - | - | 9 |
| U6 | -47369 ~ -42356 | 5014 | - | - | 8 |
| U5 | -43364 ~ -38357 | 5008 | - | - | 3 |
| U4 | -39363 ~ -34454 | 4910 | - | - | 3 |
| U3 | -35550 ~ -30442 | 5109 | - | Epibranchial placode (st. 9) | 13 |
| U2 | -31484 ~ -26456 | 5029 | - | - | 3 |
| U1 | -27456 ~ -22435 | 5022 | - | - | 5 |
| C1 | -23456 ~ -16912 | 6545 | - | - | 4 |
| C2 | -22029 ~ -14489 | 7541 | - | - | 7 |
| C3 | -16915 ~ -12683 | 4233 | - | - | 4 |
| C4 | -16353 ~ -10686 | 5668 | - | Lens placode(st. 11) | 4 |
| C5 | -13647 ~ -9325 | 4323 | - | Lens placode(st. 11) | 6 |
| C6 | -10822 ~ -6096 | 4727 | - | - | 2 |
| C7 | -7397 ~ 2548 | 9945 | - | - | 5 |
| C8 | -2027 ~ 6462 | 8489 | - | - | 5 |
| C9 | 4748 ~ 15637 | 10890 | - | - | 5 |
| C10 | 6459 ~ 11599 | 5141 | - | - | 4 |
| C11 | 13024 ~ 21441 | 8418 | - | - | 3 |
| C12 | 16899 ~ 29642 | 12744 | Diencephalon, spinal cord (st.11) | - | 8 |
| C13 | 21752 ~ 28244 | 6493 | Diencephalon, spinal cord (st.11) | - | 8 |
| D1 | 28860 ~ 33859 | 5000 | Posterior neural plate, rhombencephalon, spinal cord (st. 5) | Otic placode (st. 11) | 15 |
| D2 | 32859 ~ 37858 | 5000 | - | - | 3 |
| D3 | 36858 ~ 41867 | 5010 | - | Otic placode (st. 11) | 11 |
| D4 | 40830 ~ 45856 | 5027 | Telencephalon (st. 9) | - | 11 |
| D5 | 44847 ~ 49855 | 5009 | Tel/di/mesencephalon (st. 9) | Lens placode (st. 11) | 12 |
| D6 | 48830 ~ 53858 | 5029 | Mes/rhombencephalon, spinal cord (st. 15) | - | 8 |
| D7 | 52798 ~ 57856 | 5059 | - | - | 3 |
| D8 | 56853 ~ 61861 | 5009 | - | Epibranchial placode (st. 9) | 7 |
| D9 | 60834 ~ 65872 | 5039 | - | Lens (st. 17)b | 3 |
| D10 | 64835 ~ 69850 | 5016 | Ventral diencephalon (st. 13)b | Otic vesicle, nasal pit (st. 13)b | 3 |
| D11 | 68843 ~ 73849 | 5007 | Spinal cord (st. 9) | - | 10 |
| D12 | 72849 ~ 77848 | 5000 | Diencephalon (st. 11) | - | 11 |
| D13 | 76848 ~ 81847 | 5000 | Posterior neural plate, di/rhombencephalon, spinal cord (st. 8) | - | 12 |
| D14 | 80847 ~ 85895 | 5049 | - | - | 3 |
| D15 | 84850 ~ 88760 | 3911 | CNS (st. 8) | - | 3 |
| D16 | 88029 ~ 93228 | 5200 | - | - | 3 |
| D17 | 92231 ~ 97230 | 5000 | - | - | 5 |
| CSB19 | -12377 ~ -10964 | 1414 | - | Lens placode (st. 11) | 3 |
| CSB26 | 24264 ~ 25312 | 1049 | Diencephalon, spinal cord (st. 11) | - | 5 |
| CSB27 | 28179 ~ 29859 | 1681 | Posterior neural plate, rhombencephalon, spinal cord (st. 5) | Otic placode (st. 11) | 37 |
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Nishimura, N.; Kamimura, Y.; Ishida, Y.; Takemoto, T.; Kondoh, H.; Uchikawa, M. A Systematic Survey and Characterization of Enhancers that Regulate Sox3 in Neuro-Sensory Development in Comparison with Sox2 Enhancers. Biology 2012, 1, 714-735. https://doi.org/10.3390/biology1030714
Nishimura N, Kamimura Y, Ishida Y, Takemoto T, Kondoh H, Uchikawa M. A Systematic Survey and Characterization of Enhancers that Regulate Sox3 in Neuro-Sensory Development in Comparison with Sox2 Enhancers. Biology. 2012; 1(3):714-735. https://doi.org/10.3390/biology1030714
Chicago/Turabian StyleNishimura, Naoko, Yoshifumi Kamimura, Yoshiko Ishida, Tatsuya Takemoto, Hisato Kondoh, and Masanori Uchikawa. 2012. "A Systematic Survey and Characterization of Enhancers that Regulate Sox3 in Neuro-Sensory Development in Comparison with Sox2 Enhancers" Biology 1, no. 3: 714-735. https://doi.org/10.3390/biology1030714
APA StyleNishimura, N., Kamimura, Y., Ishida, Y., Takemoto, T., Kondoh, H., & Uchikawa, M. (2012). A Systematic Survey and Characterization of Enhancers that Regulate Sox3 in Neuro-Sensory Development in Comparison with Sox2 Enhancers. Biology, 1(3), 714-735. https://doi.org/10.3390/biology1030714