Smallholder Farmers’ Practices and African Indigenous Vegetables Affect Soil Microbial Biodiversity and Enzyme Activities in Lake Naivasha Basin, Kenya

 ,
,  , , ,
, , ,  ,
,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
- bacterial and fungal biodiversity;
- enzymatic activity; and
- soils alteration status when compared to mainstream farming approaches such as “maize monoculture” or “maize and beans.”
2. Materials and Methods
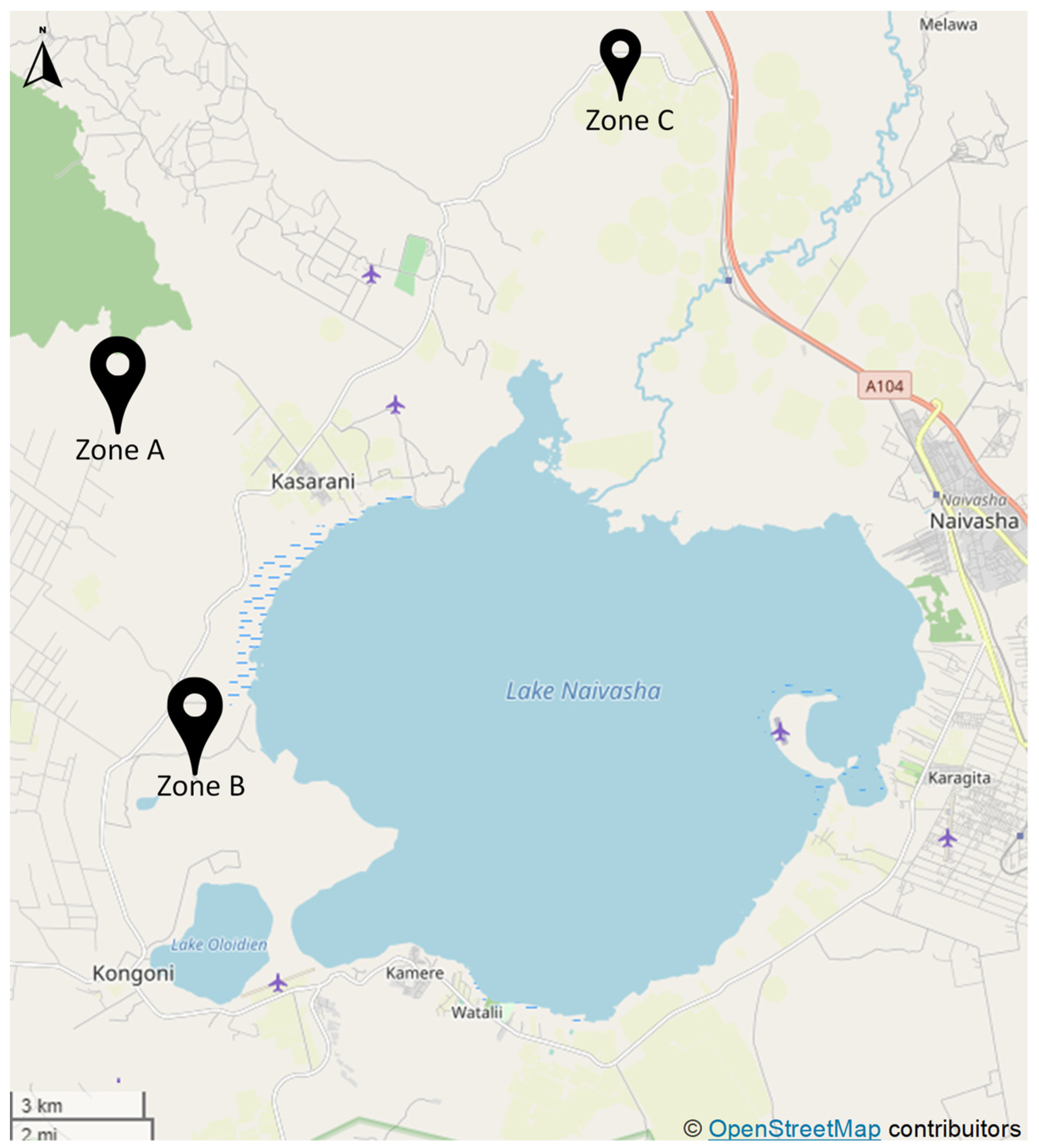
2.1. Site Description, Survey, and Soil Sampling
2.2. Analyses of Soil Microbial Diversity
2.3. Soil Enzymatic Activity
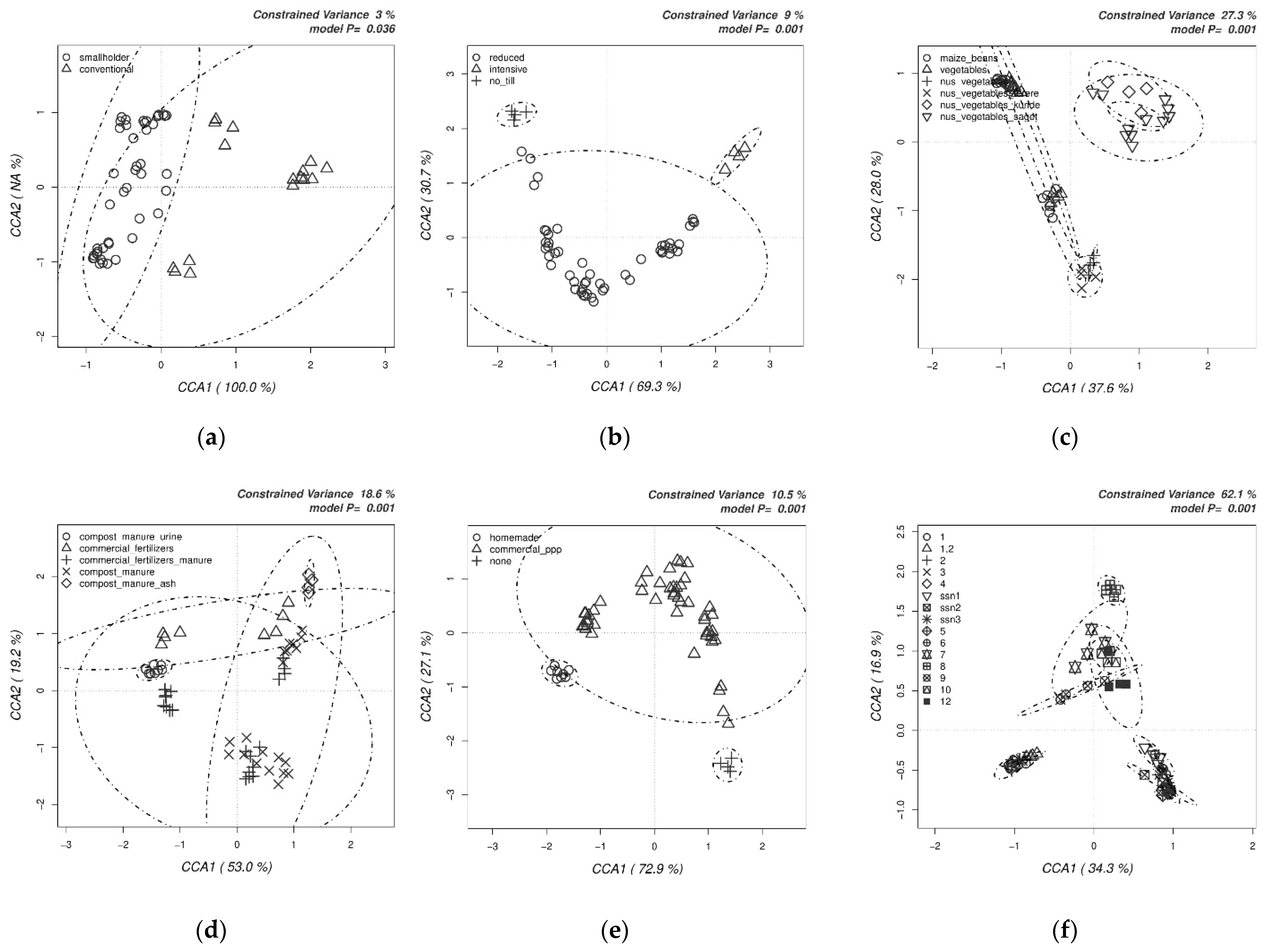
2.4. Statistical Analyses
3. Results
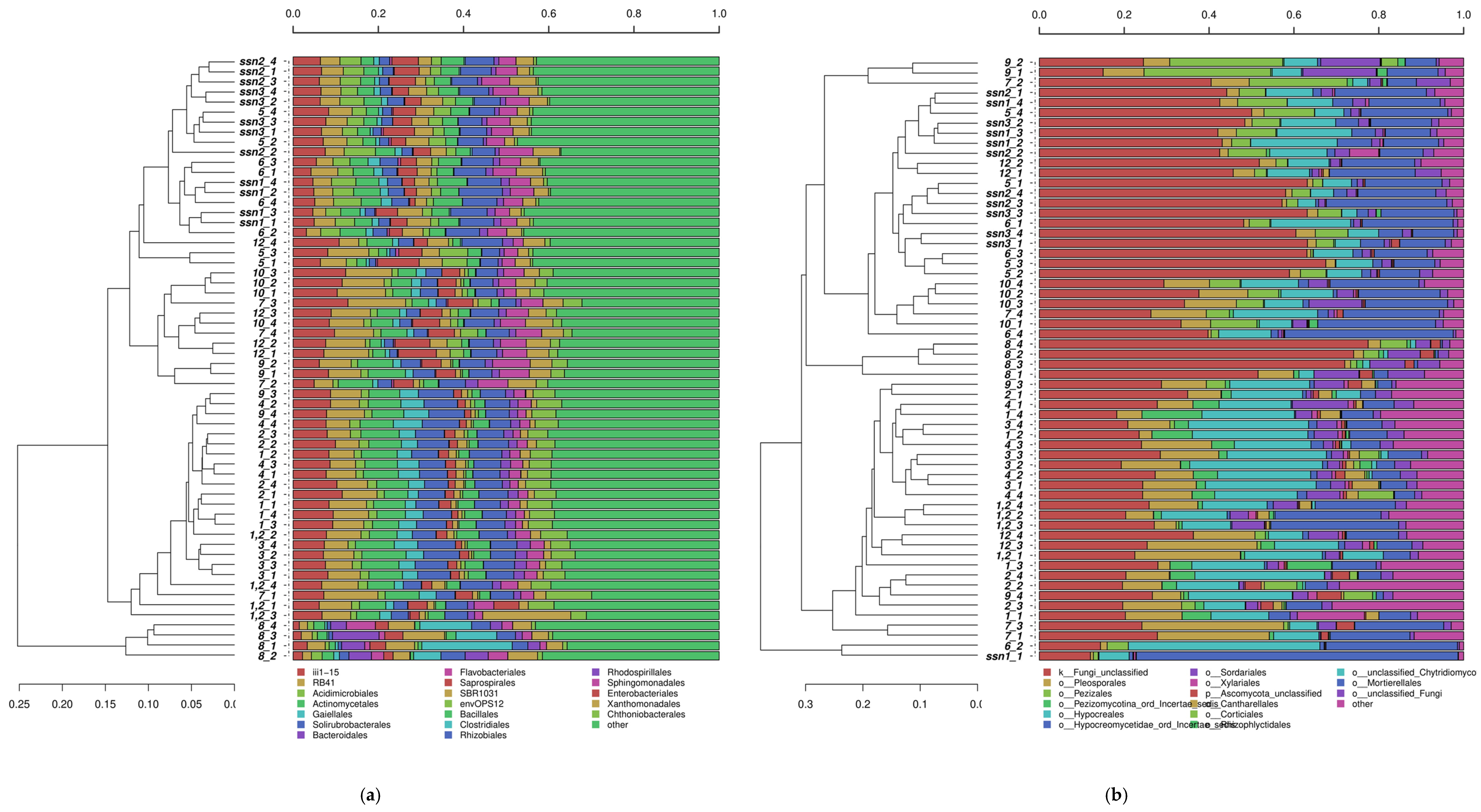
3.1. Analyses of Soil Microbial Diversity
3.2. Soil Enzymatic Activity
4. Discussion
4.1. Biodiversity of Bacterial and Fungal Communities
4.2. Enzymatic Activity
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tully, K.; Sullivan, C.; Weil, R.; Sanchez, P. The State of Soil Degradation in Sub-Saharan Africa: Baselines, Trajectories, and Solutions. Sustainability 2015, 7, 6523–6552. [Google Scholar] [CrossRef] [Green Version]
- Tittonell, P.; Giller, K.E. When yield gaps are poverty traps: The paradigm of ecological intensification in African smallholder agriculture. Field Crop. Res. 2013, 143, 76–90. [Google Scholar] [CrossRef] [Green Version]
- Mugo, J.N.; Karanja, N.N.; Gachene, C.K.; Dittert, K.; Nyawade, S.O.; Schulte-Geldermann, E. Assessment of soil fertility and potato crop nutrient status in central and eastern highlands of Kenya. Sci. Rep. 2020, 10, 7779. [Google Scholar] [CrossRef] [PubMed]
- Knox, J.; Hess, T.; Daccache, A.; Wheeler, T. Climate change impacts on crop productivity in Africa and South Asia. Environ. Res. Lett. 2012, 7. [Google Scholar] [CrossRef]
- Ringler, C.; Zhu, T.; Cai, X.; Koo, J.; Wang, D. Climate Change Impacts on Food Security in Sub-Saharan Africa; International Food Policy Research Institute: Washington, DC, USA, 2010. [Google Scholar]
- Wolfram, S.; David, B.L. Robust negative impacts of climate change on African agriculture. Environ. Res. Lett. 2010, 5, 014010. [Google Scholar]
- Kenya and FAO: Partnering to Build Resilience and Food and Nutrition Security; Food and Agriculture Organization: Rome, Italy, 2017.
- FAOSTAT. Kenya: FAO Country Profile. Available online: http://countrystat.org/home.aspx?c=KEN&p=co (accessed on 15 May 2020).
- FAO; IFAD; UNICEF; WFP; WHO. Transforming Food Systems for Affordable Healthy Diets; Food and Agriculture Organization: Rome, Italy, 2020. [Google Scholar]
- Dixon, J.A.; Gibbon, D.P.; Gulliver, A.; Hall, M. Farming Systems and Poverty: Improving Farmers’ Livelihoods in a Changing World; FAO: Rome, Italy; World Bank: Washington, DC, USA, 2001. [Google Scholar]
- Droppelmann, K.J.; Snapp, S.S.; Waddington, S.R. Sustainable intensification options for smallholder maize-based farming systems in sub-Saharan Africa. Food Secur. 2017, 9, 133–150. [Google Scholar] [CrossRef]
- Campanaro, A.; Tommasi, N.; Guzzetti, L.; Galimberti, A.; Bruni, I.; Labra, M. DNA barcoding to promote social awareness and identity of neglected, underutilized plant species having valuable nutritional properties. Food Res. Int. 2018. [Google Scholar] [CrossRef] [PubMed]
- Baldermann, S.; Blagojević, L.; Frede, K.; Klopsch, R.; Neugart, S.; Neumann, A.; Ngwene, B.; Norkeweit, J.; Schröter, D.; Schröter, A.; et al. Are Neglected Plants the Food for the Future? Crit. Rev. Plant Sci. 2016, 35, 106–119. [Google Scholar] [CrossRef] [Green Version]
- Pichop, G.N.; Abukutsa-Onyango, M.; Noorani, A.; Nono-Womdim, R. Importance of indigenous food crops in tropical Africa: Case study. Acta Hortic 2016. [Google Scholar] [CrossRef]
- Ojiewo, C.; Tenkouano, A.; Hughes, J.D.A.; Keatinge, J.D.H. Diversifying diets: Using indigenous vegetables to improve profitability, nutrition and health in Africa. In Diversifying Food and Diets: Using Agricultural Biodiversity to Improve Nutrition and Health; Taylor & Francis Group: Abingdon, UK, 2013; pp. 291–302. [Google Scholar]
- Ojiewo, C.O.; Rubyogo, J.C.; Wesonga, J.M.; Bishaw, Z.; Gelalcha, S.W.; Abang, M.M. Mainstreaming Efficient Legume Seed Systems in Eastern Africa: Challenges, Opportunities and Contributions Towards Improved Livelihoods; Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Guzzetti, L.; Fiorini, A.; Panzeri, D.; Tommasi, N.; Grassi, F.; Taskin, E.; Misci, C.; Puglisi, E.; Tabaglio, V.; Galimberti, A.; et al. Sustainability Perspectives of Vigna unguiculata L. Walp. Cultivation under No Tillage and Water Stress Conditions. Plants 2020, 9, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taskin, E.; Boselli, R.; Fiorini, A.; Misci, C.; Ardenti, F.; Bandini, F.; Guzzetti, L.; Panzeri, D.; Tommasi, N.; Galimberti, A.; et al. Combined Impact of No-Till and Cover Crops with or without Short-Term Water Stress as Revealed by Physicochemical and Microbiological Indicators. Biology 2021, 10, 23. [Google Scholar] [CrossRef] [PubMed]
- Mugizi, F.M.P.; Matsumoto, T. Population pressure and soil quality in Sub-Saharan Africa: Panel evidence from Kenya. Land Use Policy 2020, 94, 104499. [Google Scholar] [CrossRef]
- Pretty, J.; Toulmin, C.; Williams, S. Sustainable intensification in African agriculture. Int. J. Agric. Sustain. 2011, 9, 5–24. [Google Scholar] [CrossRef]
- von Arb, C.; Bünemann, E.K.; Schmalz, H.; Portmann, M.; Adamtey, N.; Musyoka, M.W.; Frossard, E.; Fliessbach, A. Soil quality and phosphorus status after nine years of organic and conventional farming at two input levels in the Central Highlands of Kenya. Geoderma 2020, 362, 114112. [Google Scholar] [CrossRef]
- Nyberg, Y.; Wetterlind, J.; Jonsson, M.; Öborn, I. The role of trees and livestock in ecosystem service provision and farm priorities on smallholder farms in the Rift Valley, Kenya. Agric. Syst. 2020, 181, 102815. [Google Scholar] [CrossRef]
- De Mastro, F.; Traversa, A.; Brunetti, G.; Debiase, G.; Cocozza, C.; Nigro, F. Soil culturable microorganisms as affected by different soil managements in a two year wheat-faba bean rotation. Appl. Soil Ecol. 2020, 149. [Google Scholar] [CrossRef]
- Mulinge, W.; Gicheru, P.; Murithi, F.; Maingi, P.; Kihiu, E.; Kirui, O.K.; Mirzabaev, A. Economics of Land Degradation and Improvement in Kenya. In Economics of Land Degradation and Improvement—A Global Assessment for Sustainable Development; Nkonya, E., Mirzabaev, A., von Braun, J., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 471–498. [Google Scholar]
- Fischer, S.; Hilger, T.; Piepho, H.P.; Jordan, I.; Karungi, J.; Towett, E.; Shepherd, K.; Cadisch, G. Soil and farm management effects on yield and nutrient concentrations of food crops in East Africa. Sci. Total Environ. 2020, 716, 137078. [Google Scholar] [CrossRef]
- Kihara, J.; Bolo, P.; Kinyua, M.; Nyawira, S.S.; Sommer, R. Soil health and ecosystem services: Lessons from sub-Sahara Africa (SSA). Geoderma 2020, 370, 114342. [Google Scholar] [CrossRef]
- Nyawade, S.O.; Gachene, C.K.K.; Karanja, N.N.; Gitari, H.I.; Schulte-Geldermann, E.; Parker, M.L. Controlling soil erosion in smallholder potato farming systems using legume intercrops. Geoderma Reg. 2019, 17, e00225. [Google Scholar] [CrossRef]
- Njoroge, S.; Schut, A.G.T.; Giller, K.E.; Zingore, S. Learning from the soil’s memory: Tailoring of fertilizer application based on past manure applications increases fertilizer use efficiency and crop productivity on Kenyan smallholder farms. Eur. J. Agron. 2019, 105, 52–61. [Google Scholar] [CrossRef]
- Kiboi, M.N.; Ngetich, K.F.; Fliessbach, A.; Muriuki, A.; Mugendi, D.N. Soil fertility inputs and tillage influence on maize crop performance and soil water content in the Central Highlands of Kenya. Agric. Water Manag. 2019, 217, 316–331. [Google Scholar] [CrossRef]
- Jena, P.R. Can minimum tillage enhance productivity? Evidence from smallholder farmers in Kenya. J. Clean. Prod. 2019, 218, 465–475. [Google Scholar] [CrossRef]
- De Mastro, F.; Traversa, A.; Cocozza, C.; Pallara, M.; Brunetti, G. Soil organic carbon stabilization: Influence of tillage on mineralogical and chemical parameters. Soil Syst. 2020, 4, 58. [Google Scholar] [CrossRef]
- Willy, D.K.; Muyanga, M.; Mbuvi, J.; Jayne, T. The effect of land use change on soil fertility parameters in densely populated areas of Kenya. Geoderma 2019, 343, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Cobo, J.G.; Dercon, G.; Cadisch, G. Nutrient balances in African land use systems across different spatial scales: A review of approaches, challenges and progress. Agric. Ecosyst. Environ. 2010, 136, 1–15. [Google Scholar] [CrossRef]
- Baraniya, D.; Puglisi, E.; Ceccherini, M.T.; Pietramellara, G.; Giagnoni, L.; Arenella, M.; Nannipieri, P.; Renella, G. Protease encoding microbial communities and protease activity of the rhizosphere and bulk soils of two maize lines with different N uptake efficiency. Soil Biol. Biochem. 2016, 96, 176–179. [Google Scholar] [CrossRef]
- Delgado Baquerizo, M.; Maestre, F.T.; Reich, P.B.; Jeffries, T.C.; Gaitan, J.J.; Encinar, D.; Berdugo, M.; Campbell, C.D.; Singh, B.K. Microbial diversity drives multifunctionality in terrestrial ecosystems. Nat. Commun. 2016, 7, 10541. [Google Scholar] [CrossRef] [Green Version]
- Jansson, J.K.; Hofmockel, K.S. The soil microbiome-from metagenomics to metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Cocconcelli, P.S.; Trevisan, M. Soil bacterial diversity screening using single 16S rRNA gene V regions coupled with multi-million read generating sequencing technologies. PLoS ONE 2012, 7. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2017, 68, 12–26. [Google Scholar] [CrossRef]
- Chepkoech, W.; Mungai, N.W.; Stöber, S.; Lotze-Campen, H. Understanding adaptive capacity of smallholder African indigenous vegetable farmers to climate change in Kenya. Clim. Risk Manag. 2020, 27, 100204. [Google Scholar] [CrossRef]
- Trivedi, P.; Delgado-Baquerizo, M.; Anderson, I.C.; Singh, B.K. Response of Soil Properties and Microbial Communities to Agriculture: Implications for Primary Productivity and Soil Health Indicators. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloter, M.; Nannipieri, P.; Sørensen, S.J.; van Elsas, J.D. Microbial indicators for soil quality. Biol. Fertil. Soil 2018, 54, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nannipieri, P.; Trasar-Cepeda, C.; Dick, R.P. Soil enzyme activity: A brief history and biochemistry as a basis for appropriate interpretations and meta-analysis. Biol. Fertil. Soil 2018, 54, 11–19. [Google Scholar] [CrossRef]
- Vischetti, C.; Casucci, C.; De Bernardi, A.; Monaci, E.; Tiano, L.; Marcheggiani, F.; Ciani, M.; Comitini, F.; Marini, E.; Taskin, E.; et al. Sub-Lethal Effects of Pesticides on the DNA of Soil Organisms as Early Ecotoxicological Biomarkers. Front. Microbiol. 2020, 11. [Google Scholar] [CrossRef]
- Burns, R.G.; DeForest, J.L.; Marxsen, J.; Sinsabaugh, R.L.; Stromberger, M.E.; Wallenstein, M.D.; Weintraub, M.N.; Zoppini, A. Soil enzymes in a changing environment: Current knowledge and future directions. Soil Biol. Biochem. 2013, 58, 216–234. [Google Scholar] [CrossRef]
- Hartman, K.; Tringe, S.G. Interactions between plants and soil shaping the root microbiome under abiotic stress. Biochem. J. 2019, 476, 2705–2724. [Google Scholar] [CrossRef] [Green Version]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The biological activities of β-glucosidase, phosphatase and urease as soil quality indicators: A review. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef] [Green Version]
- Puglisi, E.; Del Re, A.A.M.; Rao, M.A.; Gianfreda, L. Development and validation of numerical indexes integrating enzyme activities of soils. Soil Biol. Biochem. 2006, 38, 1673–1681. [Google Scholar] [CrossRef]
- Wanjiku Kamau, J.; Biber-Freudenberger, L.; Lamers, J.P.A.; Stellmacher, T.; Borgemeister, C. Soil fertility and biodiversity on organic and conventional smallholder farms in Kenya. Appl. Soil Ecol. 2019, 134, 85–97. [Google Scholar] [CrossRef]
- Jaetzold, R.; Schmidt, H.; Hornetz, B.; Shisanya, C.; Agriculture, M.O.; Zusammenarbeit, D.G.F.T. Farm Management Handbook of Kenya: Volume II: Natural Conditions and Farm Management Information; Subpart B1a: Southern Rift Valley Province Nakuru County; Ministry of Agriculture: Nairobi, Kenya, 2010.
- Soil Quality—Sampling—Part 102: Selection and Application of Sampling Techniques; 18400–102:2017; International Organization for Standardization: Geneva, Switzerland, 2017.
- Vasileiadis, S.; Puglisi, E.; Trevisan, M.; Scheckel, K.G.; Langdon, K.A.; McLaughlin, M.J.; Lombi, E.; Donner, E. Changes in soil bacterial communities and diversity in response to long-term silver exposure. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandini, F.; Misci, C.; Taskin, E.; Cocconcelli, P.S.; Puglisi, E. Biopolymers modulate microbial communities in municipal organic waste digestion. FEMS Microbiol. Ecol. 2020, 96. [Google Scholar] [CrossRef] [PubMed]
- Połka, J.; Rebecchi, A.; Pisacane, V.; Morelli, L.; Puglisi, E. Bacterial diversity in typical Italian salami at different ripening stages as revealed by high-throughput sequencing of 16S rRNA amplicons. Food Microbiol. 2015, 46, 342–356. [Google Scholar] [CrossRef] [PubMed]
- Vasileiadis, S.; Puglisi, E.; Arena, M.; Cappa, F.; Veen, J.A.; Cocconcelli, P.S.; Trevisan, M. Soil microbial diversity patterns of a lowland spring environment. FEMS Microbiol. Ecol. 2013, 86, 172–184. [Google Scholar] [CrossRef] [Green Version]
- Masella, A.P.; Bartram, A.K.; Truszkowski, J.M.; Brown, D.G.; Neufeld, J.D. PANDAseq: Paired-end assembler for illumina sequences. BMC Bioinform. 2012, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J. Introducing mothur: Open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Pruesse, E.; Quast, C.; Knittel, K.; Fuchs, B.M.; Ludwig, W.; Peplies, J.; Glöckner, F.O. SILVA: A comprehensive online resource for quality checked and aligned ribosomal RNA sequence data compatible with ARB. Nucleic Acids Res. 2007, 35, 7188–7196. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Keller, K.; Brodie, E.L.; Larsen, N.; Piceno, Y.M.; Phan, R.; Andersen, G.L. NAST: A multiple sequence alignment server for comparative analysis of 16S rRNA genes. Nucleic Acids Res. 2006, 34, W394–W399. [Google Scholar] [CrossRef]
- Schloss, P.D. The effects of alignment quality, distance calculation method, sequence filtering, and region on the analysis of 16S rRNA gene-based studies. PLoS Comput. Biol. 2010, 6, e1000844. [Google Scholar] [CrossRef] [Green Version]
- McDonald, D.; Price, M.N.; Goodrich, J.; Nawrocki, E.P.; DeSantis, T.Z.; Probst, A.; Andersen, G.L.; Knight, R.; Hugenholtz, P. An improved Greengenes taxonomy with explicit ranks for ecological and evolutionary analyses of bacteria and archaea. ISME J. 2011, 6, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Eivazi, F.; Tabatabai, M.A. Factors affecting glucosidase and galactosidase activities in soils. Soil Biol. Biochem. 1990, 22, 891–897. [Google Scholar] [CrossRef]
- Sannino, F.; Gianfreda, L. Pesticide influence on soil enzymatic activities. Chemosphere 2001, 45, 417–425. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-term assay of soil urease activity using colorimetric determination of ammonium. Biol. Fertil. Soil 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Paulson, J.N.; Pop, M.; Bravo, H.C. Metastats: An improved statistical method for analysis of metagenomic data. Genome Biol. 2011, 12, P17. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.R.; Shin, J.; Guevarra, R.; Lee, J.H.; Kim, D.W.; Seol, K.H.; Lee, J.H.; Kim, H.B.; Isaacson, R. Deciphering Diversity Indices for a Better Understanding of Microbial Communities. J. Microbiol. Biotechnol. 2017, 27, 2089–2093. [Google Scholar] [CrossRef] [Green Version]
- Xiong, C.; Zhu, Y.-G.; Wang, J.-T.; Singh, B.; Han, L.-L.; Shen, J.-P.; Li, P.-P.; Wang, G.-B.; Wu, C.-F.; Ge, A.-H.; et al. Host selection shapes crop microbiome assembly and network complexity. New Phytol. 2020. [Google Scholar] [CrossRef]
- Wolińska, A.; Górniak, D.; Zielenkiewicz, U.; Goryluk-Salmonowicz, A.; Kuzniar, A.; Stȩpniewska, Z.; Błaszczyk, M. Microbial biodiversity in arable soils is affected by agricultural practices. Int. Agrophys. 2017, 31, 259–271. [Google Scholar] [CrossRef] [Green Version]
- Mhete, M.; Eze, P.N.; Rahube, T.O.; Akinyemi, F.O. Soil properties influence bacterial abundance and diversity under different land-use regimes in semi-arid environments. Sci. Afr. 2020, 7, e00246. [Google Scholar] [CrossRef]
- Frąc, M.; Hannula, S.E.; Bełka, M.; Jędryczka, M. Fungal Biodiversity and Their Role in Soil Health. Front. Microbiol. 2018, 9. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, A.A.; Haq, S.; Bhat, R.A. Actinomycetes benefaction role in soil and plant health. Microb. Pathog. 2017, 111, 458–467. [Google Scholar] [CrossRef] [PubMed]
- Shange, R.S.; Ankumah, R.O.; Ibekwe, A.M.; Zabawa, R.; Dowd, S.E. Distinct soil bacterial communities revealed under a diversely managed agroecosystem. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleńska, E.; Małek, W.; Wójcik, M.; Swiecicka, I.; Thijs, S.; Vangronsveld, J. Beneficial features of plant growth-promoting rhizobacteria for improving plant growth and health in challenging conditions: A methodical review. Sci. Total Environ. 2020, 743. [Google Scholar] [CrossRef]
- Sun, L.; Gao, J.; Huang, T.; Kendall, J.R.A.; Shen, Q.; Zhang, R. Parental material and cultivation determine soil bacterial community structure and fertility. FEMS Microbiol. Ecol. 2015, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Starke, R.; Kermer, R.; Ullmann-Zeunert, L.; Baldwin, I.T.; Seifert, J.; Bastida, F.; von Bergen, M.; Jehmlich, N. Bacteria dominate the short-term assimilation of plant-derived N in soil. Soil Biol. Biochem. 2016, 96, 30–38. [Google Scholar] [CrossRef]
- Lin, Y.; Ye, G.; Kuzyakov, Y.; Liu, D.; Fan, J.; Ding, W. Long-term manure application increases soil organic matter and aggregation, and alters microbial community structure and keystone taxa. Soil Biol. Biochem. 2019, 134, 187–196. [Google Scholar] [CrossRef]
- Cao, Y.; Zhou, B.; Wang, X.; Meng, H.; Zhang, J.; Li, L.; Hong, J. Different fertilization treatments in coal mining-affected soils change bacterial populations and enable soil reclamation. Ann. Microbiol. 2020, 70. [Google Scholar] [CrossRef]
- De la Cruz-Barrón, M.; Cruz-Mendoza, A.; Navarro–Noya, Y.E.; Ruiz-Valdiviezo, V.M.; Ortíz-Gutiérrez, D.; Ramírez-Villanueva, D.A.; Luna-Guido, M.; Thierfelder, C.; Wall, P.C.; Verhulst, N.; et al. The Bacterial Community Structure and Dynamics of Carbon and Nitrogen when Maize (Zea mays L.) and Its Neutral Detergent Fibre Were Added to Soil from Zimbabwe with Contrasting Management Practices. Microb. Ecol. 2017, 73, 135–152. [Google Scholar] [CrossRef]
- Devane, M.; Robson, B.; Lin, S.; Scholes, P.; Wood, D.; Weaver, L.; Webster-Brown, J.; Gilpin, B. Bacterial community shifts in decomposing cowpats and the subsequent impacts on fecal source indicators for water quality monitoring. Ecol. Indic. 2020, 113. [Google Scholar] [CrossRef]
- Mamun, M.A.A.; Sandeman, M.; Rayment, P.; Brook-Carter, P.; Scholes, E.; Kasinadhuni, N.; Piedrafita, D.; Greenhill, A.R. The composition and stability of the faecal microbiota of Merino sheep. J. Appl. Microbiol. 2020, 128, 280–291. [Google Scholar] [CrossRef] [Green Version]
- de Oliveira, T.B.; de Lucas, R.C.; Scarcella, A.S.D.A.; Contato, A.G.; Pasin, T.M.; Martinez, C.A.; Teixeira de Morales Polizeli, M.D.L. Fungal communities differentially respond to warming and drought in tropical grassland soil. Mol. Ecol. 2020, 29, 1550–1559. [Google Scholar] [CrossRef] [PubMed]
- Barnett, K.; Johnson, S.N. Chapter One—Living in the Soil Matrix: Abiotic Factors Affecting Root Herbivores. In Advances in Insect Physiology; Johnson, S.N., Hiltpold, I., Turlings, T.C.J., Eds.; Academic Press: Cambridge, MA, USA, 2013; Volume 45, pp. 1–52. [Google Scholar]
- Ye, G.; Lin, Y.; Luo, J.; Di, H.J.; Lindsey, S.; Liu, D.; Fan, J.; Ding, W. Responses of soil fungal diversity and community composition to long-term fertilization: Field experiment in an acidic Ultisol and literature synthesis. Appl. Soil Ecol. 2020, 145. [Google Scholar] [CrossRef]
- Chen, X.; Henriksen, T.M.; Svensson, K.; Korsaeth, A. Long-term effects of agricultural production systems on structure and function of the soil microbial community. Appl. Soil Ecol. 2020, 147. [Google Scholar] [CrossRef]
- Ishaq, S.L.; Seipel, T.; Yeoman, C.J.; Menalled, F.D. Soil bacterial communities of wheat vary across the growing season and among dryland farming systems. Geoderma 2020, 358. [Google Scholar] [CrossRef]
- Ouyang, Y.; Norton, J.M. Short-term nitrogen fertilization affects microbial community composition and nitrogen mineralization functions in an agricultural soil. Appl. Environ. Microbiol. 2020, 86. [Google Scholar] [CrossRef]
- Schmidt, J.E.; Kent, A.D.; Brisson, V.L.; Gaudin, A.C.M. Agricultural management and plant selection interactively affect rhizosphere microbial community structure and nitrogen cycling. Microbiome 2019, 7. [Google Scholar] [CrossRef] [Green Version]
- Anderson, C.; Beare, M.; Buckley, H.L.; Lear, G. Bacterial and fungal communities respond differently to varying tillage depth in agricultural soils. PeerJ 2017, 5, e3930. [Google Scholar] [CrossRef] [Green Version]
- Sun, R.; Li, W.; Dong, W.; Tian, Y.; Hu, C.; Liu, B. Tillage Changes Vertical Distribution of Soil Bacterial and Fungal Communities. Front. Microbiol. 2018, 9, 699. [Google Scholar] [CrossRef]
- Miller, M.; Dick, R.P. Thermal stability and activities of soil enzymes as influenced by crop rotations. Soil Biol. Biochem. 1995, 27, 1161–1166. [Google Scholar] [CrossRef]
- Kwiatkowski, C.A.; Harasim, E.; Feledyn-Szewczyk, B.; Antonkiewicz, J. Enzymatic Activity of Loess Soil in Organic and Conventional Farming Systems. Agriculture 2020, 10, 135. [Google Scholar] [CrossRef] [Green Version]
- Pretty, J.; Bharucha, Z.P. Sustainable intensification in agricultural systems. Ann. Bot. 2014, 114, 1571–1596. [Google Scholar] [CrossRef] [PubMed]
- Meyer, A.H.; Wooldridge, J.; Dames, J.F. Relationship between soil alteration index three (AI3), soil organic matter and tree performance in a ‘Cripps Pink’/M7 apple orchard. S. Afr. J. Plant Soil 2014, 31, 173–175. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zone | Field | Crop at the Time of Sampling | Rotated with | Management Approach | Tillage | Amendment(s) Fertilizer(s) | Plant Protection Products (PPP) | Irrigation |
|---|---|---|---|---|---|---|---|---|
| A | 1 | Maize & bean intercropping | Kunde (AIV), cabbage | Smallholder | Reduced | Compost, manure, urine | Homemade, compost tea | Rainfed |
| 1.2 | Kale, broccoli | Onions, garlic, broccoli, kale, terere (AIV), carrot | Water harvest, Drip irrigation | |||||
| 2 | Maize and bean intercropping | Maize and bean intercropping | Conventional | Intensive | C. fertilizer | Commercial | Rainfed | |
| 3 | Cabbage, managu (nightshade—AIV) | Broccoli, kale, managu (AIV) | Smallholder | Reduced | C. fertilizer, manure | |||
| 4 | Maize and bean intercropping | |||||||
| B | 5 | Maize and bean intercropping | Dill, nasturtium | Smallholder | Reduced | Compost | Commercial | Sprinkler from well |
| 6 | Terere (amaranth—AIV) | Carrot, terere (AIV) | Compost, ash | |||||
| 7 | Kunde (cowpea—AIV) | Cowpea, carrot, broccoli | Compost | |||||
| 8 | Saget (spider plant—AIV) | Carrot, butternut squash | ||||||
| 9 | Managu (nightshade—AIV) | Previously empty | ||||||
| C | SSN1 | AIV mix | Cowpea, pigeon pea, chickpea, saget, sorghum | Smallholder | No till | Compost, manure | n/a | Sprinkler from well |
| SSN2 | Maize and bean intercropping | Cauliflower, beans | Conventional | Reduced | C. fertilizer | Commercial | ||
| SSN3 | Terere, managu, pogo (AIV mix) | AIV mix, coriander, onions, cabbage, beans, carrot, melon, pumpkin | C. fertilizer, compost | |||||
| 10 | Managu (nightshade) | Previously empty | Intensive | C. fertilizer, manure | Sprinkler from Melewa River | |||
| 12 | Saget (spider plant) | Previously empty |
| Bacteria | Fungi | ||||||
|---|---|---|---|---|---|---|---|
| Zone | Field | SOBS | Simpson | Chao | SOBS | Simpson | Chao |
| A | 1 | 3482 (±68) a | 492 (±22) a | 10439 (±483) a | 1835 (±48) b | 33 (±5) b | 3507 (±105) a |
| 1.2 | 3288 (±95) b | 274 (±101) b | 9607 (±336) a | 2046 (±12) a | 32 (±6) b | 3735 (±302) a | |
| 2 | 3513 (±59) a | 428 (±16) ab | 10390 (±413) a | 2028 (±60) a | 59 (±17) a | 3878 (±110) a | |
| 3 | 3284 (±59) b | 403 (±21) ab | 9640 (±304) a | 1849 (±33) ab | 48 (±1) ab | 3727 (±129) a | |
| 4 | 3321 (±44) ab | 425 (±24) ab | 9534 (±388) a | 1825 (±48) b | 55 (±5) ab | 3414 (±101) a | |
| B | 5 | 3114 (±64) a | 410 (±20) a | 8829 (±316) ab | 1485 (±65) b | 31 (±3) b | 2692 (±88) b |
| 6 | 3173 (±70) a | 349 (±26) a | 9438 (±195) a | 1144 (±114) bc | 17 (±6) b | 2078 (±212) c | |
| 7 | 3202 (±50) a | 365 (±16) a | 9416 (±161) a | 1443 (±44) c | 22 (±2) b | 2834 (±89) b | |
| 8 | 3083 (±68) a | 315 (±58) a | 8735 (±104) b | 1930 (±80) a | 54 (±10) a | 4319 (±244) a | |
| 9 | 3051 (±37) a | 366 (±25) a | 8208 (±218) b | 1620 (±155) b | 27 (±9) b | 3264 (±262) b | |
| C | SSN1 | 2895 (±51) B | 299 (±15) C | 8867 (±233) CD | 1375 (±167) B | 16 (±7) B | 2728 (±190) C |
| SSN2 | 2842 (±39) B | 302 (±22) C | 8667 (±251) D | 1680 (±108) AB | 33 (±5) AB | 3098 (±183) BC | |
| SSN3 | 3180 (±70) A | 325 (±26) BC | 9933 (±414) AB | 1522 (±84) AB | 33 (±3) AB | 2922 (±109) BC | |
| 10 | 3274 (±45) A | 415 (±19) A | 10597 (±368) A | 1822 (±36) A | 29 (±3) AB | 3415 (±141) AB | |
| 12 | 3244 (±35) A | 386 (±31) AB | 9652 (±156) BC | 1780 (±112) A | 38 (±9) A | 3728 (±235) A | |
| Practices | Crop | + | + | n.s. | + | + | + |
| Tillage | + | + | + | + | + | + | |
| Fertilization | + | n.s. | + | + | + | + | |
| PPP | + | + | n.s. | + | + | + | |
| Soil Alteration * | Measured Soil Enzymes | ||||
|---|---|---|---|---|---|
| Zone | Field | AI3 Score | β GLU (μmol PNG g−1 h−1) | PHO (μmol PNP g−1 h−1) | URE (μg Urea g−1 h−1) |
| A | 1 | 31.62 (±4.2) b | 9.99 (±0.6) a | 5.2 (±0.1) a | 8.8 (±0.4) a |
| 1.2 | 41.94 (±2.6) a | 10.36 (±0.5) a | 4.44 (±0.2) b | 6.35 (±0.2) b | |
| 2 | 17.19 (±4.1) c | 5.94 (±0.8) b | 3.22 (±0.4) c | 6.28 (±0.4) b | |
| 3 | 37.82 (±3.7) ab | 10.04 (±0.5) a | 4.51 (±0.1) ab | 8.4 (±0.1) a | |
| 4 | 6.08 (±1.4) d | 3.83 (±0.2) c | 2.64 (±0.1) c | 4.71 (±0.1) c | |
| B | 5 | 16.96 (±1.8) c | 4.34 (±0.16) c | 1.09 (±0.06) d | 1.76 (±0.13) e |
| 6 | 27.34 (±4.2) b | 6.09 (±0.72) c | 0.5 (±0.06) e | 2.45 (±0.08) d | |
| 7 | 25.44 (±2.1) b | 14.19 (±0.88) a | 5.12 (±0.14) a | 8.07 (±0.12) b | |
| 8 | 24.33 (±1.5) b | 12.43 (±0.98) a | 3.74 (±0.1) c | 5.18 (±0.12) c | |
| 9 | 42.61 (±5.6) a | 9 (±0.54) b | 4.23 (±0.06) b | 10.74 (±0.21) a | |
| C | SSN1 | 65.56 (±6.9) B | 3.23 (±0.15) C | 0.73 (±0.16) C | 5.02 (±0.24) B |
| SSN2 | 64.59 (±8.2) B | 4.43 (±0.47) C | 0.72 (±0.06) C | 3.38 (±0.22) C | |
| SSN3 | 30.76 (±4.2) C | 4.63 (±0.21) C | 1.18 (±0.1) B | 2.63 (±0.3) D | |
| 10 | 85.1 (±6.2) A | 13.21 (±0.74) A | 1.94 (±0.1) A | 5.95 (±0.1) A | |
| 12 | 63.47 (±5) B | 9.57 (±0.58) B | 1.1 (±0.1) B | 5.62 (±0.17) AB | |
| Biodiversity | Community | Enzymes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Bacteria | Fungi | Bacteria | Fungi | AI3 Score Impact | ||||||
| Practice | SOBS | Chao | Simpson | SOBS | Chao | Simpson | ||||
| Significance | Crop | + | + | n.s. | + | + | + | + | + | + |
| Tillage | + | + | + | + | + | + | + | + | + | |
| Fertilization | + | n.s. | + | + | + | + | + | + | + | |
| Plant protection | + | + | n.s. | + | + | + | + | + | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Taskin, E.; Misci, C.; Bandini, F.; Fiorini, A.; Pacini, N.; Obiero, C.; Sila, D.N.; Tabaglio, V.; Puglisi, E. Smallholder Farmers’ Practices and African Indigenous Vegetables Affect Soil Microbial Biodiversity and Enzyme Activities in Lake Naivasha Basin, Kenya. Biology 2021, 10, 44. https://doi.org/10.3390/biology10010044
Taskin E, Misci C, Bandini F, Fiorini A, Pacini N, Obiero C, Sila DN, Tabaglio V, Puglisi E. Smallholder Farmers’ Practices and African Indigenous Vegetables Affect Soil Microbial Biodiversity and Enzyme Activities in Lake Naivasha Basin, Kenya. Biology. 2021; 10(1):44. https://doi.org/10.3390/biology10010044
Chicago/Turabian StyleTaskin, Eren, Chiara Misci, Francesca Bandini, Andrea Fiorini, Nic Pacini, Clifford Obiero, Daniel Ndaka Sila, Vincenzo Tabaglio, and Edoardo Puglisi. 2021. "Smallholder Farmers’ Practices and African Indigenous Vegetables Affect Soil Microbial Biodiversity and Enzyme Activities in Lake Naivasha Basin, Kenya" Biology 10, no. 1: 44. https://doi.org/10.3390/biology10010044
APA StyleTaskin, E., Misci, C., Bandini, F., Fiorini, A., Pacini, N., Obiero, C., Sila, D. N., Tabaglio, V., & Puglisi, E. (2021). Smallholder Farmers’ Practices and African Indigenous Vegetables Affect Soil Microbial Biodiversity and Enzyme Activities in Lake Naivasha Basin, Kenya. Biology, 10(1), 44. https://doi.org/10.3390/biology10010044