Hazardous Effects of SiO2 Nanoparticles on Liver and Kidney Functions, Histopathology Characteristics, and Transcriptomic Responses in Nile Tilapia (Oreochromis niloticus) Juveniles

 ,
,  ,
, 
 ,
,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Silicon Dioxide Nanoparticles (SiO2NPs)
2.1.1. Preparation and Characterization Methods
2.1.2. SiO2NP Stock Solution
2.2. Animal Ethics
2.3. Fish Procurement, Acclimation, and Rearing
2.4. Water Quality Parameters
2.5. Fish Exposure to SiO2NPs and Experimental Setup
2.6. Sampling
2.6.1. Serum Samples
2.6.2. Tissue Samples
2.7. Serum Biochemical Measurements
2.8. Gene Transcription
2.8.1. Extraction of RNA and Synthesis of cDNA
2.8.2. qRT-PCR
2.9. Histopathological Studies
2.10. Statistical Analytics
3. Results
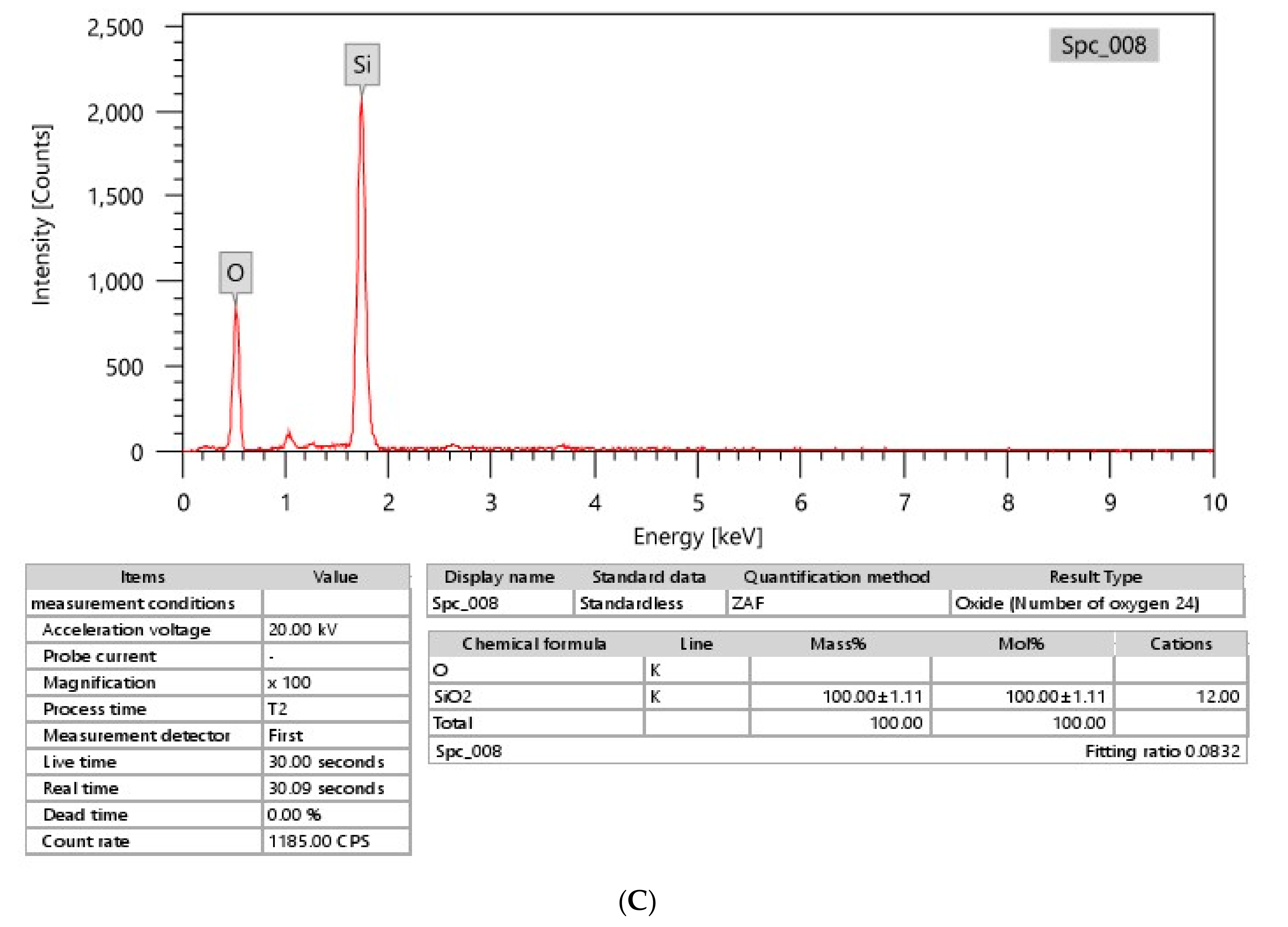
3.1. SiO2NP Characterizations
3.2. Serum Hepato-Renal Functions
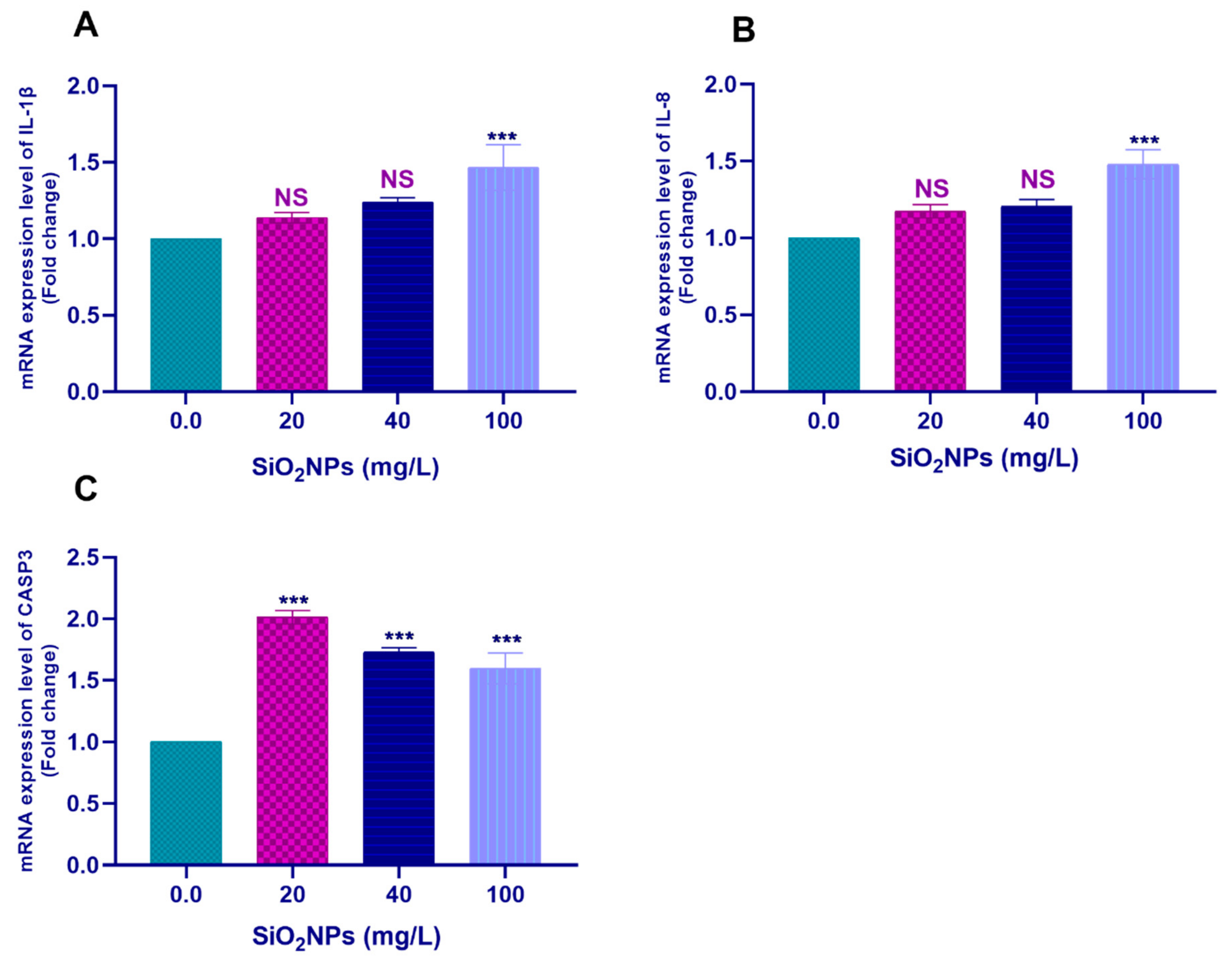
3.3. Gene Transcription Profile
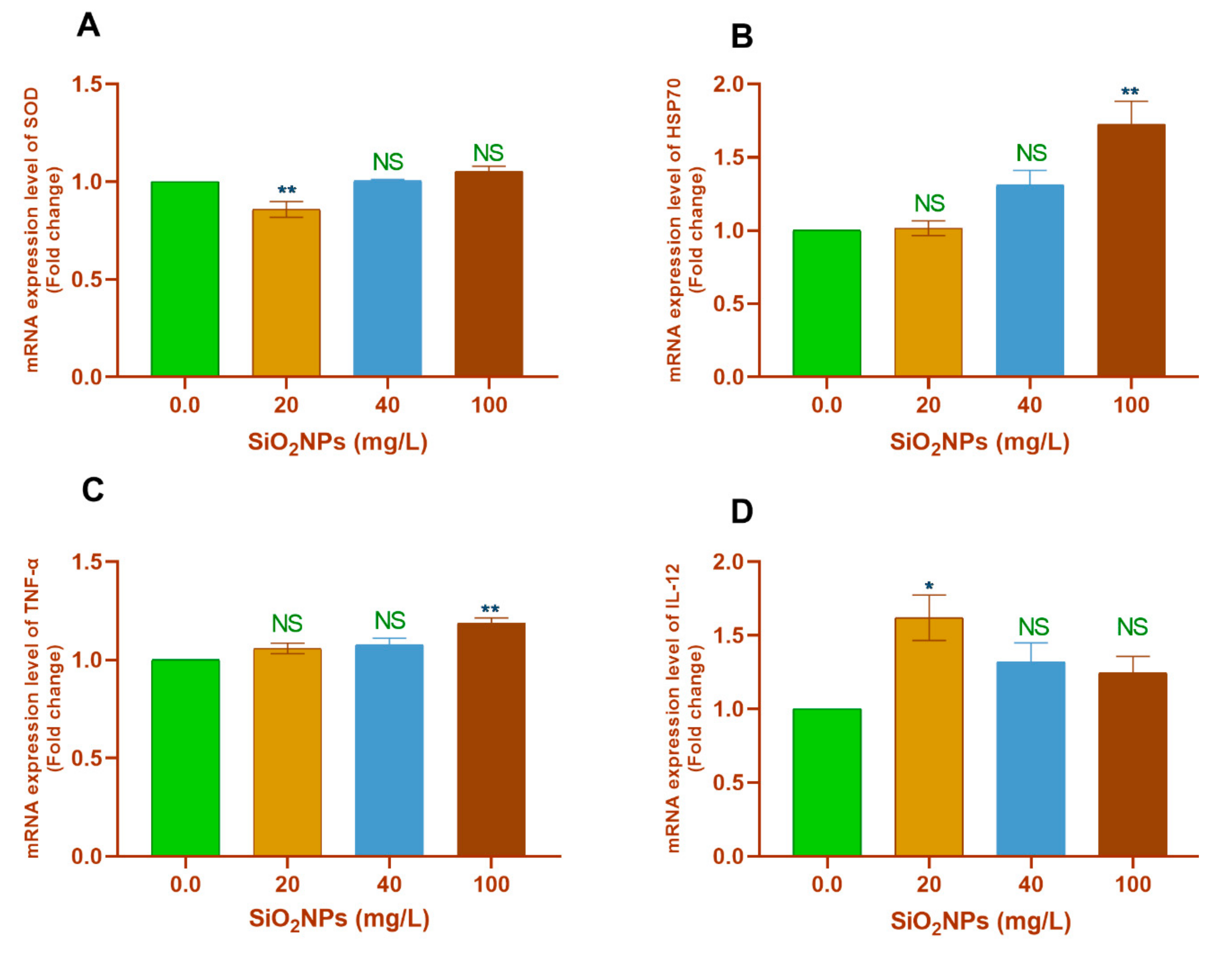
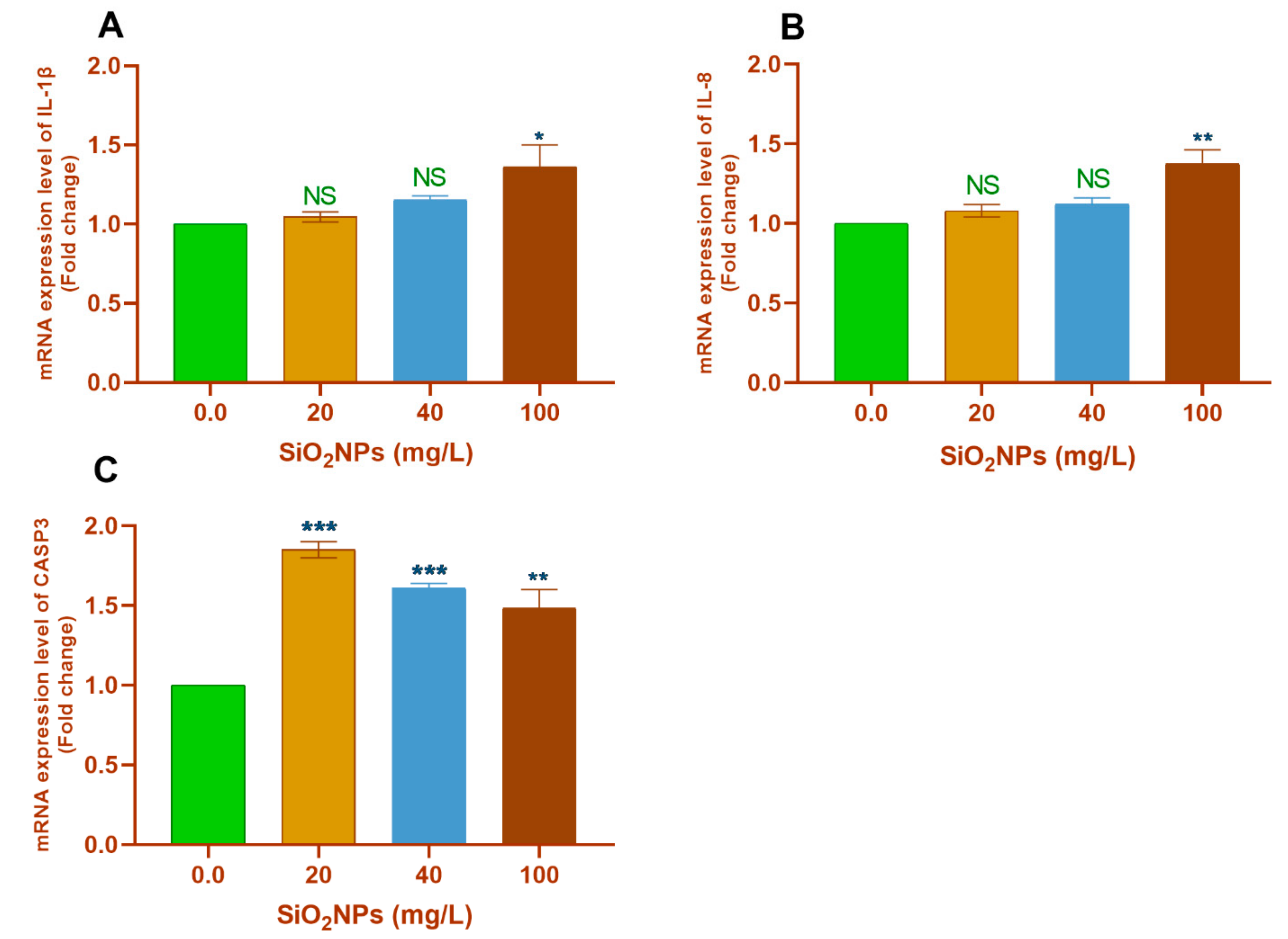
3.3.1. Gill Tissues
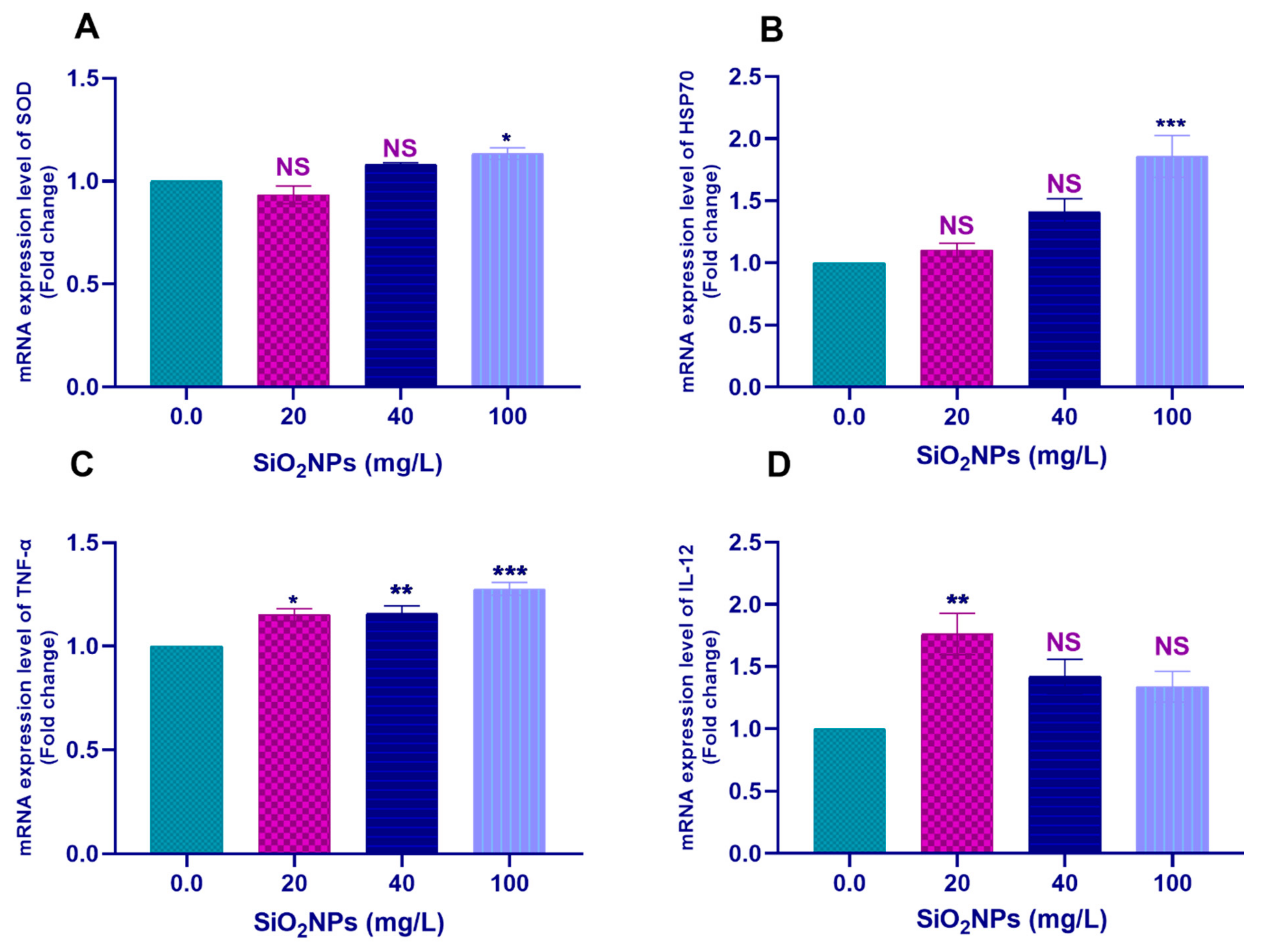
3.3.2. Hepatic Tissues
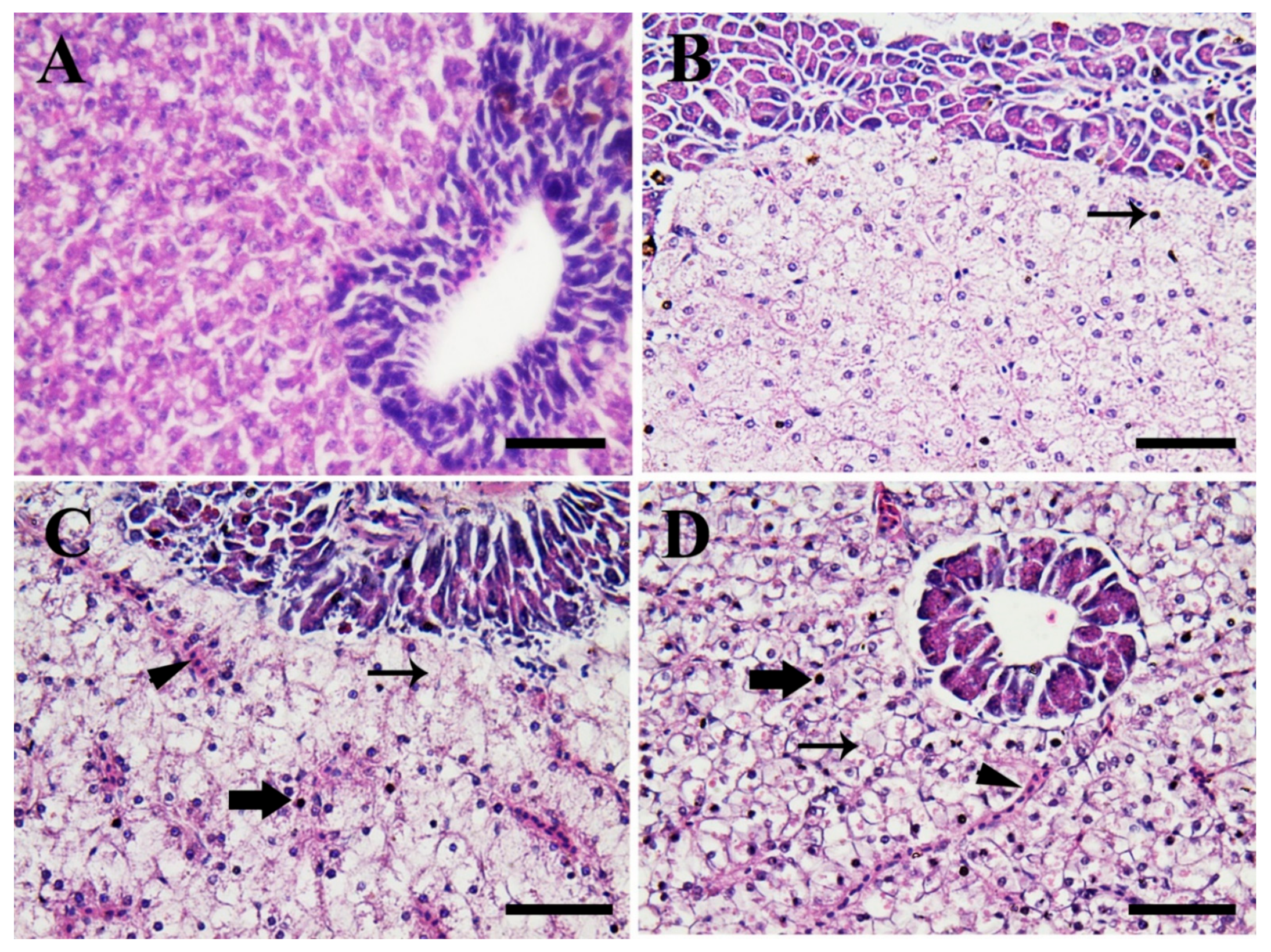
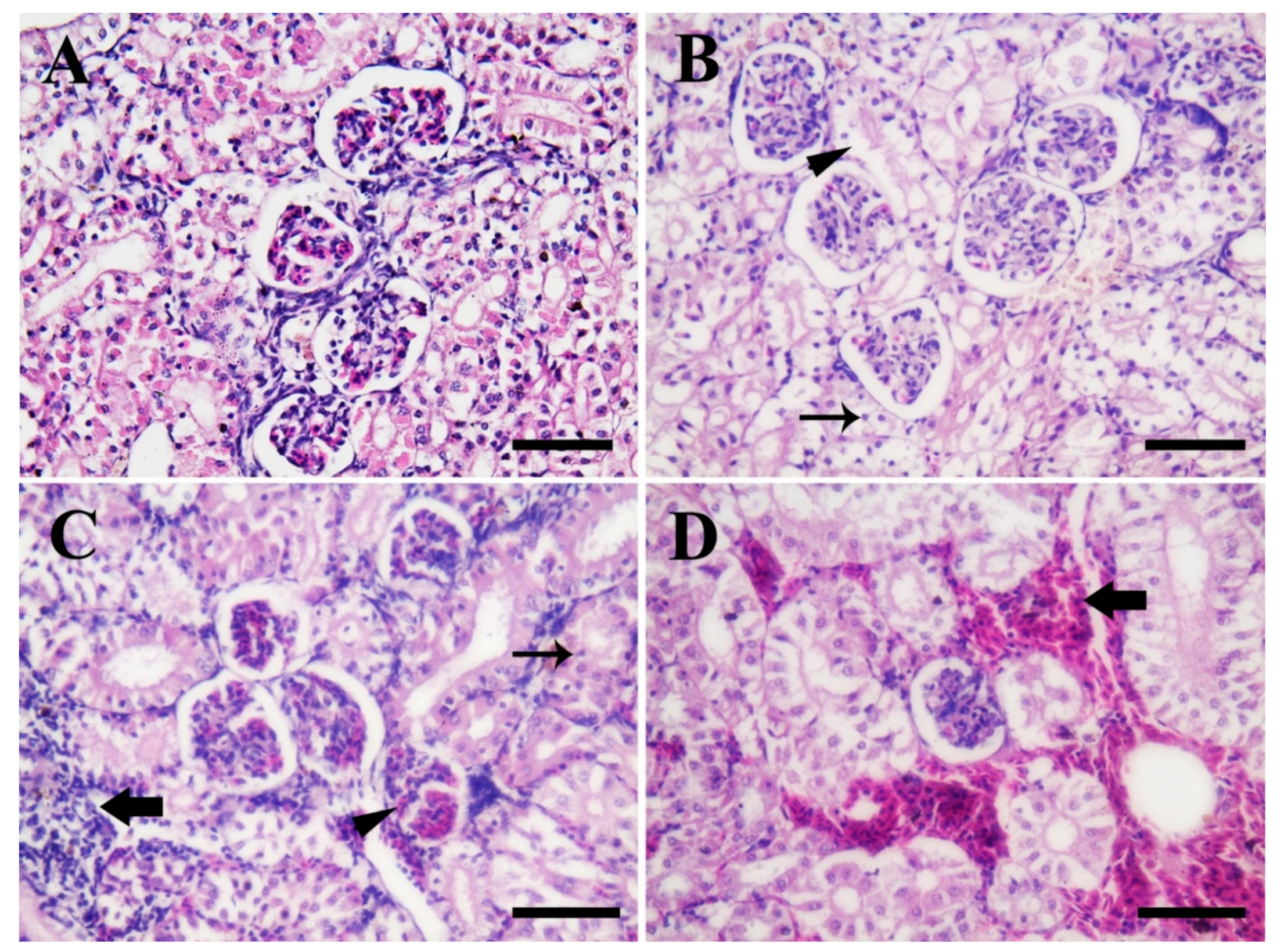
3.4. Histopathological Findings
3.4.1. Hepatopancreas
3.4.2. Gills
3.4.3. Posterior Kidney
4. Discussion
4.1. Hepato-Renal Functions
4.2. Gene Transcription Profile
4.3. Histopathological Alterations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Animal Welfare Statement
Appendix A
References
- Radad, K.; Al-Shraim, M.; Moldzio, R.; Rausch, W.-D. Recent advances in benefits and hazards of engineered nanoparticles. Environ. Toxicol. Pharmacol. 2012, 34, 661–672. [Google Scholar] [CrossRef] [PubMed]
- Baker, T.J.; Tyler, C.R.; Galloway, T.S. Impacts of metal and metal oxide nanoparticles on marine organisms. Environ. Pollut. 2014, 186, 257–271. [Google Scholar] [CrossRef]
- Nel, A.; Xia, T.; Mädler, L.; Li, N. Toxic Potential of Materials at the Nanolevel. Science 2006, 311, 622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handy, R.D.; Owen, R.; Valsami-Jones, E. The ecotoxicology of nanoparticles and nanomaterials: Current status, knowledge gaps, challenges, and future needs. Ecotoxicology 2008, 17, 315–325. [Google Scholar] [CrossRef]
- Abdel-Daim, M.M.; Eissa, I.A.M.; Abdeen, A.; Abdel-Latif, H.M.R.; Ismail, M.; Dawood, M.A.O.; Hassan, A.M. Lycopene and resveratrol ameliorate zinc oxide nanoparticles-induced oxidative stress in Nile tilapia, Oreochromis niloticus. Environ. Toxicol. Pharmacol. 2019, 69, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Latif, H.M.R.; Dawood, M.A.O.; Menanteau-Ledouble, S.; El-Matbouli, M. Environmental transformation of n-TiO2 in the aquatic systems and their ecotoxicity in bivalve mollusks: A systematic review. Ecotoxicol. Environ. Saf. 2020, 200, 110776. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, H.; Godin, B. Biocompatibility assessment of Si-based nano- and micro-particles. Adv. Drug Deliv. Rev. 2012, 64, 1800–1819. [Google Scholar] [CrossRef] [Green Version]
- Santos, H.A.; Mäkilä, E.; Airaksinen, A.J.; Bimbo Luis, M.; Hirvonen, J. Porous silicon nanoparticles for nanomedicine: Preparation and biomedical applications. Nanomedicine 2014, 9, 535–554. [Google Scholar] [CrossRef] [PubMed]
- Rastogi, A.; Tripathi, D.K.; Yadav, S.; Chauhan, D.K.; Živčák, M.; Ghorbanpour, M.; El-Sheery, N.I.; Brestic, M. Application of silicon nanoparticles in agriculture. 3 Biotech. 2019, 9, 90. [Google Scholar] [CrossRef] [Green Version]
- Mizutani, T.; Arai, K.; Miyamoto, M.; Kimura, Y. Application of silica-containing nano-composite emulsion to wall paint: A new environmentally safe paint of high performance. Prog. Org. Coat. 2006, 55, 276–283. [Google Scholar] [CrossRef]
- Tong, X.; Ding, W.; Cetinkaya, C. Effects of Nanoparticle Coating on the Adhesion of Emulsion Aggregation Toner Particles. J. Adhes. Sci. Technol. 2010, 24, 371–387. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.-H.; Bae, H.C.; Jang, Y.; Jeong, S.H.; Lee, H.N.; Ryu, W.-I.; Yoo, M.G.; Kim, Y.-R.; Kim, M.-K.; Lee, J.K. Effect of the size and surface charge of silica nanoparticles on cutaneous toxicity. Mol. Cell. Toxicol. 2013, 9, 67–74. [Google Scholar] [CrossRef]
- Vidya, P.V.; Chitra, K.C. Sublethal effects of silicon dioxide nanoparticles on the structure of gill, liver and brain tissues in the fish, Oreochromis mossambicus (Peters, 1852). Int. J. Appl. Res. 2018, 4, 228–232. [Google Scholar]
- Vidya, P.V.; Chitra, K.C. Irreversible nanotoxicity of silicon dioxide nanoparticles in the freshwater fish Oreochromis mossambicus (Peters, 1852). Asian Fish. Sci. 2018, 31, 146–160. [Google Scholar] [CrossRef]
- Vidya, P.V.; Asifa, K.P.; Chitra, K.C. Hepatic histopathology in Oreochromis mossambicus (Peters, 1852) under silica nanoparticles toxicity. Indian J. Exp. Biol. 2019, 57, 293–296. [Google Scholar]
- Athif, P.; Suganthi, P.; Murali, M.; Sadiq Bukhari, A.; Syed Mohamed, H.E.; Basu, H.; Singhal, R.K. Hepatic toxicological responses of SiO2 nanoparticle on Oreochromis mossambicus. Environ. Toxicol. Pharmacol. 2020, 78, 103398. [Google Scholar] [CrossRef]
- Athif, P.; Murali, M.; Suganthi, P.; Sadiq Bukhari, A.; Syed Mohamed, H.E.; Basu, H.; Singhal, R.K. Alterations in renal markers of tilapia fish exposed to silicon dioxide nanoparticle. Uttar Pradesh J. Zool. 2020, 41, 48–55. [Google Scholar]
- Krishna Priya, K.; Ramesh, M.; Saravanan, M.; Ponpandian, N. Ecological risk assessment of silicon dioxide nanoparticles in a freshwater fish Labeo rohita: Hematology, ionoregulation and gill Na+/K+ ATPase activity. Ecotoxicol. Environ. Saf. 2015, 120, 295–302. [Google Scholar] [CrossRef]
- Ramesh, R.; Kavitha, P.; Kanipandian, N.; Arun, S.; Thirumurugan, R.; Subramanian, P. Alteration of antioxidant enzymes and impairment of DNA in the SiO2 nanoparticles exposed zebra fish (Danio rerio). Environ. Monit. Assess. 2013, 185, 5873–5881. [Google Scholar] [CrossRef] [PubMed]
- Vranic, S.; Shimada, Y.; Ichihara, S.; Kimata, M.; Wu, W.; Tanaka, T.; Boland, S.; Tran, L.; Ichihara, G. Toxicological evaluation of SiO2 nanoparticles by zebrafish embryo toxicity test. Int. J. Mol. Sci. 2019, 20, 882. [Google Scholar] [CrossRef] [Green Version]
- Ye, R.; Yu, X.; Yang, S.; Yuan, J.; Yang, X. Effects of Silica Dioxide Nanoparticles on the Embryonic Development of Zebrafish. Integr. Ferroelectr. 2013, 147, 166–174. [Google Scholar] [CrossRef]
- Miroshnikova, E.; Kosyan, D.; Arizhanov, A.; Sizova, E.; Kalashnikov, V. Assessment of general toxicity and prooxidant effects of CeO2 and SiO2 nanoparticles on Danio rerio. Agric. Biol. 2016, 51, 921. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; He, W.; Hu, S.; Kong, R.; Yang, L. The fate and oxidative stress of different sized SiO2 nanoparticles in zebrafish (Danio rerio) larvae. Chemosphere 2019, 225, 705–712. [Google Scholar] [CrossRef]
- Li, X.; Ji, X.; Wang, R.; Zhao, J.; Dang, J.; Gao, Y.; Jin, M. Zebrafish behavioral phenomics employed for characterizing behavioral neurotoxicity caused by silica nanoparticles. Chemosphere 2020, 240, 124937. [Google Scholar] [CrossRef]
- Stöber, W.; Fink, A.; Bohn, E. Controlled growth of monodisperse silica spheres in the micron size range. J. Colloid Interface Sci. 1968, 26, 62–69. [Google Scholar] [CrossRef]
- National Research Council (NRC). Nutrient Requirements of Fish; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Boyd, C.E.; Tucker, C.S. Pond Aquaculture Water Quality Management; Kluwer Academic Publishers: Boston, MA, USA, 2012. [Google Scholar]
- Finney, D.J. Probit Analysis: A Statistical Treatment of the Sigmoid Response Curve; Cambridge University Press: Cambridge, UK, 1952. [Google Scholar]
- Coulombe, J.; Favreau, L. A new simple semimicro method for colorimetric determination of urea. Clin. Chem. 1963, 9, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Larsen, K. Creatinine assay by a reaction-kinetic principle. Clin. Chim. Acta 1972, 41, 209–217. [Google Scholar] [CrossRef]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Tietz, N.; Burtis, C.; Duncan, P.; Ervin, K.; Petitclerc, C.; Rinker, A.; Shuey, D.; Zygowicz, E. A reference method for measurement of alkaline phosphatase activity in human serum. Clin. Chem. 1983, 29, 751–761. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. The Hematoxylin and eosin. In Theory and Practice of Histological Techniques, 7th ed.; Suvarna, S.K., Layton, C., Bancroft, J.D., Eds.; Churchill Livingstone: Edinburgh, UK; New York, NY, USA, 2013; pp. 179–220. [Google Scholar]
- Khafaga, A.F.; Naiel, M.A.E.; Dawood, M.A.O.; Abdel-Latif, H.M.R. Dietary Origanum vulgare essential oil attenuates cypermethrin-induced biochemical changes, oxidative stress, histopathological alterations, apoptosis, and reduces DNA damage in Common carp (Cyprinus carpio). Aquat. Toxicol. 2020, 228, 105624. [Google Scholar] [CrossRef]
- Abdel-Latif, H.M.R.; Abdel-Tawwab, M.; Khafaga, A.F.; Dawood, M.A.O. Dietary oregano essential oil improved the growth performance via enhancing the intestinal morphometry and hepato-renal functions of common carp (Cyprinus carpio L.) fingerlings. Aquaculture 2020, 526, 735432. [Google Scholar] [CrossRef]
- Ismael, N.E.M.; Abd El-hameed, S.A.A.; Salama, A.M.; Naiel, M.A.E.; Abdel-Latif, H.M.R. The effects of dietary clinoptilolite and chitosan nanoparticles on growth, body composition, haemato-biochemical parameters, immune responses, and antioxidative status of Nile tilapia exposed to imidacloprid. Environ. Sci. Pollut. Res. 2021, 10. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abdel-Tawwab, M.; Abdel-Latif, H.M.R. Lycopene reduces the impacts of aquatic environmental pollutants and physical stressors in fish. Rev. Aquac. 2020, 12, 2511–2526. [Google Scholar] [CrossRef]
- Kumar, N.; Krishnani, K.K.; Singh, N.P. Comparative study of selenium and selenium nanoparticles with reference to acute toxicity, biochemical attributes, and histopathological response in fish. Environ. Sci. Pollut. Res. 2018, 25, 8914–8927. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.T.A. Toxicological impact of green synthesized silver nanoparticles and protective role of different selenium type on Oreochromis niloticus: Hematological and biochemical response. J. Trace Elem. Med. Biol. 2020, 61, 126507. [Google Scholar] [CrossRef] [PubMed]
- Basu, N.; Todgham, A.E.; Ackerman, P.A.; Bibeau, M.R.; Nakano, K.; Schulte, P.M.; Iwama, G.K. Heat shock protein genes and their functional significance in fish. Gene 2002, 295, 173–183. [Google Scholar] [CrossRef]
- Iwama, G.K.; Thomas, P.T.; Forsyth, R.B.; Vijayan, M.M. Heat shock protein expression in fish. Rev. Fish Biol. Fish. 1998, 8, 35–56. [Google Scholar] [CrossRef]
- Iwama, G.K.; Vijayan, M.M.; Forsyth, R.B.; Ackerman, P.A. Heat shock proteins and physiological stress in fish. Am. Zool. 1999, 39, 901–909. [Google Scholar] [CrossRef] [Green Version]
- Goetz, F.W.; Planas, J.V.; MacKenzie, S. Tumor necrosis factors. Dev. Comp. Immunol. 2004, 28, 487–497. [Google Scholar] [CrossRef]
- Secombes, C.J.; Hardie, L.J.; Daniels, G. Cytokines in fish: An update. Fish Shellfish Immunol. 1996, 6, 291–304. [Google Scholar] [CrossRef]
- Savan, R.; Sakai, M. Genomics of fish cytokines. Comp. Biochem. Physiol. Part D Genom. Proteom. 2006, 1, 89–101. [Google Scholar] [CrossRef]
- Reyes-Cerpa, S.; Maisey, K.; Reyes-López, F.; Toro-Ascuy, D.; Sandino, A.M.; Imarai, M. Fish cytokines and immune response. In New Advances and Contributions to Fish Biology; Narendra Publishing House: Delhi, India, 2012; pp. 3–57. [Google Scholar]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Le Bras, M.; Clément, M.V.; Pervaiz, S.; Brenner, C. Reactive oxygen species and the mitochondrial signaling pathway of cell death. Histol. Histopathol. 2005, 20, 205–219. [Google Scholar] [PubMed]
- Di Giulio, R.T.; Washburn, P.C.; Wenning, R.J.; Winston, G.W.; Jewell, C.S. Biochemical responses in aquatic animals: A review of determinants of oxidative stress. Environ. Toxicol. Chem. 1989, 8, 1103–1123. [Google Scholar] [CrossRef]
- Fang, Y.-Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Rotruck, J.T.; Pope, A.L.; Ganther, H.E.; Swanson, A.B.; Hafeman, D.G.; Hoekstra, W.G. Selenium: Biochemical role as a component of glutathione peroxidase. Science 1973, 179, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Ren, X.; Zhu, R.; Luo, Z.; Ren, B. Zinc oxide nanoparticles induce oxidative DNA damage and ROS-triggered mitochondria-mediated apoptosis in zebrafish embryos. Aquat. Toxicol. 2016, 180, 56–70. [Google Scholar] [CrossRef]
- Fırat, Ö.; Bozat, R.C. Assessment of biochemical and toxic responses induced by titanium dioxide nanoparticles in Nile tilapia Oreochromis niloticus. Hum. Ecol. Risk Assess. Int. J. 2019, 25, 1438–1447. [Google Scholar] [CrossRef]
- Chen, M.; Yin, J.; Liang, Y.; Yuan, S.; Wang, F.; Song, M.; Wang, H. Oxidative stress and immunotoxicity induced by graphene oxide in zebrafish. Aquat. Toxicol. 2016, 174, 54–60. [Google Scholar] [CrossRef]
- Teles, M.; Reyes-López, F.E.; Fierro-Castro, C.; Tort, L.; Soares, A.M.V.M.; Oliveira, M. Modulation of immune genes mRNA levels in mucosal tissues and DNA damage in red blood cells of Sparus aurata by gold nanoparticles. Mar. Pollut. Bull. 2018, 133, 428–435. [Google Scholar] [CrossRef]
- Wang, T.; Long, X.; Liu, Z.; Cheng, Y.; Yan, S. Effect of copper nanoparticles and copper sulphate on oxidation stress, cell apoptosis and immune responses in the intestines of juvenile Epinephelus coioides. Fish Shellfish Immunol. 2015, 44, 674–682. [Google Scholar] [CrossRef] [PubMed]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Jänicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.; Wen, X.; Hu, Y.; Zhang, X.; Wang, D.; Yin, S. Copper nanoparticles induced oxidation stress, cell apoptosis and immune response in the liver of juvenile Takifugu fasciatus. Fish Shellfish Immunol. 2019, 84, 648–655. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.W.; Choi, Y.C.; Kim, R.; Lee, S.K. Multiwall carbon nanotube-induced apoptosis and antioxidant gene expression in the gills, liver, and intestine of Oryzias latipes. Biomed Res. Int. 2015, 2015, 485343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dawood, M.A.O.; El-Salam Metwally, A.; Elkomy, A.H.; Gewaily, M.S.; Abdo, S.E.; Abdel-Razek, M.A.S.; Soliman, A.A.; Amer, A.A.; Abdel-Razik, N.I.; Abdel-Latif, H.M.R.; et al. The impact of menthol essential oil against inflammation, immunosuppression, and histopathological alterations induced by chlorpyrifos in Nile tilapia. Fish Shellfish Immunol. 2020, 102, 316–325. [Google Scholar] [CrossRef]
- El Euony, O.I.; Elblehi, S.S.; Abdel-Latif, H.M.; Abdel-Daim, M.M.; El-Sayed, Y.S. Modulatory role of dietary Thymus vulgaris essential oil and Bacillus subtilis against thiamethoxam-induced hepatorenal damage, oxidative stress, and immunotoxicity in African catfish (Clarias garipenus). Environ. Sci. Pollut. Res. 2020, 27, 23108–23128. [Google Scholar] [CrossRef]
- Abdel-Latif, H.; Abou Khashaba, A. Subchronic toxicity of Nile tilapia with different exposure routes to Microcystis aeruginosa: Histopathology, liver functions, and oxidative stress biomarkers. Vet. World 2017, 10, 955. [Google Scholar] [CrossRef] [Green Version]
- Kaya, H.; Duysak, M.; Akbulut, M.; Yılmaz, S.; Gürkan, M.; Arslan, Z.; Demir, V.; Ateş, M. Effects of subchronic exposure to zinc nanoparticles on tissue accumulation, serum biochemistry, and histopathological changes in tilapia (Oreochromis niloticus). Environ. Toxicol. 2017, 32, 1213–1225. [Google Scholar] [CrossRef] [Green Version]
- Murali, M.; Suganthi, P.; Athif, P.; Sadiq Bukhari, A.; Syed Mohamed, H.E.; Basu, H.; Singhal, R.K. Histological alterations in the hepatic tissues of Al2O3 nanoparticles exposed freshwater fish Oreochromis mossambicus. J. Trace Elem. Med. Biol. 2017, 44, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Bernet, D.; Schmidt, H.; Meier, W.; Burkhardt-Holm, P.; Wahli, T. Histopathology in fish: Proposal for a protocol to assess aquatic pollution. J. Fish Dis. 1999, 22, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Campos-Garcia, J.; Martinez, D.S.T.; Rezende, K.F.O.; da Silva, J.R.M.C.; Alves, O.L.; Barbieri, E. Histopathological alterations in the gills of Nile tilapia exposed to carbofuran and multiwalled carbon nanotubes. Ecotoxicol. Environ. Saf. 2016, 133, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Aydın, F.; Gürkan, M.; Yılmaz, S.; Ates, M.; Demir, V.; Arslan, Z. A comparative toxicity study between small and large size zinc oxide nanoparticles in tilapia (Oreochromis niloticus): Organ pathologies, osmoregulatory responses and immunological parameters. Chemosphere 2016, 144, 571–582. [Google Scholar] [CrossRef]
- Hao, L.; Wang, Z.; Xing, B. Effect of sub-acute exposure to TiO2 nanoparticles on oxidative stress and histopathological changes in Juvenile Carp (Cyprinus carpio). J. Environ. Sci. 2009, 21, 1459–1466. [Google Scholar] [CrossRef]
- Gupta, Y.R.; Sellegounder, D.; Kannan, M.; Deepa, S.; Senthilkumaran, B.; Basavaraju, Y. Effect of copper nanoparticles exposure in the physiology of the common carp (Cyprinus carpio): Biochemical, histological and proteomic approaches. Aquac. Fish. 2016, 1, 15–23. [Google Scholar] [CrossRef] [Green Version]
- Chupani, L.; Niksirat, H.; Velíšek, J.; Stará, A.; Hradilová, Š.; Kolařík, J.; Panáček, A.; Zusková, E. Chronic dietary toxicity of zinc oxide nanoparticles in common carp (Cyprinus carpio L.): Tissue accumulation and physiological responses. Ecotoxicol. Environ. Saf. 2018, 147, 110–116. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target mRNA | Primer Sequences (F: Forward, R: Reverse) | NCBI GenBank Accession No. | Slope | Efficiency % |
|---|---|---|---|---|
| SOD | F: 5′-CCCTACGTCAGTGCAGAGAT-3′ | JF801727.1 | −3.51 | 92.708% |
| R: 5′-GTCACGTCTCCCTTTGCAAG-3′ | ||||
| GPX | F: 5′-CGCCGAAGGTCTCGTTATTT-3′ | NM_001279711.1 | −3.55 | 91.289% |
| R: 5′-TCCCTGGACGGACATACTT-3′ | ||||
| CAT | F: 5′-CCCAGCTCTTCATCCAGAAAC-3′ | JF801726.1 | −3.41 | 96.451% |
| R: 5′-GCCTCCGCATTGTACTTCTT-3′ | ||||
| HSP70 | F: 5′-CATCGCCTACGGTCTGGACAA-3′ | FJ207463.1 | −3.60 | 89.574% |
| R: 5′-TGCCGTCTTCAATGGTCAGGAT-3′ | ||||
| TNF-α | F: 5′-GGAAGCAGCTCCACTCTGATGA-3′ | JF957373.1 | −3.33 | 99.664% |
| R: 5′-CACAGCGTGTCTCCTTCGTTCA-3′ | ||||
| IL-1β | F: 5′-CAAGGATGACGACAAGCCAACC-3′ | XM_003460625.2 | −3.65 | 87.92% |
| R: 5′-AGCGGACAGACATGAGAGTGC-3′ | ||||
| IL-8 | F: 5′-TCATTGTCAGCTCCATCGTG-3′ | NM_001279704.1 | −3.62 | 88.905% |
| R: 5′-CCTGTCCTTTTCAGTGTGGC-3′ | ||||
| CASP3 | F: 5′-GGCTCTTCGTCTGCTTCTGT-3′ | GQ421464.1 | −3.36 | 98.435% |
| R: 5′-GGGAAATCGAGGCGGTATCT-3′ | ||||
| IL-10 | F: 5′-CTGCTAGATCAGTCCGTCGAA-3′ | XM_003441366.2 | −3.44 | 95.298% |
| R: 5′-GCAGAACCGTGTCCAGGTAA-3′ | ||||
| IL-12 | F: 5′-GGGTGCGAGTCAGCTATGAG-3′ | XM_003437924.4 | −3.59 | 89.912% |
| R: 5′-GGTTGTGGATTGGTTGCGTC-3′ | ||||
| β-actin | F: 5′-CCACACAGTGCCCATCTACGA-3′ | EU887951.1 | −3.522 | 92.278% |
| R: 5′-CCACGCTCTGTCAGGATCTTCA-3′ |
| Parameters | SiO2NP Concentrations | |||
|---|---|---|---|---|
| Control (0.0 mg/L) | 20 mg/L | 40 mg/L | 100 mg/L | |
| Liver function enzymes | ||||
| AST (U/L) | 44.91 ± 0.88 d | 51.84 ± 2.00 c | 61.23 ± 2.30 b | 84.27 ± 1.50 a |
| ALT (U/L) | 13.28 ± 0.61 c | 14.28 ± 0.99 b | 16.95 ± 1.35 b | 22.61 ± 1.51 a |
| ALP (U/L) | 9.62 ± 0.78 b | 11.17 ± 1.20 ab | 11.20 ± 0.58 ab | 15.34 ± 1.56 a |
| Kidney function indices | ||||
| Urea (mg/dL) | 6.67 ± 0.55 c | 9.33 ± 0.61 b | 11.42 ± 1.04 b | 16.11 ± 0.86 a |
| Creatinine (mg%) | 0.66 ± 0.06 c | 1.06 ± 0.13 b | 1.12 ± 0.07 ab | 1.65 ± 0.36 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abdel-Latif, H.M.R.; Shukry, M.; El Euony, O.I.; Mohamed Soliman, M.; Noreldin, A.E.; Ghetas, H.A.; Dawood, M.A.O.; Khallaf, M.A. Hazardous Effects of SiO2 Nanoparticles on Liver and Kidney Functions, Histopathology Characteristics, and Transcriptomic Responses in Nile Tilapia (Oreochromis niloticus) Juveniles. Biology 2021, 10, 183. https://doi.org/10.3390/biology10030183
Abdel-Latif HMR, Shukry M, El Euony OI, Mohamed Soliman M, Noreldin AE, Ghetas HA, Dawood MAO, Khallaf MA. Hazardous Effects of SiO2 Nanoparticles on Liver and Kidney Functions, Histopathology Characteristics, and Transcriptomic Responses in Nile Tilapia (Oreochromis niloticus) Juveniles. Biology. 2021; 10(3):183. https://doi.org/10.3390/biology10030183
Chicago/Turabian StyleAbdel-Latif, Hany M.R., Mustafa Shukry, Omnia I. El Euony, Mohamed Mohamed Soliman, Ahmed E. Noreldin, Hanan A. Ghetas, Mahmoud A.O. Dawood, and Mohamed A. Khallaf. 2021. "Hazardous Effects of SiO2 Nanoparticles on Liver and Kidney Functions, Histopathology Characteristics, and Transcriptomic Responses in Nile Tilapia (Oreochromis niloticus) Juveniles" Biology 10, no. 3: 183. https://doi.org/10.3390/biology10030183
APA StyleAbdel-Latif, H. M. R., Shukry, M., El Euony, O. I., Mohamed Soliman, M., Noreldin, A. E., Ghetas, H. A., Dawood, M. A. O., & Khallaf, M. A. (2021). Hazardous Effects of SiO2 Nanoparticles on Liver and Kidney Functions, Histopathology Characteristics, and Transcriptomic Responses in Nile Tilapia (Oreochromis niloticus) Juveniles. Biology, 10(3), 183. https://doi.org/10.3390/biology10030183