
Extensive GJD2 Expression in the Song Motor Pathway Reveals the Extent of Electrical Synapses in the Songbird Brain

 , , ,
, , , 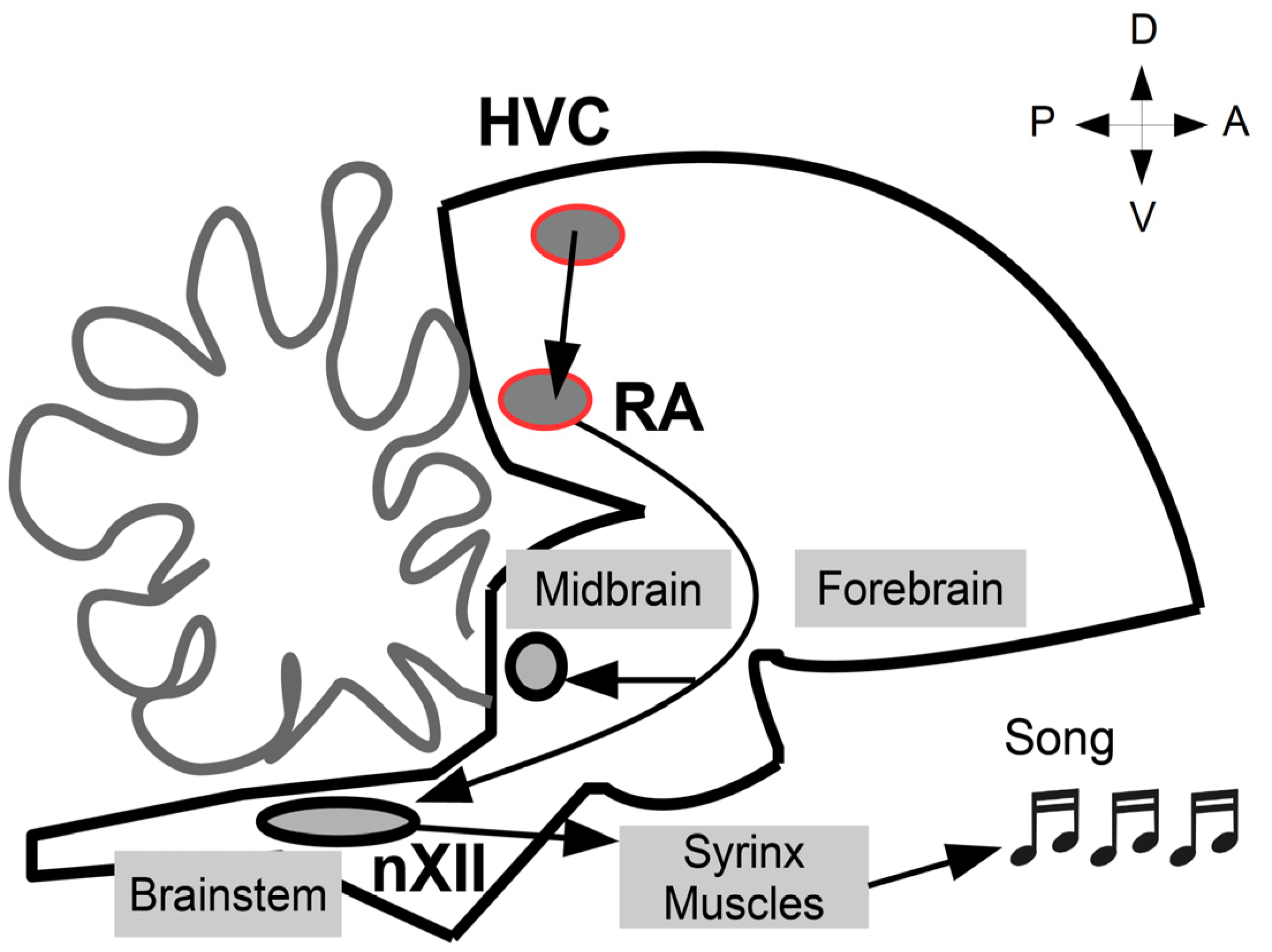{kind=link}
{kind=link}
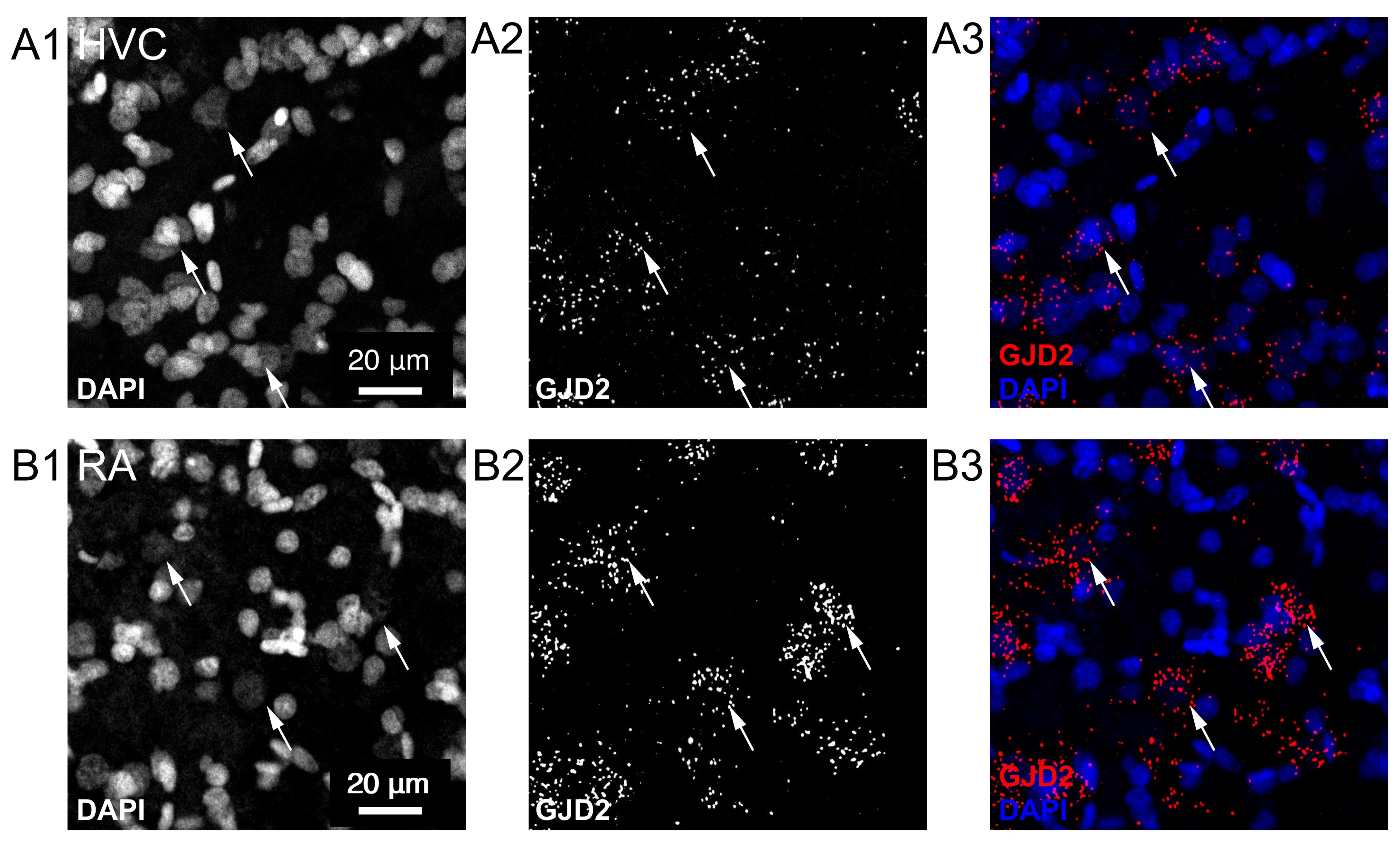{kind=link}
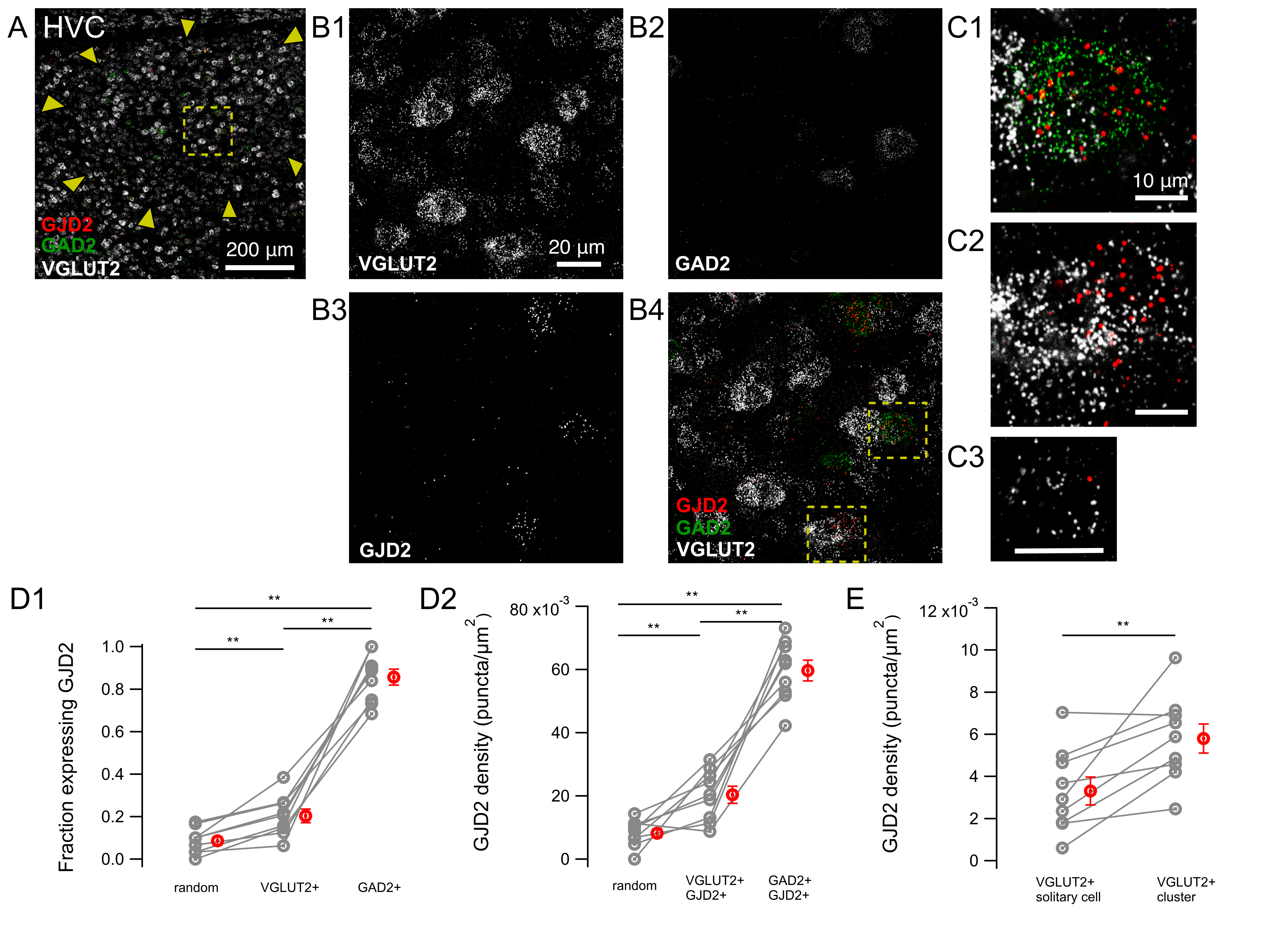{kind=link}
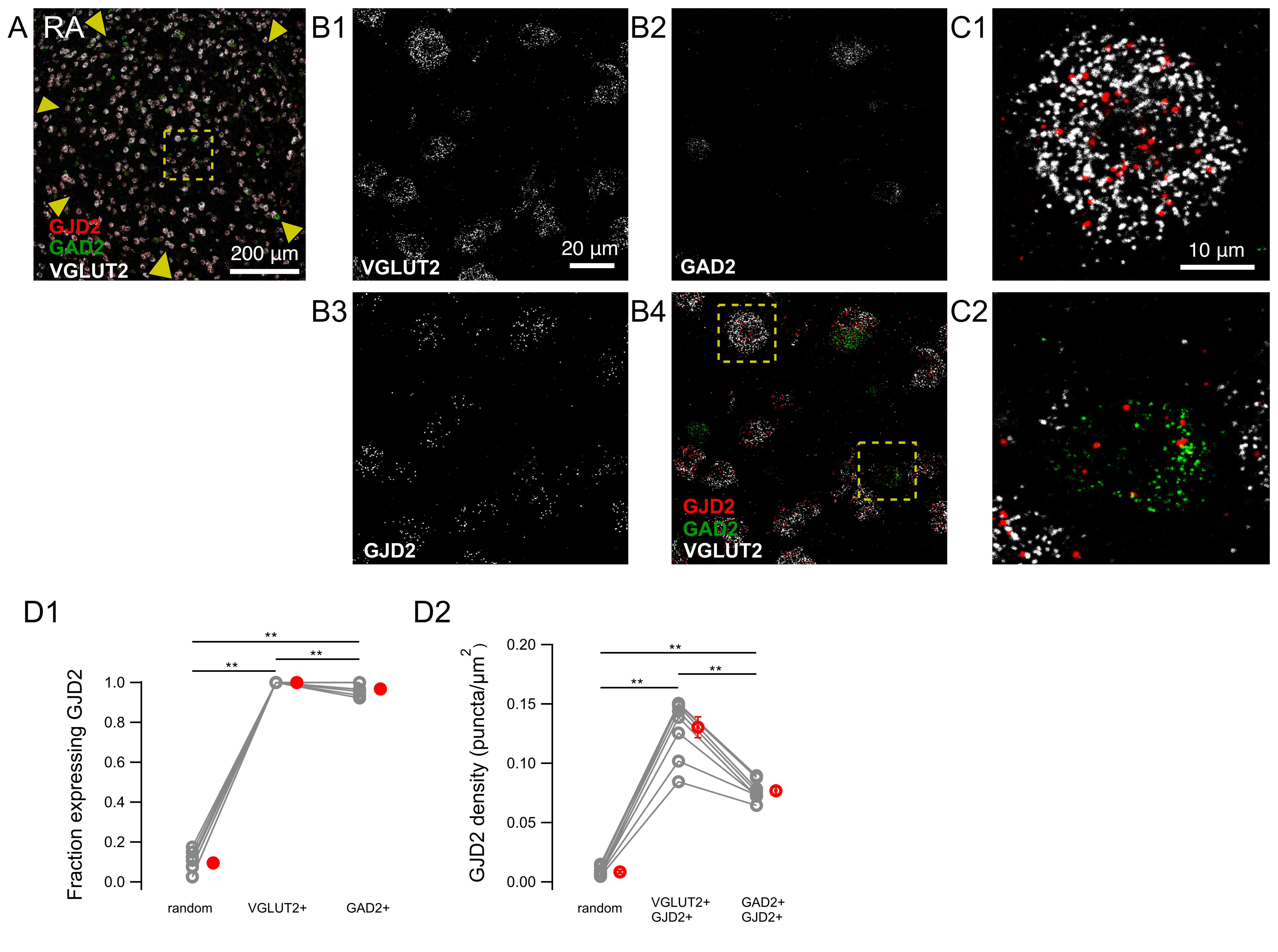{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Fluorescence in Situ Hybridizations
2.3. Confocal Microscopy
2.4. Analysis
2.5. Cellular Transcriptomics Dataset
2.6. Statistics
3. Results
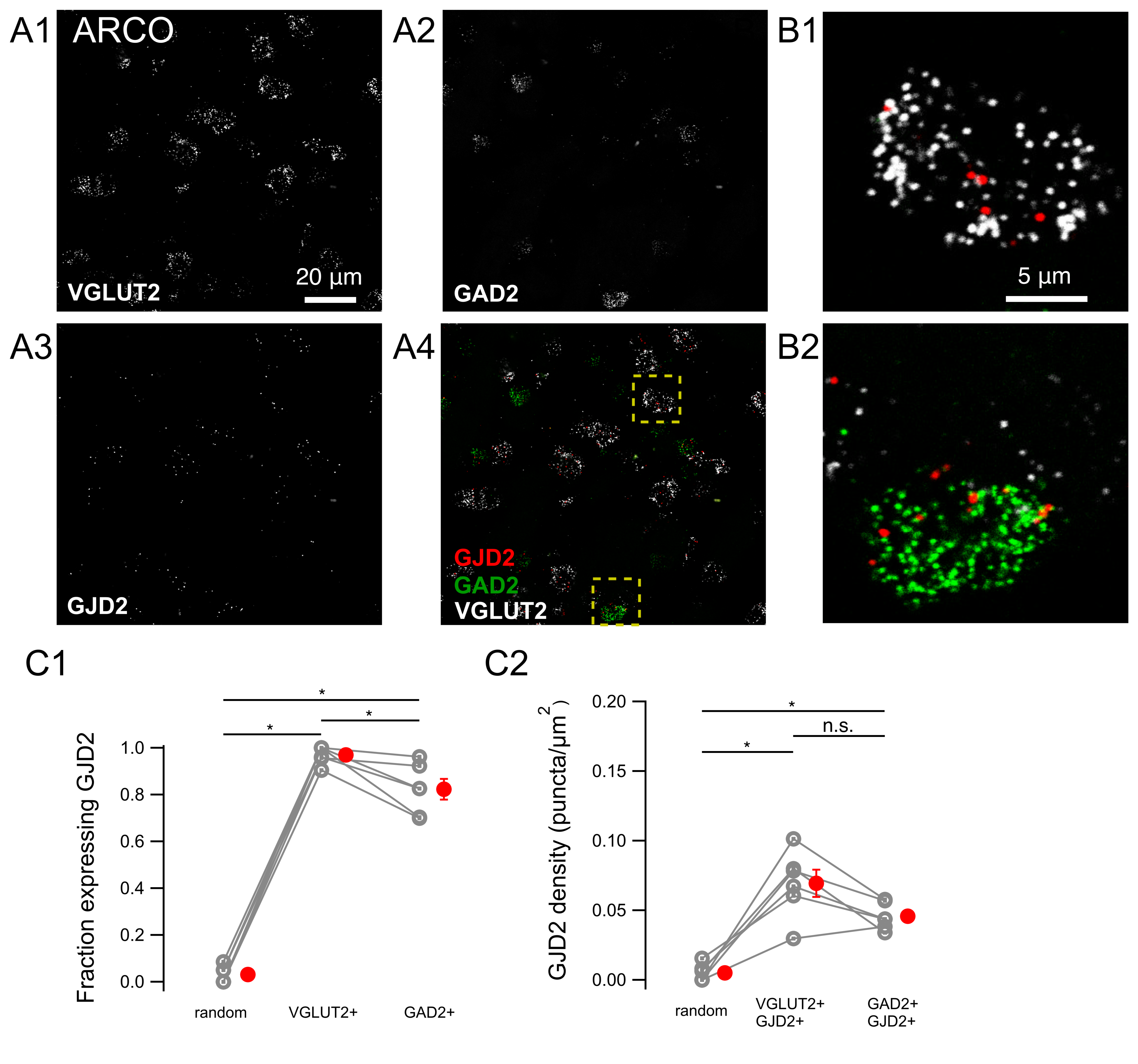
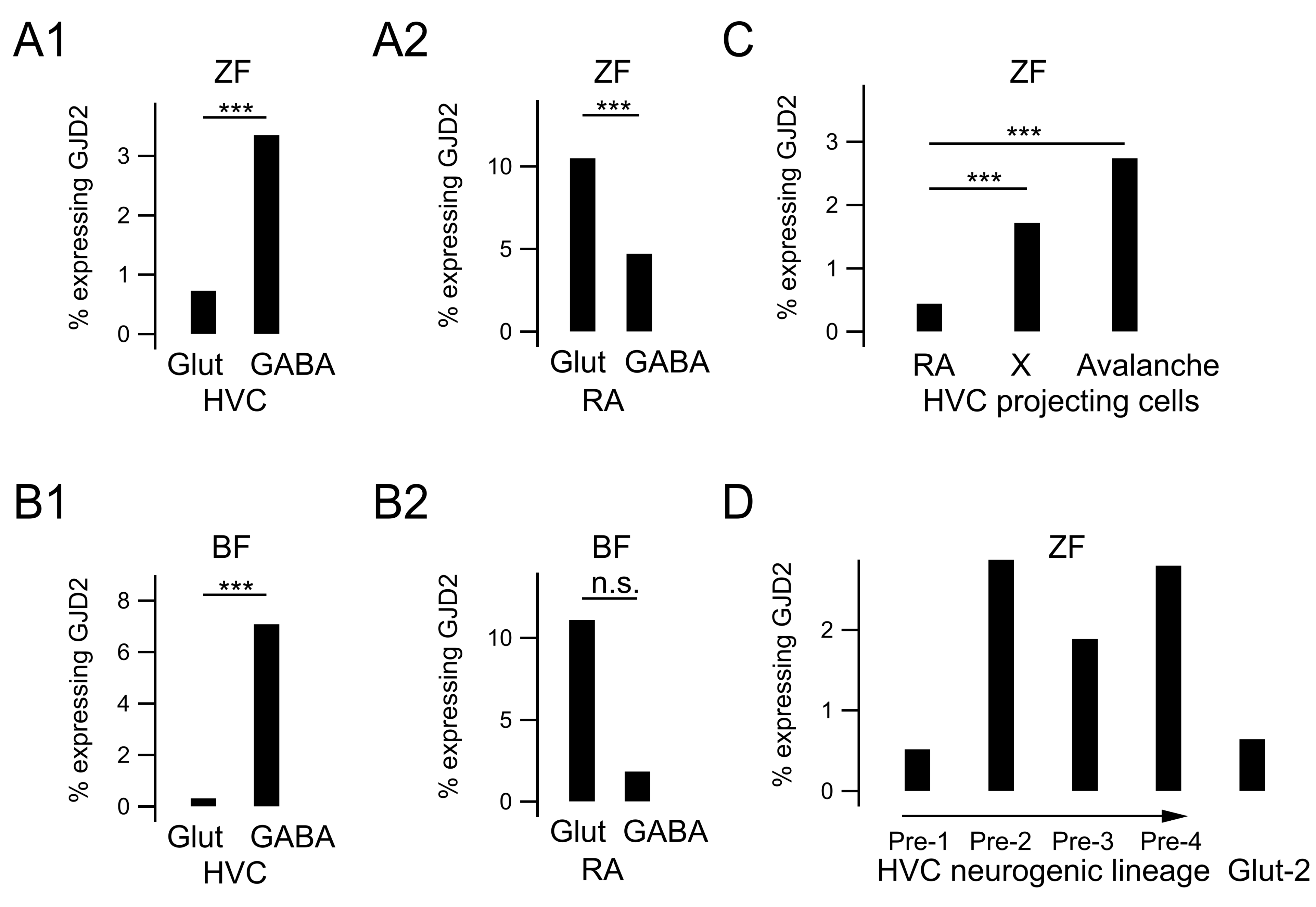
3.1. GJD2 mRNA Is Sparsely Expressed in HVC and Ubiquitously in RA
3.2. GJD2 mRNA Expression in GABAergic HVC Neurons and A Subset of Glutamatergic HVC Neurons
3.3. GJD2 mRNA Expression in RA GABAergic and Glutamatergic Neurons
3.4. GJD2 mRNA Expression in the Nidopallium
3.5. GJD2 mRNA Expression in the Arcopallium
3.6. GJD2 Expression in Single-Cell RNA Sequencing Data
4. Discussion
4.1. GJD2 in the Song Control Circuits
4.2. Comparison of in Situ Hybridizations and Single-Cell Transcriptomics
4.3. Electrical Synaptic Phenotype of Song Nuclei: Evolutionary Considerations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RA | the robust nucleus of the arcopallium |
| HVC | previously used as an abbreviation of ‘High Vocal Center’; used here as a proper name |
| GJD2 | gap junction protein delta 2 |
| ROI | region of interest |
| VGLUT2 | vesicular glutamate transporter 2 |
| GAD2 | glutamate decarboxylase 2 |
References
- Rela, L.; Szczupak, L. Gap junctions: Their importance for the dynamics of neural circuits. Mol. Neurobiol. 2004, 30, 341–357. [Google Scholar] [CrossRef]
- Connors, B.W. Synchrony and so much more: Diverse roles for electrical synapses in neural circuits. Dev. Neurobiol. 2017, 77, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Alcamí, P.; Pereda, A.E. Beyond plasticity: The dynamic impact of electrical synapses on neural circuits. Nat. Rev. Neurosci. 2019, 20, 253–271. [Google Scholar] [CrossRef] [PubMed]
- Tamás, G.; Buhl, E.H.; Lörincz, A.; Somogyi, P. Proximally targeted GABAergic synapses and gap junctions synchronize cortical interneurons. Nat. Neurosci. 2000, 3, 366–371. [Google Scholar] [CrossRef] [PubMed]
- Galarreta, M.; Hestrin, S. Electrical synapses between GABA-releasing interneurons. Nat. Rev. Neurosci. 2001, 2, 425–433. [Google Scholar] [CrossRef]
- Vervaeke, K.; Lőrincz, A.; Gleeson, P.; Farinella, M.; Nusser, Z.; Silver, R.A. Rapid Desynchronization of an Electrically Coupled Interneuron Network with Sparse Excitatory Synaptic Input. Neuron 2010, 67, 435–451. [Google Scholar] [CrossRef] [Green Version]
- Alcami, P. Electrical Synapses Enhance and Accelerate Interneuron Recruitment in Response to Coincident and Sequential Excitation. Front. Cell. Neurosci. 2018, 12, 156. [Google Scholar] [CrossRef] [Green Version]
- Alcami, P.; Marty, A. Estimating functional connectivity in an electrically coupled interneuron network. Proc. Nat. Acad. Sci. 2013, 110, E4798–E4807. [Google Scholar] [CrossRef] [Green Version]
- Bennett, M.V.L.; Zukin, R.S. Electrical coupling and neuronal synchronization in the Mammalian brain. Neuron 2004, 41, 495–511. [Google Scholar] [CrossRef] [Green Version]
- Pereda, A.E. Electrical synapses and their functional interactions with chemical synapses. Nat. Rev. Neurosci. 2014, 15, 250–263. [Google Scholar] [CrossRef]
- Nagy, J.I.; Pereda, A.E.; Rash, J.E. Electrical synapses in mammalian CNS: Past eras, present focus and future directions. Biochim. Biophys. Acta (BBA) Biomembr. 2018, 1860, 102–123. [Google Scholar] [CrossRef] [PubMed]
- Berthoud, V.M.; Singh, R.; Minogue, P.J.; Ragsdale, C.W.; Beyer, E.C. Highly restricted pattern of connexin36 expression in chick somite development. Anat. Embryol. 2004, 209, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Frankl-Vilches, C.; Kuhl, H.; Werber, M.; Klages, S.; Kerick, M.; Bakker, A.; de Oliveira, E.H.C.; Reusch, C.; Capuano, F.; Vowinckel, J.; et al. Using the canary genome to decipher the evolution of hormone-sensitive gene regulation in seasonal singing birds. Genome Biol. 2015, 16, 19. [Google Scholar] [CrossRef] [Green Version]
- Kihara, A.H.; Paschon, V.; Cardoso, C.M.; Higa, G.S.V.; Castro, L.M.; Hamassaki, D.E.; Britto, L.R.G. Connexin36, an essential element in the rod pathway, is highly expressed in the essentially rodless retina of Gallus gallus. J. Comp. Neurol. 2009, 512, 651–663. [Google Scholar] [CrossRef]
- Paschon, V.; Higa, G.S.V.; Resende, R.R.; Britto, L.R.G.; Kihara, A.H. Blocking of Connexin-Mediated Communication Promotes Neuroprotection during Acute Degeneration Induced by Mechanical Trauma. PLoS ONE 2012, 7, e45449. [Google Scholar] [CrossRef]
- Hahnloser, R.H.R.; Kozhevnikov, A.A.; Fee, M.S. An ultra-sparse code underlies the generation of neural sequences in a songbird. Nature 2002, 419, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Cohen, Y.; Shen, J.; Semu, D.; Leman, D.P.; Liberti, W.A.; Perkins, L.N.; Liberti, D.C.; Kotton, D.N.; Gardner, T.J. Hidden neural states underlie canary song syntax. Nature 2020, 582, 539–544. [Google Scholar] [CrossRef] [PubMed]
- Sober, S.J.; Wohlgemuth, M.J.; Brainard, M.S. Central Contributions to Acoustic Variation in Birdsong. J. Neurosci. 2008, 28, 10370–10379. [Google Scholar] [CrossRef]
- Lynch, G.F.; Okubo, T.S.; Hanuschkin, A.; Hahnloser, R.H.R.; Fee, M.S. Rhythmic Continuous-Time Coding in the Songbird Analog of Vocal Motor Cortex. Neuron 2016, 90, 877–892. [Google Scholar] [CrossRef] [Green Version]
- Long, M.A.; Jin, D.Z.; Fee, M.S. Support for a synaptic chain model of neuronal sequence generation. Nature 2010, 468, 394–399. [Google Scholar] [CrossRef] [Green Version]
- Miller, M.N.; Cheung, C.Y.J.; Brainard, M.S. Vocal learning promotes patterned inhibitory connectivity. Nat. Commun. 2017, 8, 2105. [Google Scholar] [CrossRef] [Green Version]
- Kosche, G.; Vallentin, D.; Long, M.A. Interplay of Inhibition and Excitation Shapes a Premotor Neural Sequence. J. Neurosci. 2015, 35, 1217–1227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamaguchi, K.; Tanaka, M.; Mooney, R. A Distributed Recurrent Network Contributes to Temporally Precise Vocalizations. Neuron 2016, 91, 680–693. [Google Scholar] [CrossRef] [Green Version]
- Burd, G.D.; Nottebohm, F. Ultrastructural characterization of synaptic terminals formed on newly generated neurons in a song control nucleus of the adult canary forebrain. J. Comp. Neurol. 1985, 240, 143–152. [Google Scholar] [CrossRef]
- Chakraborty, M.; Jarvis, E.D. Brain evolution by brain pathway duplication. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20150056. [Google Scholar] [CrossRef] [Green Version]
- Colquitt, B.M.; Merullo, D.P.; Konopka, G.; Roberts, T.F.; Brainard, M.S. Cellular transcriptomics reveals evolutionary identities of songbird vocal circuits. Science 2021, 371, eabd9704. [Google Scholar] [CrossRef]
- Mikalsen, S.-O.; í Kongsstovu, S.; Tausen, M. Connexins during 500 Million Years—From Cyclostomes to Mammals. Int. J. Mol. Sci. 2021, 22, 1584. [Google Scholar] [CrossRef] [PubMed]
- Fiete, I.R.; Seung, H.S. Neural network models of birdsong production, learning, and coding. In New Encyclopedia of Neuroscience; Squire, L., Albright, T., Bloom, F., Gage, F., Spitzer, N., Eds.; Elsevier: New York, NY, USA, 2007. [Google Scholar]
- Gahr, M.; Garcia-Segura, L.M. Testosterone-dependent increase of gap-junctions in HVC neurons of adult female canaries. Brain Res. 1996, 712, 69–73. [Google Scholar] [CrossRef]
- Dutar, P.; Vu, H.M.; Perkel, D.J. Multiple Cell Types Distinguished by Physiological, Pharmacological, and Anatomic Properties in Nucleus HVc of the Adult Zebra Finch. J. Neurophysiol. 1998, 80, 1828–1838. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, A.; Kirn, J.R.; Nottebohm, F. Birth of projection neurons in adult avian brain may be related to perceptual or motor learning. Science 1990, 249, 1444–1446. [Google Scholar] [CrossRef] [PubMed]
- Rörig, B.; Klausa, G.; Sutor, B. Intracellular acidification reduced gap junction coupling between immature rat neocortical pyramidal neurones. J. Physiol. 1996, 490, 31–49. [Google Scholar] [CrossRef] [Green Version]
- Peinado, A.; Yuste, R.; Katz, L.C. Extensive dye coupling between rat neocortical neurons during the period of circuit formation. Neuron 1993, 10, 103–114. [Google Scholar] [CrossRef]
- Haque, A.; Engel, J.; Teichmann, S.A.; Lönnberg, T. A practical guide to single-cell RNA-sequencing for biomedical research and clinical applications. Genome Med. 2017, 9, 75. [Google Scholar] [CrossRef]
- Kozareva, V.; Martin, C.; Osorno, T.; Rudolph, S.; Guo, C.; Vanderburg, C.; Nadaf, N.; Regev, A.; Regehr, W.G.; Macosko, E. A transcriptomic atlas of mouse cerebellar cortex comprehensively defines cell types. Nature 2021, 598, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Lovell, P.V.; Wirthlin, M.; Kaser, T.; Buckner, A.A.; Carleton, J.B.; Snider, B.R.; McHugh, A.K.; Tolpygo, A.; Mitra, P.P.; Mello, C.V. ZEBrA: Zebra finch Expression Brain Atlas—A resource for comparative molecular neuroanatomy and brain evolution studies. J. Comp. Neurol. 2020, 528, 2099–2131. [Google Scholar] [CrossRef] [PubMed]
- Nevue, A.A.; Lovell, P.V.; Wirthlin, M.; Mello, C.V. Molecular specializations of deep cortical layer analogs in songbirds. Sci. Rep. 2020, 10, 18767. [Google Scholar] [CrossRef]
- Zemel, B.M.; Nevue, A.A.; Dagostin, A.; Lovell, P.V.; Mello, C.V.; von Gersdorff, H. Resurgent Na+ currents promote ultrafast spiking in projection neurons that drive fine motor control. BioRxiv 2021, 459677. [Google Scholar] [CrossRef]
- Kuhl, H.; Frankl-Vilches, C.; Bakker, A.; Mayr, G.; Nikolaus, G.; Boerno, S.T.; Klages, S.; Timmermann, B.; Gahr, M. An Unbiased Molecular Approach Using 3′-UTRs Resolves the Avian Family-Level Tree of Life. Mol. Biol. Evol. 2021, 38, 108–127. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alcami, P.; Totagera, S.; Sohnius-Wilhelmi, N.; Leitner, S.; Grothe, B.; Frankl-Vilches, C.; Gahr, M. Extensive GJD2 Expression in the Song Motor Pathway Reveals the Extent of Electrical Synapses in the Songbird Brain. Biology 2021, 10, 1099. https://doi.org/10.3390/biology10111099
Alcami P, Totagera S, Sohnius-Wilhelmi N, Leitner S, Grothe B, Frankl-Vilches C, Gahr M. Extensive GJD2 Expression in the Song Motor Pathway Reveals the Extent of Electrical Synapses in the Songbird Brain. Biology. 2021; 10(11):1099. https://doi.org/10.3390/biology10111099
Chicago/Turabian StyleAlcami, Pepe, Santhosh Totagera, Nina Sohnius-Wilhelmi, Stefan Leitner, Benedikt Grothe, Carolina Frankl-Vilches, and Manfred Gahr. 2021. "Extensive GJD2 Expression in the Song Motor Pathway Reveals the Extent of Electrical Synapses in the Songbird Brain" Biology 10, no. 11: 1099. https://doi.org/10.3390/biology10111099
APA StyleAlcami, P., Totagera, S., Sohnius-Wilhelmi, N., Leitner, S., Grothe, B., Frankl-Vilches, C., & Gahr, M. (2021). Extensive GJD2 Expression in the Song Motor Pathway Reveals the Extent of Electrical Synapses in the Songbird Brain. Biology, 10(11), 1099. https://doi.org/10.3390/biology10111099