
The Oleaginous Yeast Metschnikowia pulcherrima Displays Killer Activity against Avian-Derived Pathogenic Bacteria




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Strains, Strain Maintenance and Media
2.3. Zone of Inhibition and Liquid Competition Assays
2.3.1. Zone of Inhibition Assays
2.3.2. Liquid Competition Assays
2.4. Supernatant Generation for Bacterial Growth Assay and SDS-PAGE
2.5. Statistical Analyses
3. Results
3.1. ZOI Assays
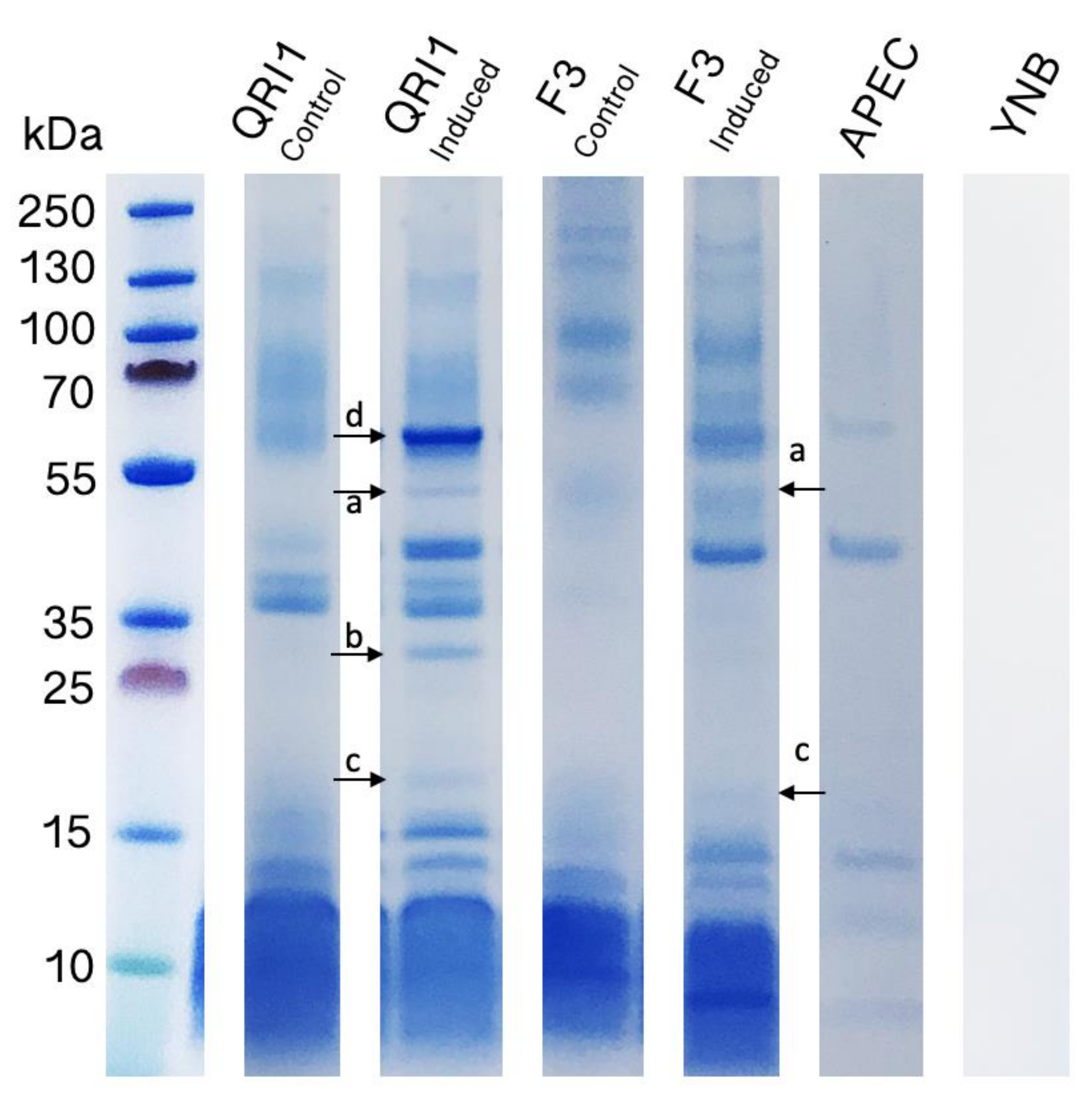
3.2. Antimicrobial Effects and Competition in Liquid
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chattopadhyay, M.K. Use of Antibiotics as Feed Additives: A Burning Question. Front. Microbiol. 2014, 5, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darwish, W.S.; Eldaly, E.A.; El-Abbasy, M.T.; Ikenaka, Y.; Nakayama, S.; Ishizuka, M. Antibiotic Residues in Food: The African Scenario. Jpn. J. Vet. Res. 2013, 61, S13–S22. [Google Scholar] [PubMed]
- Chaucheyras-Durand, F.; Durand, H. Probiotics in Animal Nutrition and Health. Benef. Microbes 2010, 1, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Haque, M.; Islam, K.; Khaleduzzaman, A. A Review On Antibiotics In An Animal Feed. Bangladesh J. Anim. Sci. 2009, 38, 22–32. [Google Scholar] [CrossRef] [Green Version]
- Chowdhary, A.; Meis, J.F. Emergence of Azole Resistant Aspergillus Fumigatus and One Health: Time to Implement Environmental Stewardship. Environ. Microbiol. 2018, 20, 1299–1301. [Google Scholar] [CrossRef] [Green Version]
- Fisher, M.C.; Hawkins, N.J.; Sanglard, D.; Gurr, S.J. Worldwide Emergence of Resistance to Antifungal Drugs Challenges Human Health and Food Security. Science 2018, 360, 739–742. [Google Scholar] [CrossRef] [Green Version]
- Woolhouse, M.; Ward, M.; Van Bunnik, B.; Farrar, J. Antimicrobial Resistance in Humans, Livestock and the Wider Environment. Philos. Trans. R. Soc. B Biol. Sci. 2015, 370, 20140083. [Google Scholar] [CrossRef]
- Oie List of Antimicrobials of Veterinary Importance; World Organization for Animal Health: Paris France, 2007.
- Marshall, B.M.; Levy, S.B. Food Animals and Antimicrobials: Impacts on Human Health. Clin. Microbiol. Rev. 2011, 24, 718–733. [Google Scholar] [CrossRef] [Green Version]
- Kirchhelle, C. Pharming Animals: A Global History of Antibiotics in Food Production (1935–2017). Palgrave Commun. 2018, 4, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Hoelzer, K.; Bielke, L.; Blake, D.P.; Cox, E.; Cutting, S.M.; Devriendt, B.; Erlacher-Vindel, E.; Goossens, E.; Karaca, K.; Lemiere, S.; et al. Vaccines as Alternatives to Antibiotics for Food Producing Animals. Part 2: New Approaches and Potential Solutions. Vet. Res. 2018, 49, 13567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Sangthong, R.; McNeil, E.; Tang, R.; Chongsuvivatwong, V. Antibiotic Use in Chicken Farms in Northwestern China. Antimicrob. Resist. Infect. Control 2020, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. Probiotics in Animal Nutrition-Production, Impact and Regulation; FAO: Rome, Italy, 2016; ISBN 2-9524354-0-5. [Google Scholar]
- Markowiak, P.; Ślizewska, K. The Role of Probiotics, Prebiotics and Synbiotics in Animal Nutrition. Gut Pathog. 2018, 10, 21. [Google Scholar] [CrossRef] [PubMed]
- Micciche, A.C.; Foley, S.L.; Pavlidis, H.O.; McIntyre, D.R.; Ricke, S.C. A Review of Prebiotics against Salmonella in Poultry: Current and Future Potential for Microbiome Research Applications. Front. Vet. Sci. 2018, 5, 191. [Google Scholar] [CrossRef] [Green Version]
- Vohra, A.; Syal, P.; Madan, A. Probiotic Yeasts in Livestock Sector. Anim. Feed. Sci. Technol. 2016, 219, 31–47. [Google Scholar] [CrossRef]
- Line, J.E.; Bailey, J.S.; Cox, N.A.; Stern, N.J.; Tompkins, T. Effect of Yeast-Supplemented Feed on Salmonella and Campylobacter Populations in Broilers. Poult. Sci. 1998, 77, 405–410. [Google Scholar] [CrossRef]
- Hassanein, S.M.; Soliman, N.K. Effect of Probiotic (Saccharomyces Cerevisae) Adding to Diets on Intestinal Microflora and Performance of Hy-Line Layer Hens. J. Am. Sci. 2010, 6, 159–169. [Google Scholar]
- Abeln, F.; Chuck, C.J. Achieving a High-Density Oleaginous Yeast Culture: Comparison of Four Processing Strategies Using Metschnikowia Pulcherrima. Biotechnol. Bioeng. 2019, 12, 3200–3214. [Google Scholar] [CrossRef] [PubMed]
- Hicks, R.H.; Chuck, C.J.; Scott, R.J.; Leak, D.J.; Henk, D.A. Comparison of Nile Red and Cell Size Analysis for High-Throughput Lipid Estimation Within Oleaginous Yeast. Eur. J. Lipid Sci. Technol. 2019, 121, 1800355. [Google Scholar] [CrossRef] [Green Version]
- Gore-Lloyd, D.; Sumann, I.; Brachmann, A.O.; Schneeberger, K.; Ortiz-Merino, R.A.; Moreno-Beltrán, M.; Schläfli, M.; Kirner, P.; Santos Kron, A.; Rueda-Mejia, M.P.; et al. Snf2 Controls Pulcherriminic Acid Biosynthesis and Antifungal Activity of the Biocontrol Yeast Metschnikowia Pulcherrima. Mol. Microbiol. 2019, 124, 1058–1067. [Google Scholar] [CrossRef] [Green Version]
- Sipiczki, M. Metschnikowia Strains Isolated from Botrytized Grapes Antagonize Fungal and Bacterial Growth by Iron Depletion. Appl. Environ. Microbiol. 2006, 72, 6716–6724. [Google Scholar] [CrossRef] [Green Version]
- Piano, S.; Neyrotti, V.; Migheli, Q.; Gullino, M.L. Biocontrol Capability of Metschnikowia Pulcherrima against Botrytis Postharvest Rot of Apple. Postharvest Biol. Technol. 1997, 3, 131–140. [Google Scholar] [CrossRef]
- Oro, L.; Ciani, M.; Comitini, F. Antimicrobial Activity of Metschnikowia Pulcherrima on Wine Yeasts. J. Appl. Microbiol. 2014, 116, 1209–1217. [Google Scholar] [CrossRef]
- Sipiczki, M. Metschnikowia Pulcherrima and Related Pulcherrimin-Producing Yeasts: Fuzzy Species Boundaries and Complex Antimicrobial Antagonism. Microorganisms 2020, 8, 1029. [Google Scholar] [CrossRef]
- Abeln, F.; Fan, J.; Budarin, V.L.; Briers, H.; Parsons, S.; Allen, M.J.; Henk, D.A.; Clark, J.; Chuck, C.J. Lipid Production through the Single-Step Microwave Hydrolysis of Macroalgae Using the Oleaginous Yeast Metschnikowia Pulcherrima. Algal Research 2019, 38, 101411. [Google Scholar] [CrossRef] [Green Version]
- Moreno-Beltrán, M.; Gore-Lloyd, D.; Chuck, C.; Henk, D. Variation among Metschnikowia Pulcherrima Isolates for Genetic Modification and Homologous Recombination. Microorganisms 2021, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Hicks, R.H.; Sze, Y.; Chuck, C.J.; Henk, D.A. Enhanced Inhibitor Tolerance and Increased Lipid Productivity through Adaptive Laboratory Evolution in the Oleaginous Yeast Metshnikowia Pulcherrima. bioRxiv 2020, 2020.02.17.952291. [Google Scholar] [CrossRef] [Green Version]
- Moreno Beltran, M. Using Diverse Methods to Expand the Sustainable Industrial Promise of an All-Rounder Yeast, Metschnikowia Pulcherrima. Ph.D. Thesis, University of Bath, Bath, UK, 2021. [Google Scholar]
- Kogut, M.H. The Effect of Microbiome Modulation on the Intestinal Health of Poultry. Anim. Feed. Sci. Technol. 2019, 250, 32–40. [Google Scholar] [CrossRef]
- Türkel, S.; Korukluoğlu, M.; Yavuz, M. Biocontrol Activity of the Local Strain of Metschnikowia Pulcherrima on Different Postharvest Pathogens. Biotechnol. Res. Int. 2014, 2014, 397167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanakumar, D.; Ciavorella, A.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Metschnikowia Pulcherrima Strain MACH1 Outcompetes Botrytis Cinerea, Alternaria Alternata and Penicillium Expansum in Apples through Iron Depletion. Postharvest Biol. Technol. 2008, 49, 121–128. [Google Scholar] [CrossRef]
- Magliani, W.; Conti, S.; Gerloni, M.; Bertolotti, D.; Polonelli, L. Yeast Killer Systems. Clin. Microbiol. Rev. 1997, 10, 369–400. [Google Scholar] [CrossRef] [PubMed]
- Pawlikowska, E.; James, S.A.; Breierova, E.; Antolak, H.; Kregiel, D. Biocontrol Capability of Local Metschnikowia Sp. Isolates. Antonie van Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2019, 112, 1425–1445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández de Ullivarri, M.; Mendoza, L.M.; Raya, R.R. Killer Yeasts as Biocontrol Agents of Spoilage Yeasts and Bacteria Isolated from Wine. BIO Web Conf. 2014, 3, 02001. [Google Scholar] [CrossRef]
- Al-Qaysi, S.A.S.; Al-Haideri, H.; Thabit, Z.A.; Al-Kubaisy, W.H.A.A.R.; Ibrahim, J.A.A.R. Production, Characterization, and Antimicrobial Activity of Mycocin Produced by Debaryomyces Hansenii DSMZ70238. Int. J. Microbiol. 2017, 2017, 2605382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miranda, C.; Bettencourt, S.; Pozdniakova, T.; Pereira, J.; Sampaio, P.; Franco-Duarte, R.; Pais, C. Modified High-Throughput Nile Red Fluorescence Assay for the Rapid Screening of Oleaginous Yeasts Using Acetic Acid as Carbon Source. BMC Microbiol. 2020, 20, 12866. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hicks, R.H.; Moreno-Beltrán, M.; Gore-Lloyd, D.; Chuck, C.J.; Henk, D.A. The Oleaginous Yeast Metschnikowia pulcherrima Displays Killer Activity against Avian-Derived Pathogenic Bacteria. Biology 2021, 10, 1227. https://doi.org/10.3390/biology10121227
Hicks RH, Moreno-Beltrán M, Gore-Lloyd D, Chuck CJ, Henk DA. The Oleaginous Yeast Metschnikowia pulcherrima Displays Killer Activity against Avian-Derived Pathogenic Bacteria. Biology. 2021; 10(12):1227. https://doi.org/10.3390/biology10121227
Chicago/Turabian StyleHicks, Robert H., Mauro Moreno-Beltrán, Deborah Gore-Lloyd, Christopher J. Chuck, and Daniel A. Henk. 2021. "The Oleaginous Yeast Metschnikowia pulcherrima Displays Killer Activity against Avian-Derived Pathogenic Bacteria" Biology 10, no. 12: 1227. https://doi.org/10.3390/biology10121227
APA StyleHicks, R. H., Moreno-Beltrán, M., Gore-Lloyd, D., Chuck, C. J., & Henk, D. A. (2021). The Oleaginous Yeast Metschnikowia pulcherrima Displays Killer Activity against Avian-Derived Pathogenic Bacteria. Biology, 10(12), 1227. https://doi.org/10.3390/biology10121227