
E3 Ubiquitin Ligase ASB17 Promotes Apoptosis by Ubiquitylating and Degrading BCLW and MCL1

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Studies
2.2. Intratesticular Injections
2.3. Cell Lines and Cultures
2.4. Reagents
2.5. Antibodies
2.6. RNA Extraction and Quantitative PCR
2.7. Plasmids and Constructions
2.8. Western Blot Analysis
2.9. Coimmunoprecipitation
2.10. Lentiviral Package and Infection
2.11. Immunofluorescence Microscopy
2.12. In Vivo Ubiquitylation Assay
2.13. TUNEL Assay
2.14. Fertility Evaluation
2.15. Statistics
3. Results
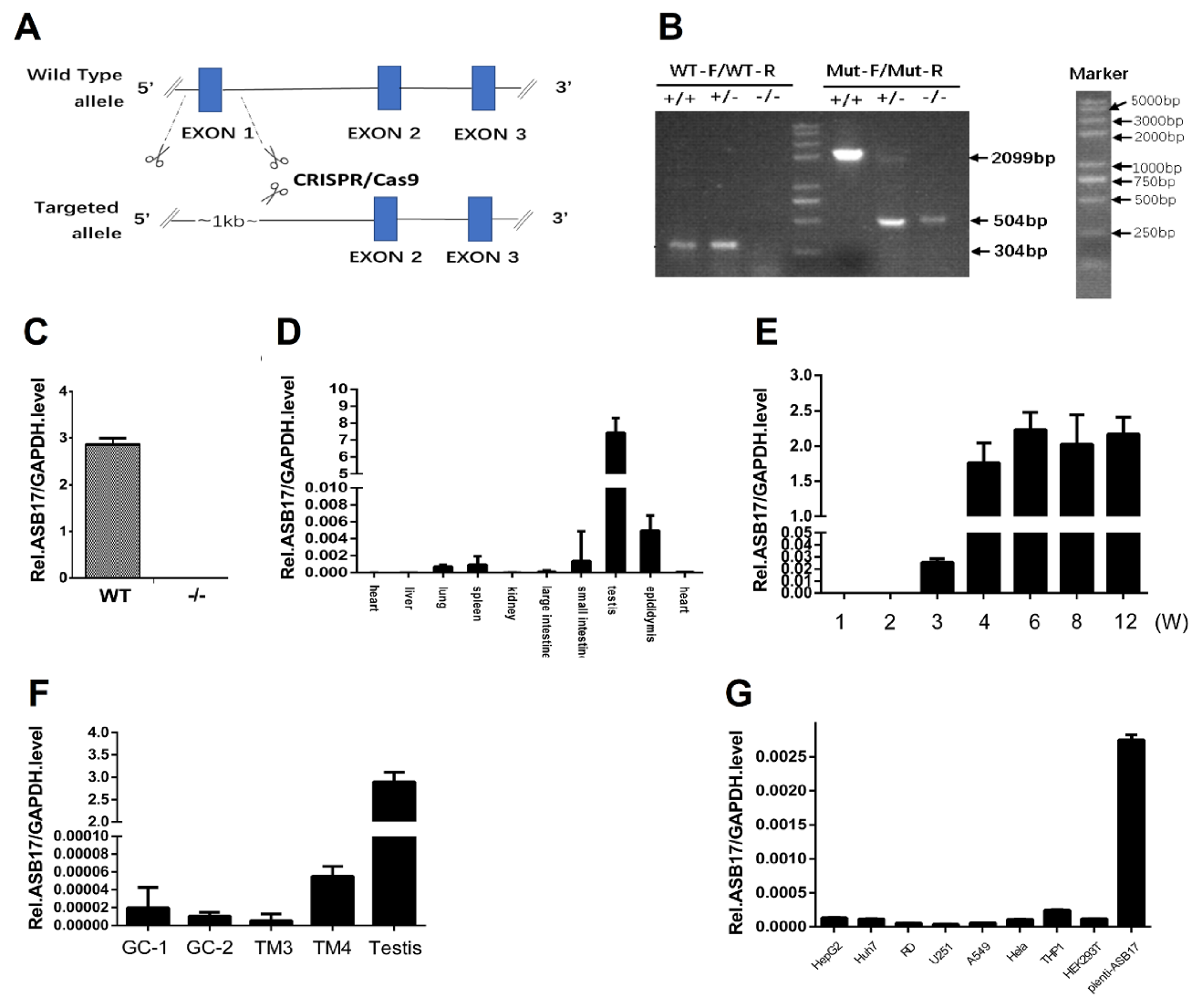
3.1. ASB17 Is Mainly Expressed in the Testis and Epididymis and Slightly Elevated in Spleen and Lung
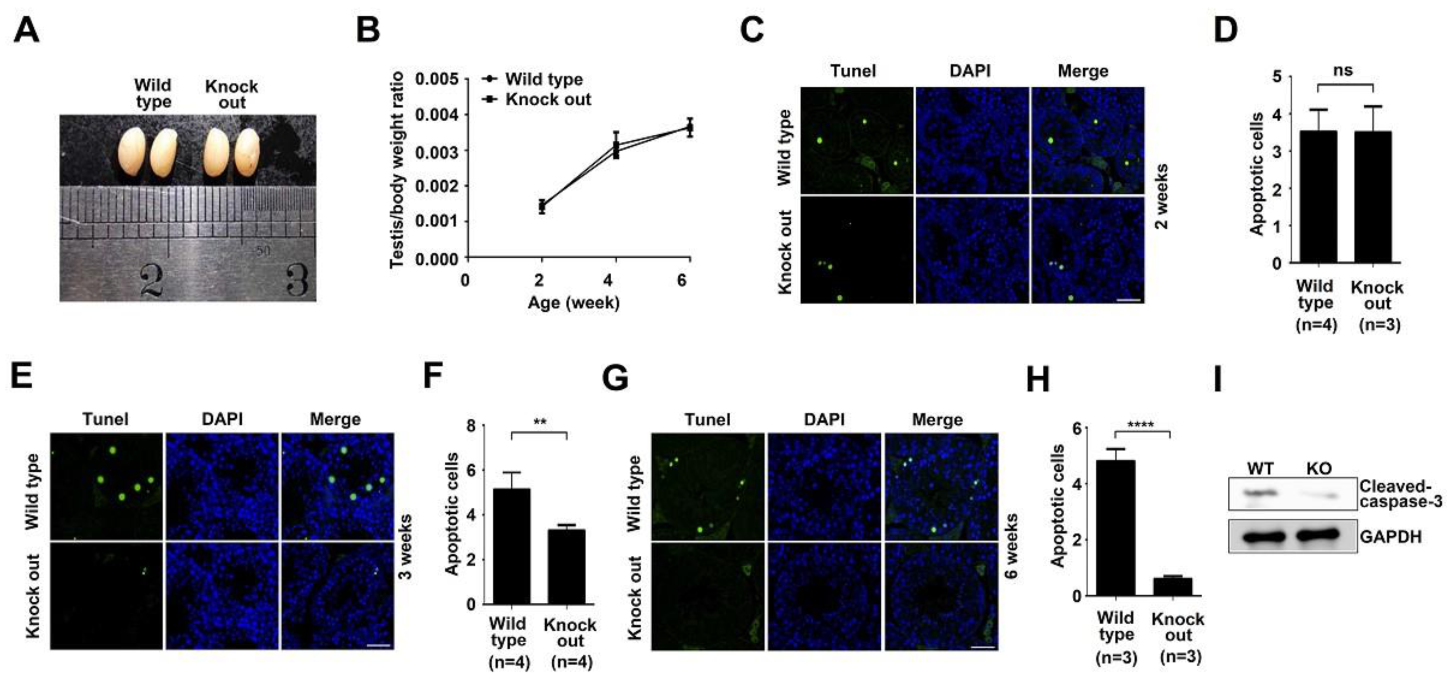
3.2. ASB17 Deficiency in the Testes Attenuated Apoptosis with No Effect on Testes Development
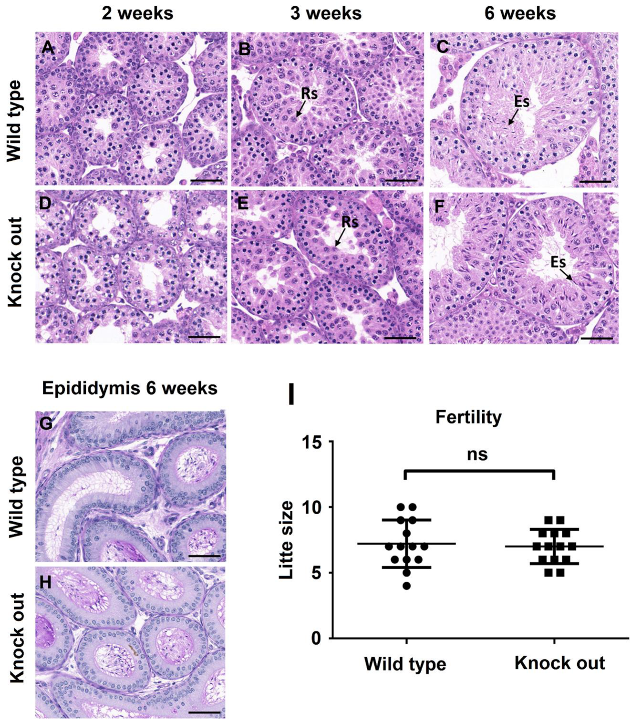
3.3. ASB17−/− Mice Display Normal Spermatogenesis
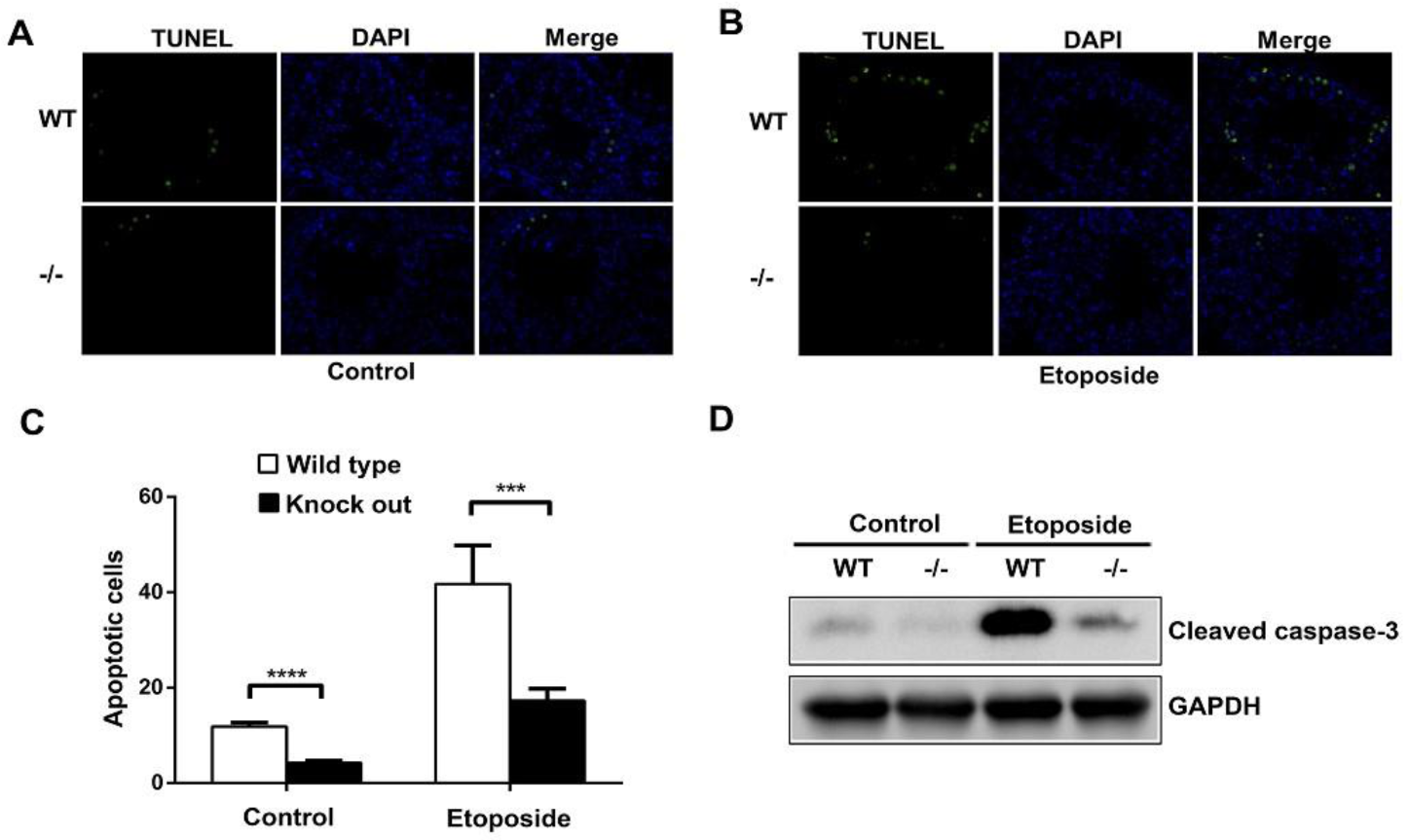
3.4. ASB17 Deficiency Prevents the Apoptosis of Spermatogonia Induced by Etoposide in Male Mice
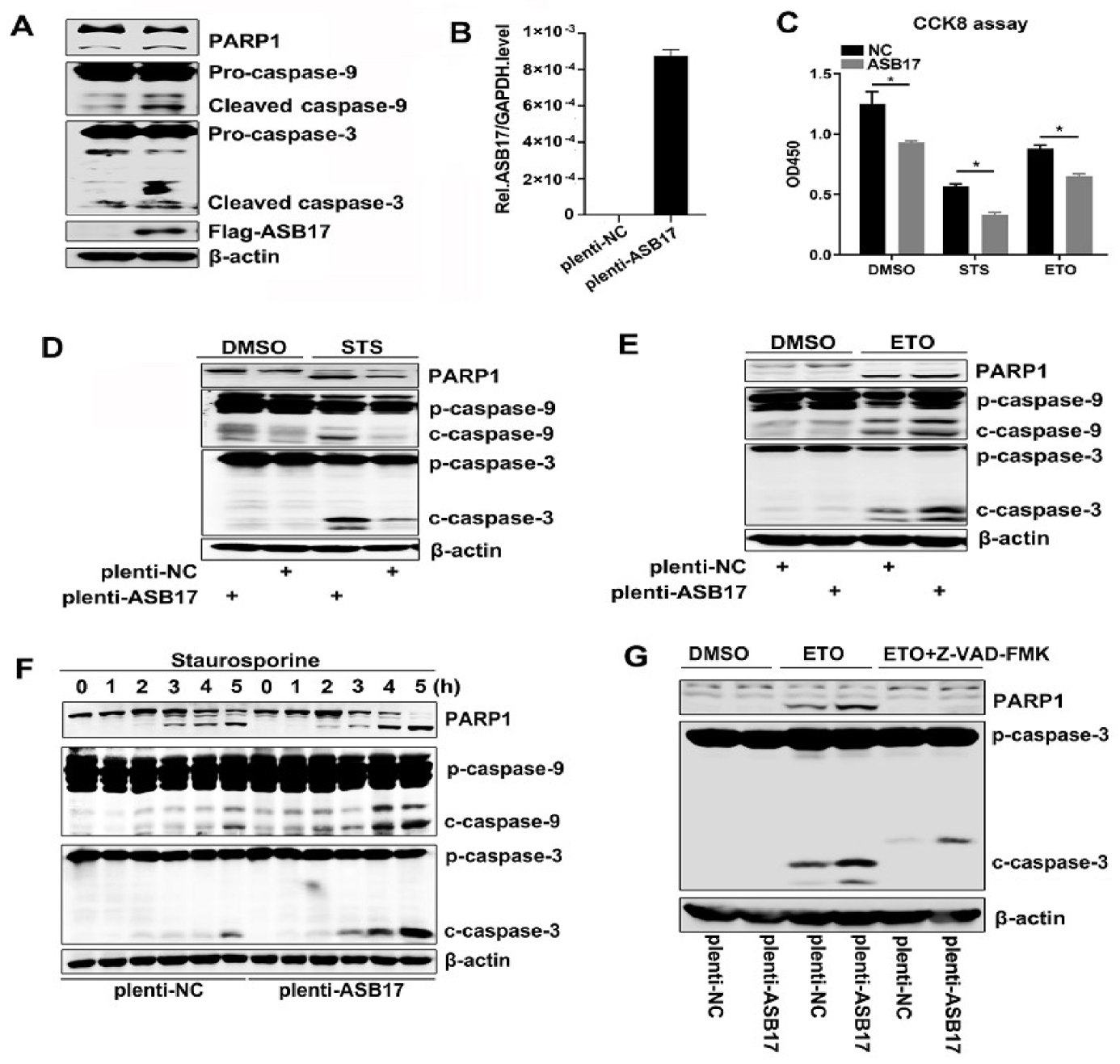
3.5. ASB17 Promotes Apoptosis In Vitro
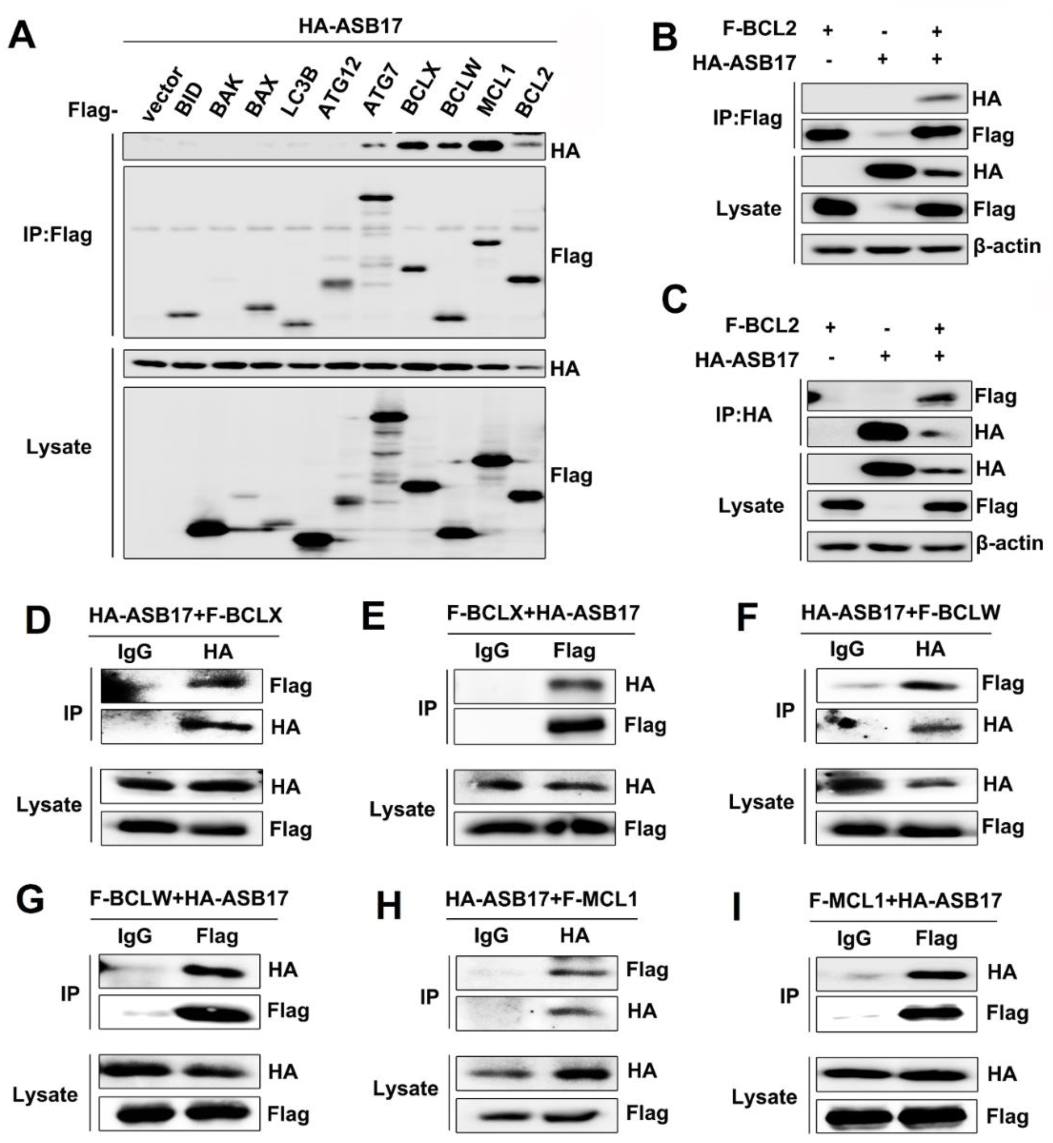
3.6. ASB17 Interacts with BCL2, BCLX, BCLW, and MCL1
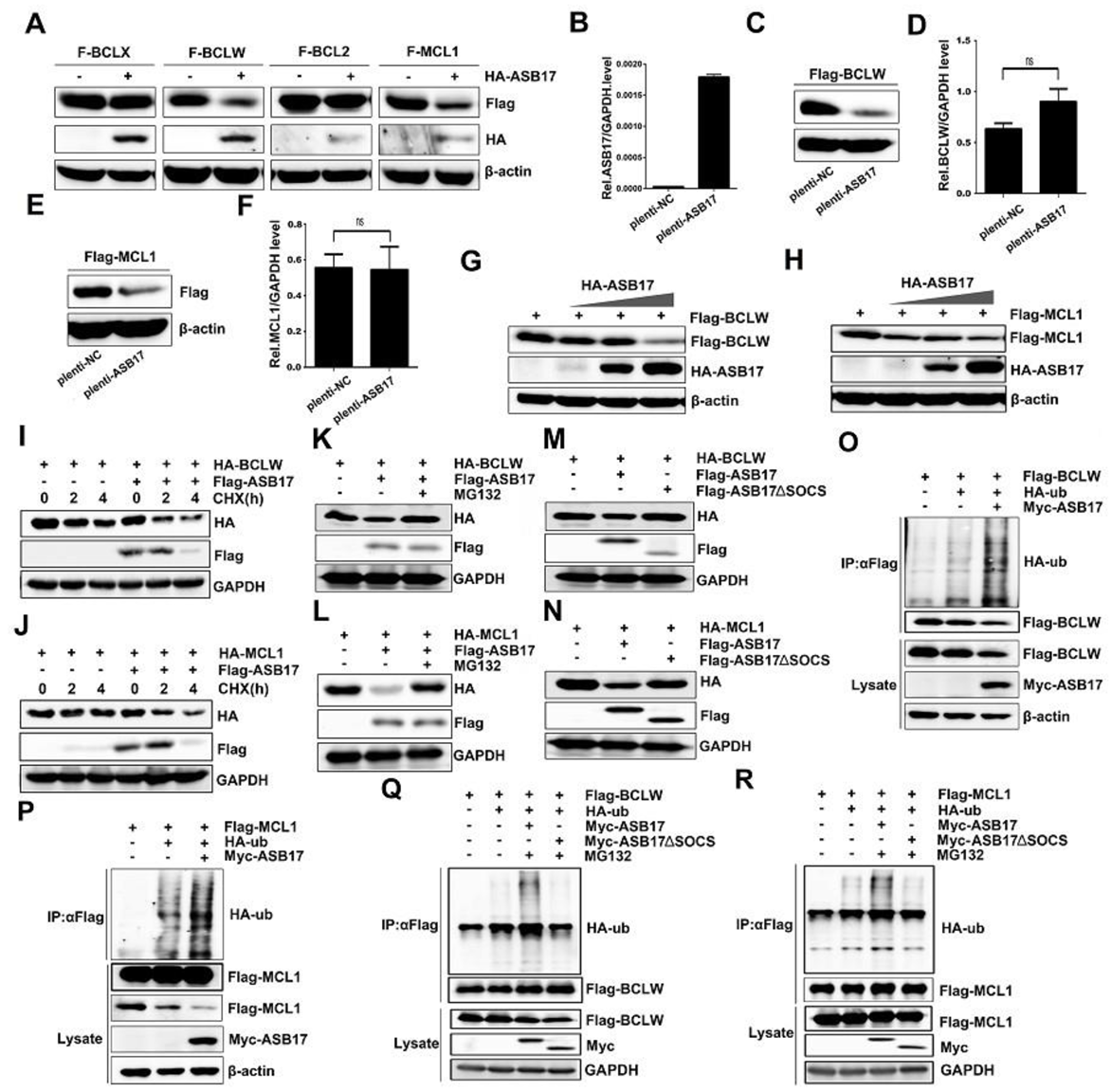
3.7. ASB17 Promotes the Ubiquitylation and Degradation of BCLW and MCL1
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hengartner, M.O. The biochemistry of apoptosis. Nature 2000, 407, 770–776. [Google Scholar] [CrossRef]
- Tait, S.W.G.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef]
- Kerr, J.F.; Wyllie, A.H.; Currie, A.R. Apoptosis: A basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, R.C.; Cullen, S.P.; Martin, S.J. Apoptosis: Controlled demolition at the cellular level. Nat. Rev. Mol. Cell Biol. 2008, 9, 231–241. [Google Scholar] [CrossRef]
- Danial, N.N.; Korsmeyer, S.J. Cell death: Critical control points. Cell 2004, 116, 205–219. [Google Scholar] [CrossRef] [Green Version]
- Hata, A.N.; Engelman, J.A.; Faber, A.C. The BCL2 Family: Key Mediators of the Apoptotic Response to Targeted Anticancer Therapeutics. Cancer Discov. 2015, 5, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Nagata, S. Apoptosis by death factor. Cell 1997, 88, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Zheng, W.; Lin, A.; Uyhazi, K.; Zhao, H.; Lin, H. Pumilio 1 suppresses multiple activators of p53 to safeguard spermatogenesis. Curr. Biol. 2012, 22, 420–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beumer, T.L.; Roepers-Gajadien, H.L.; Gademan, I.S.; van Buul, P.P.; Gil-Gomez, G.; Rutgers, D.H.; de Rooij, D.G. The role of the tumor suppressor p53 in spermatogenesis. Cell Death Differ. 1998, 5, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Knudson, C.M.; Tung, K.S.; Tourtellotte, W.G.; Brown, G.A.; Korsmeyer, S.J. Bax-deficient mice with lymphoid hyperplasia and male germ cell death. Science 1995, 270, 96–99. [Google Scholar] [CrossRef] [PubMed]
- Russell, L.D.; Chiarini-Garcia, H.; Korsmeyer, S.J.; Knudson, C.M. Bax-dependent spermatogonia apoptosis is required for testicular development and spermatogenesis. Biol. Reprod. 2002, 66, 950–958. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Stahl, B.C.; DeWolf, W.C.; Morgentaler, A. P53 and Fas are sequential mechanisms of testicular germ cell apoptosis. J. Androl. 2002, 23, 64–70. [Google Scholar] [CrossRef]
- Lizama, C.; Rojas-Benitez, D.; Antonelli, M.; Ludwig, A.; Moreno, R.D. Involvement of TACE/ADAM17 and ADAM10 in etoposide-induced apoptosis of germ cells in rat spermatogenesis. J. Cell Physiol. 2012, 227, 829–838. [Google Scholar] [CrossRef]
- Codelia, V.A.; Cisterna, M.; Alvarez, A.R.; Moreno, R.D. p73 participates in male germ cells apoptosis induced by etoposide. Mol. Hum. Reprod. 2010, 16, 734–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, P.; Verhaar, A.P.; Peppelenbosch, M.P. Signaling Size: Ankyrin and SOCS Box-Containing ASB E3 Ligases in Action. Trends Biochem. Sci. 2019, 44, 64–74. [Google Scholar] [CrossRef]
- Guo, J.H.; Saiyin, H.; Wei, Y.H.; Chen, S.; Chen, L.; Bi, G.; Ma, L.J.; Zhou, G.J.; Huang, C.Q.; Yu, L.; et al. Expression of testis specific ankyrin repeat and SOCS box-containing 17 gene. Arch. Androl. 2004, 50, 155–161. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.S.; Kim, M.S.; Kim, S.K.; Baek, K.H. Murine Asb-17 expression during mouse testis development and spermatogenesis. Zygote 2004, 12, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Kohroki, J.; Nishiyama, T.; Nakamura, T.; Masuho, Y. ASB proteins interact with Cullin5 and Rbx2 to form E3 ubiquitin ligase complexes. FEBS Lett. 2005, 579, 6796–6802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uematsu, K.; Okumura, F.; Tonogai, S.; Joo-Okumura, A.; Alemayehu, D.H.; Nishikimi, A.; Fukui, Y.; Nakatsukasa, K.; Kamura, T. ASB7 regulates spindle dynamics and genome integrity by targeting DDA3 for proteasomal degradation. J. Cell Biol. 2016, 215, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.; Kim, D.; Rhee, J.W.; Park, J.A.; Kim, D.W.; Kim, D.S.; Lee, Y.; Kwon, H.J. ASB9 interacts with ubiquitous mitochondrial creatine kinase and inhibits mitochondrial function. BMC. Biol. 2010, 8, 1–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.Y.; Huang, M.Y.; Lin, Y.M.; Ho, C.H.; Lin, S.Y.; Chen, H.Y.; Hung, M.C.; Chen, R.H. BIK ubiquitination by the E3 ligase Cul5-ASB11 determines cell fate during cellular stress. J. Cell Biol. 2019, 218, 3002–3018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, J.; Grow, E.J.; Mlcochova, H.; Maher, G.J.; Lindskog, C.; Nie, X.; Guo, Y.; Takei, Y.; Yun, J.; Cai, L.; et al. The adult human testis transcriptional cell atlas. Cell Res. 2018, 28, 1141–1157. [Google Scholar] [CrossRef]
- Ogura, K.; Yahiro, K.; Tsutsuki, H.; Nagasawa, S.; Yamasaki, S.; Moss, J.; Noda, M. Characterization of Cholix toxin-induced apoptosis in HeLa cells. J. Biol. Chem. 2011, 286, 37207–37215. [Google Scholar] [CrossRef] [Green Version]
- Sixt, B.S.; Nunez-Otero, C.; Kepp, O.; Valdivia, R.H.; Kroemer, G. Chlamydia trachomatis fails to protect its growth niche against pro-apoptotic insults. Cell Death Differ. 2019, 26, 1485–1500. [Google Scholar] [CrossRef] [Green Version]
- Torriglia, A.; Negri, C.; Chaudun, E.; Prosperi, E.; Courtois, Y.; Counis, M.F.; Scovassi, A.I. Differential involvement of DNases in HeLa cell apoptosis induced by etoposide and long term-culture. Cell Death Differ. 1999, 6, 234–244. [Google Scholar] [CrossRef]
- Harper, N.; Farrow, S.N.; Kaptein, A.; Cohen, G.M.; MacFarlane, M. Modulation of tumor necrosis factor apoptosis-inducing ligand-induced NF-kappa B activation by inhibition of apical caspases. J. Biol. Chem. 2001, 276, 34743–34752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kale, J.; Osterlund, E.J.; Andrews, D.W. BCL-2 family proteins: Changing partners in the dance towards death. Cell Death Differ. 2018, 25, 65–80. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Zhang, W.; Xiao, Z.; Gan, H.; Lin, X.; Liao, S.; Han, C. Mining and characterization of ubiquitin E3 ligases expressed in the mouse testis. BMC Genom. 2012, 13, 495. [Google Scholar] [CrossRef] [Green Version]
- Richburg, J.H.; Myers, J.L.; Bratton, S.B. The role of E3 ligases in the ubiquitin-dependent regulation of spermatogenesis. Semin. Cell Dev. Biol. 2014, 30, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.R.; Dong, H.S.; Yang, W.X. Regulators in the apoptotic pathway during spermatogenesis: Killers or guards? Gene 2016, 582, 97–111. [Google Scholar] [CrossRef] [PubMed]
- Print, C.G.; Loveland, K.L.; Gibson, L.; Meehan, T.; Stylianou, A.; Wreford, N.; de Kretser, D.; Metcalf, D.; Kontgen, F.; Adams, J.M.; et al. Apoptosis regulator bcl-w is essential for spermatogenesis but appears otherwise redundant. Proc. Natl. Acad. Sci. USA 1998, 95, 12424–12431. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Wang, Y.; Ogawa, O.; Lee, H.G.; Raina, A.K.; Siedlak, S.L.; Harris, P.L.; Fujioka, H.; Shimohama, S.; Tabaton, M.; et al. Neuroprotective properties of Bcl-w in Alzheimer disease. J. Neurochem. 2004, 89, 1233–1240. [Google Scholar] [CrossRef] [PubMed]
- Hartman, M.L.; Czyz, M. BCL-w: Apoptotic and non-apoptotic role in health and disease. Cell Death Dis. 2020, 11, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perciavalle, R.M.; Opferman, J.T. Delving deeper: MCL-1′s contributions to normal and cancer biology. Trends Cell Biol. 2013, 23, 22–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Luo, Q.; Liu, Z. Ubiquitination and deubiquitination of MCL1 in cancer: Deciphering chemoresistance mechanisms and providing potential therapeutic options. Cell Death Dis. 2020, 11, 556. [Google Scholar] [CrossRef]
- Inuzuka, H.; Shaik, S.; Onoyama, I.; Gao, D.; Tseng, A.; Maser, R.S.; Zhai, B.; Wan, L.; Gutierrez, A.; Lau, A.W.; et al. SCF(FBW7) regulates cellular apoptosis by targeting MCL1 for ubiquitylation and destruction. Nature 2011, 471, 104–109. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, G.; Wan, P.; Xiang, Q.; Huang, S.; Huang, S.; Wang, J.; Wu, K.; Wu, J. E3 Ubiquitin Ligase ASB17 Promotes Apoptosis by Ubiquitylating and Degrading BCLW and MCL1. Biology 2021, 10, 234. https://doi.org/10.3390/biology10030234
Yang G, Wan P, Xiang Q, Huang S, Huang S, Wang J, Wu K, Wu J. E3 Ubiquitin Ligase ASB17 Promotes Apoptosis by Ubiquitylating and Degrading BCLW and MCL1. Biology. 2021; 10(3):234. https://doi.org/10.3390/biology10030234
Chicago/Turabian StyleYang, Ge, Pin Wan, Qi Xiang, Shanyu Huang, Siyu Huang, Jun Wang, Kailang Wu, and Jianguo Wu. 2021. "E3 Ubiquitin Ligase ASB17 Promotes Apoptosis by Ubiquitylating and Degrading BCLW and MCL1" Biology 10, no. 3: 234. https://doi.org/10.3390/biology10030234
APA StyleYang, G., Wan, P., Xiang, Q., Huang, S., Huang, S., Wang, J., Wu, K., & Wu, J. (2021). E3 Ubiquitin Ligase ASB17 Promotes Apoptosis by Ubiquitylating and Degrading BCLW and MCL1. Biology, 10(3), 234. https://doi.org/10.3390/biology10030234