
Emergence of Colistin and Carbapenem Resistance in Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae Isolated from Chickens and Humans in Egypt

 , ,
, ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Sampling
2.2. Isolation and Identification of K. pneumoniae
2.3. Antimicrobial Susceptibility Testing
2.4. Phenotypic Screening for ESBLs and Carbapenemases Production
2.5. Detection of ESBL, Carbapenemase-Encoding Genes, and Colistin Resistance Determinant mcr-1
2.6. ERIC Genotyping
2.7. Data Analysis
3. Results
3.1. Prevalence of K. pneumoniae in Chickens, Their Environment, Contact Workers and Hospitalized Patients in the Study Area
3.2. Antimicrobial Resistance of K. pneumoniae Isolates
3.3. Distributions of ESBLs, Carbapenemase Genes and Colistin Resistance Determinant mcr-1 among K. pneumoniae Isolates
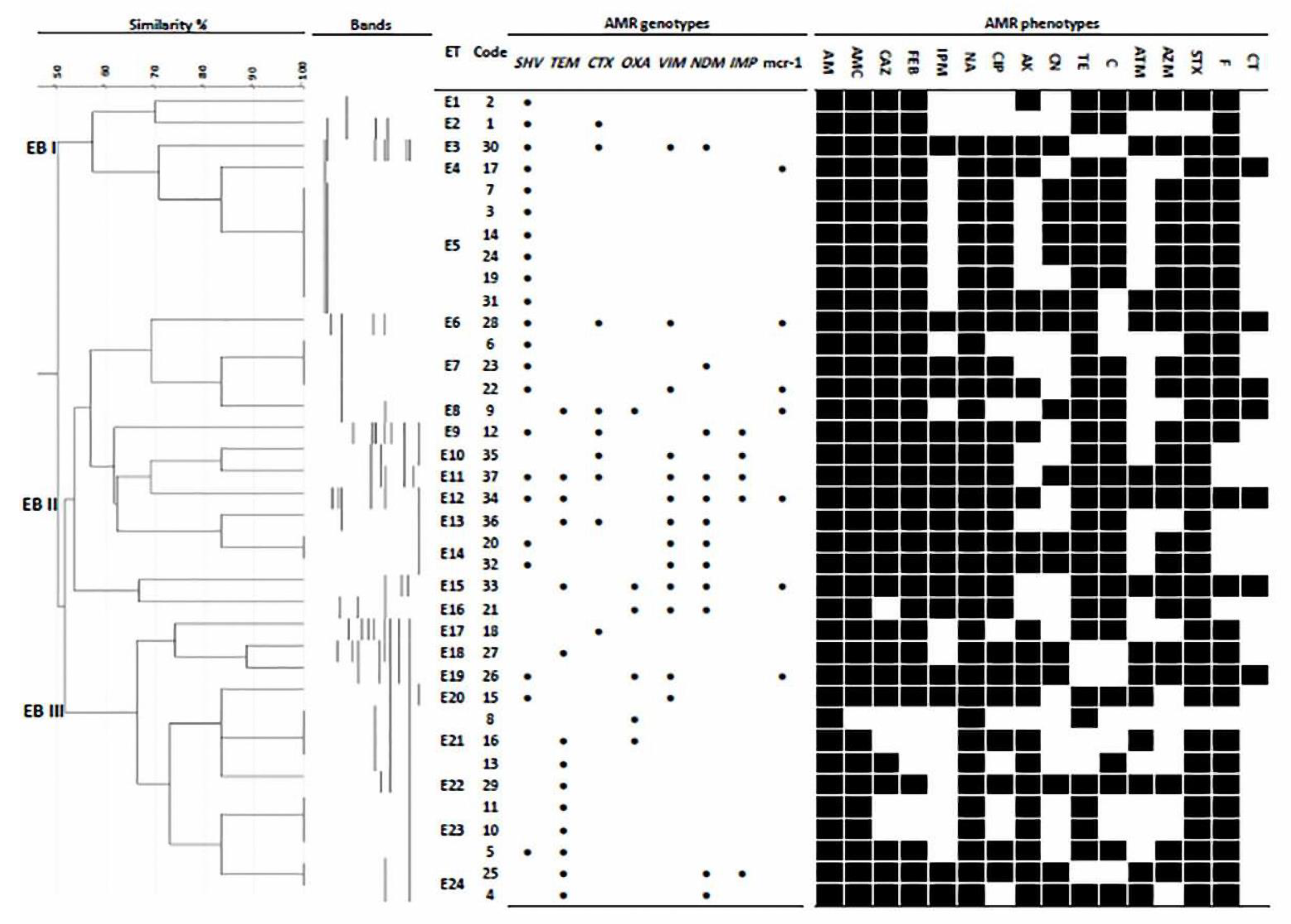
3.4. Genotyping and Epidemiological Association of Chicken and Human Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Cheng, F.; Li, Z.; Lan, S.; Liu, W.; Li, X.; Zhou, Z.; Song, Z.; Wu, J.; Zhang, M.; Shan, W. Characterization of Klebsiella pneumoniae associated with cattle infections in southwest China using multi-locus sequence typing (MLST), antibiotic resistance and virulence-associated gene profile analysis. Braz. J. Microbiol. 2018, 49, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Ripabelli, G.; Tamburro, M.; Guerrizio, G.; Fanelli, I.; Flocco, R.; Scutellà, M.; Sammarco, M.L. Tracking multidrug-resistant Klebsiella pneumoniae from an Italian hospital: Molecular epidemiology and surveillance by PFGE, RAPD and PCR-Based resistance genes prevalence. Curr. Microbiol. 2018, 75, 977–987. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.K.; Gharib, A.A. Coexistence of plasmid-mediated quinolone resistance determinants and AmpC-beta-lactamases in Escherichia coli strains in Egypt. Cell. Mol. Biol. 2015, 61, 29–35. [Google Scholar] [CrossRef]
- Dahshan, H.; Abd-Elall, A.M.M.; Megahed, A.M.; Abd-El-Kader, M.A.; Nabawy, E.E. Veterinary antibiotic resistance, residues, and ecological risks in environmental samples obtained from poultry farms, Egypt. Environ. Monit. Assess. 2015, 187, 2. [Google Scholar] [CrossRef]
- WHO. Report on the Consultative Meeting on Antimicrobial Resistance for Countries in the Eastern Mediterranean Region: From Policies to Action Sharm el Sheikh, Egypt, 12–14 November 2013. Available online: http://apps.who.int/iris/handle/10665/116211?show=full (accessed on 28 April 2020).
- Overdevest, I.T.M.A.; Heck, M.; van der Zwaluw, K.; Huijsdens, X.; van Santen, M.; Rijnsburger, M.; Eustace, A.; Xu, L.; Hawkey, P.; Savelkou, P.; et al. Extended-spectrum β-lactamase producing Klebsiella spp. in chicken meat and humans: A comparison of typing methods. Clin. Microbiol. Infect. 2014, 20, 251–255. [Google Scholar] [CrossRef] [Green Version]
- Elmowalid, G.A.; Ahmed, A.M.; Hassan, M.N.; Abd El-Aziz, N.K.; Abdelwahab, A.M.; Elwan, S.I. Molecular Detection of New SHV β-lactamase variants in clinical Escherichia coli and Klebsiella pneumoniae isolates from Egypt. Comp. Immunol. Microbiol. Infect. Dis. 2018, 60, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Alizadeh, N.; Rezaee, M.A.; Kafil, H.S.; Barhaghi, M.H.S.; Memar, M.Y.; Milani, M.; Hasani, A.; Ghotaslou, R. Detection of carbapenem-resistant Enterobacteriaceae by chromogenic screening media. J. Microbiol. Methods 2018, 153, 40–44. [Google Scholar] [CrossRef]
- Petrosillo, N.; Taglietti, F.; Granata, G. Treatment options for colistin resistant Klebsiella pneumoniae: Present and Future. J. Clin. Med. 2019, 8, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magiorakos, A.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; OlssonLiljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, G.; Bonomo, R.A. “Stormy waters ahead”: Global emergence of carbapenemases. Front. Microbiol. 2013, 4, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skov, R.L.; Monnet, D.L. Plasmid-mediated colistin resistance (mcr-1 gene): Three months later, the story unfolds. Eurosurveillance 2016, 21, 30155. [Google Scholar] [CrossRef] [Green Version]
- Durante-Mangoni, E.; Andini, R.; Zampino, R. Management of carbapenem-resistant Enterobacteriaceae infections. Clin. Microbiol. Infect. 2019, 25, 943–950. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Bush, K. Carbapenemases: The versatile β-lactamases. Clin. Microbiol. Rev. 2007, 20, 440–458. [Google Scholar] [CrossRef] [Green Version]
- Silva, N.; Carvalho, I.; Currie, C.; Sousa, M.; Igrejas, G.; Poeta, P. Extended-Spectrum-β-Lactamase and carbapenemase-producing Enterobacteriaceae in food-producing animals in Europe. In Antibiotic Drug Resistance; Igrejas, J.C.G., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; pp. 261–273. [Google Scholar]
- Zafer, M.M.; El-Mahallawy, H.A.; Abdulhak, A.; Amin, M.A.; Al-Agamy, M.H.; Radwan, H.H. Emergence of colistin resistance in multidrug-resistant Klebsiella pneumoniae and Escherichia coli strains isolated from cancer patients. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 40. [Google Scholar] [CrossRef] [PubMed]
- El-Badawy, M.F.; El-Far, S.W.; Althobaiti, S.S.; Abou-Elazm, F.I.; Shohayeb, M.M. The first Egyptian report showing the co-existence of blaNDM-25, blaOXA-23, blaOXA-181, and blaGES-1 among carbapenem-resistant K. Pneumoniae clinical isolates genotyped by BOX-PCR. Infect. Drug Resist. 2020, 13, 1237–1250. [Google Scholar] [CrossRef] [PubMed]
- Abdel Salam, S.A.; Hager, R. Colistin susceptibility among multidrug resistant Gram-negative bacilli isolated from Tertiary hospital in Egypt. Nov. Res. Microbiol. J. 2020, 4, 968–978. [Google Scholar] [CrossRef]
- Wilson, L.A.; Sharp, P.M. Enterobacterial repetitive intergenic consensus (ERIC) sequences in Escherichia coli: Evolution and implications for ERIC-PCR. Mol. Biol. Evol. 2006, 23, 1156–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tartor, Y.H.; EL-Naenaeey, E.-S.Y.; Abdallah, H.M.; Samir, M.; Yassen, M.M.; Abdelwahab, A.M. Virulotyping and genetic diversity of Aeromonas hydrophila isolated from Nile tilapia (Oreochromis niloticus) in aquaculture farms in Egypt. Aquaculture 2021, 541, 736781. [Google Scholar] [CrossRef]
- Markey, B.; Leonard, F.; Archambault, M.; Cullinane, A.; Maguire, D. Clinical Veterinary Microbiology, 2nd ed.; Mosby: London, UK, 2013. [Google Scholar]
- Pomakova, D.K.; Hsiao, C.B.; Beanan, J.M.; Olson, R.; MacDonald, U.; Keynan, Y.; Russo, T.A. Clinical and phenotypic differences between classic and hypervirulent Klebsiella pneumonia: An emerging and under-recognized pathogenic variant. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 31, 981–989. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, C.; Zheng, W.; Zhang, X.; Yu, J.; Gao, Q.; Hou, Y.; Huang, X. PCR detection of Klebsiella pneumoniae in infant formula based on 16S-23S internal transcribed spacer. Int. J. Food Microbiol. 2008, 125, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Siu, G.K.H.; Yeung, A.S.F.; Chen, J.H.K.; Ho, P.L.; Leung, K.W.; Tsang, J.L.Y.; Cheng, V.C.C.; Guo, L.; Yang, J.; et al. Performance of the VITEK MS matrix-assisted laser desorption ionization-time of flight mass spectrometry system for rapid bacterial identification in two diagnostic centres in China. J. Med. Microbiol. 2015, 64, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Bauer, A.W.; Kirby, W.M.; Sherris, J.C.; Truck, M. Antibiotic susceptibility testing by a standardized single disk method. Am. J. Clin. Pathol. 1966, 45, 493–496. [Google Scholar] [CrossRef]
- Plumb, D. Plumb’s Veterinary Drug Handbook, 8th ed.; (Pocket); Wiley-Blackwell: Ames, IA, USA, 2015; ISBN 9781118911921. [Google Scholar]
- Rankin, D. Test Methods: MIC Testing. In Manual of Antimicrobial Susceptibility Testing; Coyle, B.M., Ed.; American Society for Microbiology: Washington, DC, USA, 2005; pp. 53–62. [Google Scholar]
- EUCAST. Recommendations for MIC Determination of Colistin (Polymyxin E) as Recommended by the Joint CLSI-EUCAST Polymyxin Breakpoints Working Group; European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2016; Available online: http://www.Eucast.Org (accessed on 20 March 2020).
- Tambekar, D.; Dhanorkar, D.; Gulhane, S.; Khandelwal, V.; Dudhane, M. Antibacterial susceptibility of some urinary tract pathogens to commonly used antibiotics. Afr. J. Biotechnol. 2006, 5, 1562–1565. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing. Twenty Second Informational Supplement Update; CLSI Document M100-S22 U; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fourth Informational Supplement; CLSI Document M100-S24; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Pournaras, S.; Poulou, A.; Tsakris, A. Inhibitor-based methods for the detection of KPC carbapenemase-producing Enterobacteriaceae in clinical practice by using boronic acid compounds. J. Antimicrob. Chemother. 2010, 65, 1319–1321. [Google Scholar] [CrossRef]
- Ogutu, J.O.; Zhang, Q.; Huang, Y.; Yan, H.; Su, L.; Gao, B.; Zhang, W.; Zhao, J.; Cai, W.; Li, W.; et al. Development of a multiplex PCR system and its application in detection of blaSHV, blaTEM, blaCTX-M-1, blaCTX-M-9 and blaOXA-1 group genes in clinical Klebsiella pneumoniae and Escherichia coli strains. J. Antibiot. 2015, 68, 725–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, C.; Malczynski, M.; Parker, M.; Scheetz, M.H. Characterization of genetic diversity of carbapenem-resistant Acinetobacter baumannii clinical strains collected from 2004 to 2007. J. Clin. Microbiol. 2008, 46, 1106–1109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, Y.; Liang, Z.; Su, X.; Xiong, Y. Characterization of carbapenemase genes in Enterobacteriaceae species exhibiting decreased susceptibility to carbapenems in a university hospital in Chongqing, China. Ann. Lab. Med. 2012, 32, 270–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-β-lactamase gene, blaNDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.Y.; Wang, Y.; Walsh, T.R.; Yi, L.X.; Zhang, R.; Spencer, J.; Doi, Y.; Tian, G.; Dong, B.; Huang, X.; et al. Emergence of plasmid-mediated colistin resistance mechanism MCR-1 in animals and human beings in China: A microbiological and molecular biological study. Lancet Infect. Dis. 2016, 16, 161–168. [Google Scholar] [CrossRef]
- Versalovic, J.; Koeuth, T.; Lupski, R. Distribution of repetitive DNA sequences in eubacteria and application to finerpriting of bacterial enomes. Nucleic Acids Res. 1991, 19, 6823–6831. [Google Scholar] [CrossRef]
- Heras, J.; Domínguez, C.; Mata, E.; Pascual, V.; Lozano, C.; Torres, C.; Zarazaga, M. GelJ—A tool for analyzing DNA fingerprint gel images. BMC Bioinformatics 2015, 16, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, P.R.; Gaston, M.A. Numerical index of the discriminatory ability of typing systems: An application of Simpson’s index of diversity. J. Clin. Microbiol. 1988, 26, 2465–2466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempf, I.; Fleury, M.A.; Drider, D.; Bruneau, M.; Sanders, P.; Chauvin, C.; Madec, J.Y.; Jouy, E. What do we know about resistance to colistin in Enterobacteriaceae in avian and pig production in Europe? Int. J. Antimicrob. Agents 2013, 42, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Hamza, E.; Dorgham, S.M.; Hamza, D.A. Carbapenemase-producing Klebsiella pneumoniae in broiler poultry farming in Egypt. J. Glob. Antimicrob. Resist. 2016, 7, 8–10. [Google Scholar] [CrossRef]
- Elwakil, B.H.; Ali, S.M.; Hafez, S.F.; Bekhit, A.A.; El-Naggar, M.Y.; Olama, Z.A. Resistance prevalence profile of Klebsiella pneumoniae in the Intensive Care Units of Al-Shatby pediatric hospital, Alexandria, Egypt. Nov. Res. Microbiol. J. 2019, 3, 535–545. [Google Scholar] [CrossRef]
- Yang, F.; Deng, B.; Liao, W.; Wang, P.; Chen, P.; Wei, J. High rate of multiresistant Klebsiella pneumoniae from human and animal origin. Infect. Drug Resist. 2019, 12, 2729. [Google Scholar] [CrossRef] [Green Version]
- Juan, C.H.; Fang, S.Y.; Chou, C.H.; Tsai, T.Y.; Lin, Y.T. Clinical characteristics of patients with pneumonia caused by Klebsiella pneumoniae in Taiwan and prevalence of antimicrobial-resistant and hypervirulent strains: A retrospective study. Antimicrob. Resist. Infect. Control 2020, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, G.S.; Waits, K.; Nordstrom, L.; Weaver, B.; Aziz, M.; Gauld, L.; Grande, H.; Bigler, R.; Horwinski, J.; Porter, S.; et al. Intermingled Klebsiella pneumoniae populations between retail meats and human urinary tract infections. Clin. Infect. Dis. 2015, 61, 892–899. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.H.; Song, X.Y.; Ma, X.B.; Zhang, S.Y.; Zhang, J.Q. Molecular characterization of multidrug-resistant Klebsiella pneumoniae isolates. Braz. J. Microbiol. 2015, 46, 759–768. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Gao, H.; Liu, Y.; Jin, L.; Wang, R.; Wang, X.; Wang, Q.; Yin, Y.; Zhang, Y.; Wang, H. Co-existence of a novel plasmid-mediated efflux pump with colistin resistance gene mcr in one plasmid confers transferable multidrug resistance in Klebsiella pneumoniae. Emerg. Microbes Infect. 2020, 9, 1102–1113. [Google Scholar] [CrossRef] [PubMed]
- Kidd, T.J.; Mills, G.; Sá-Pessoa, J.; Dumigan, A.; Frank, C.G.; Insua, J.L.; Ingram, R.; Hobley, L.; Bengoechea, J.A. A Klebsiella pneumoniae antibiotic resistance mechanism that subdues host defences and promotes virulence. EMBO Mol. Med. 2017, 9, 430–447. [Google Scholar] [CrossRef]
- Hayati, M.; Indrawati, A.; Mayasari, N.L.P.I.; Istiyaningsih, I.; Atikah, N. Molecular detection of extended-spectrum β-lactamase-producing Klebsiella pneumoniae isolates of chicken origin from East Java, Indonesia. Vet. World 2019, 12, 578–583. [Google Scholar] [CrossRef] [Green Version]
- Nirwati, H.; Sinanjung, K.; Fahrunissa, F.; Wijaya, F.; Napitupulu, S.; Hati, V.P.; Hakim, M.S.; Meliala, A.; Aman, A.T.; Nuryastuti, T. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc. 2019, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Atterby, C.; Osbjer, K.; Tepper, V.; Rajala, E.; Hernandez, J.; Seng, S.; Holl, D.; Bonnedahl, J.; Börjesson, S.; Magnusson, U.; et al. Carriage of carbapenemase- and extended-spectrum cephalosporinase-producing Escherichia coli and Klebsiella pneumoniae in humans and livestock in rural Cambodia; gender and age differences and detection of blaOXA-48in humans. Zoonoses Public Health 2019, 66, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Elalamy, R.A.; Tartor, Y.H.; Ammar, A.M.; Eldesouky, I.E.; Esawy, A.E.I. Molecular characterization of extensively drug-resistant Pasteurella multocida isolated from apparently healthy and diseased chickens in Egypt. Pak. Vet. J. 2020, 40, 319–324. [Google Scholar] [CrossRef]
- Prestinaci, F.; Pezzotti, P.; Pantosti, A. Antimicrobial resistance: A global multifaceted phenomenon. Pathog. Glob. Health 2015, 109, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Cantón, R.; Akóva, M.; Carmeli, Y.; Giske, C.G.; Glupczynski, Y.; Gniadkowski, M.; Livermore, D.M.; Miriagou, V.; Naas, T.; Rossolini, G.M.; et al. Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 2012, 18, 413–431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paterson, D.L.; Bonomo, R.A. Extended-spectrum β-lactamases: A clinical update. Clin. Microbiol. Rev. 2005, 18, 657–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livermore, D.M.; Canton, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.M.; Arlet, G.; Ayala, J.; Coque, T.M.; Kern-Zdanowicz, I.; Luzzaro, F.; et al. CTX-M: Changing the face of ESBLs in Europe. J. Antimicrob. Chemother. 2007, 59, 165–174. [Google Scholar] [CrossRef] [Green Version]
- Karanika, S.; Karantanos, T.; Arvanitis, M.; Grigoras, C.; Mylonakis, E. Fecal colonization with extended-spectrum Beta-lactamase-producing Enterobacteriaceae and risk factors among healthy individuals: A systematic review and metaanalysis. Clin. Infect. Dis. 2016, 63, 310–318. [Google Scholar] [CrossRef] [Green Version]
- Daehre, K.; Projahn, M.; Friese, A.; Semmler, T.; Guenther, S.; Roesler, U.H. ESBL-producing Klebsiella pneumoniae in the broiler production chain and the first description of ST3128. Front. Microbiol. 2018, 9, 302. [Google Scholar] [CrossRef]
- Delarampour, A.; Ghalehnoo, Z.R.; Khademi, F.; Delarampour, M.; Vaez, H. Molecular detection of carbapenem-resistant genes in clinical isolates of Klebsiella pneumoniae. Ann. Ig. 2019, 31, 349–355. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Lee, C.R.; Lee, J.H.; Park, K.S.; Kim, Y.B.; Jeong, B.C.; Lee, S.H. Global dissemination of carbapenemase-producing Klebsiella pneumoniae: Epidemiology, genetic context, treatment options, and detection methods. Front. Microbiol. 2016, 7, 895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eftekhar, F.; Naseh, Z. Extended-spectrum beta-lactamase and carbapenemase production among burn and non-burn clinical isolates of Klebsiella pneumoniae. Iran. J. Microbiol. 2015, 7, 144–149. [Google Scholar] [PubMed]
- Caspar, Y.; Maillet, M.; Pavese, P.; Francony, G.; Brion, J.; Mallaret, M.; Bonnet, R. Colistin resistance in ESBL-producing Klebsiella pneumoniae, France. Emerg. Infect. Dis. 2017, 23, 874–876. [Google Scholar] [CrossRef] [PubMed]
- Haeili, M.; Feizabadi, M.M. The threat of colistin resistance among carbapenem-resistant Klebsiella pneumoniae isolates in Iran. Iran. J. Microbiol. 2018, 10, 72–73. [Google Scholar] [PubMed]
- Rojas, L.J.; Salim, M.; Cober, E.; Richter, S.S.; Perez, F.; Salata, R.A.; Kalayjian, R.C.; Watkins, R.R.; Marshall, S.; Rudin, S.D.; et al. Colistin resistance in carbapenem-resistant Klebsiella pneumoniae: Laboratory detection and impact on mortality. Clin. Infect. Dis. 2017, 64, 711–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Centre for Disease Prevention and Control. Outbreak of Carbapenemase-Producing (NDM-1 and OXA-48) and Colistin-Resistant Klebsiella pneumoniae ST307, North-East Germany, 2019; ECDC: Stockholm, Sweden, 2019. [Google Scholar]
- Erdem, F.; Abulaila, A.; Aktas, Z.; Oncul, O. In vitro evaluation of double carbapenem and colistin combinations against OXA-48, NDM carbapenemase-producing colistin-resistant Klebsiella pneumoniae strains. Antimicrob. Resist. Infect. Control 2020, 9, 70. [Google Scholar] [CrossRef]
- Sedighi, P.; Zarei, O.; Karimi, K.; Taheri, M.; Karami, P.; Shokoohizadeh, L. Molecular typing of Klebsiella pneumoniae Clinical Isolates by Enterobacterial Repetitive Intergenic Consensus Polymerase Chain Reaction. Int. J. Microbiol. 2020, 1–5. [Google Scholar] [CrossRef]
- Shaikh, S.; Rizvi, S.M.D.; Anis, R.; Shakil, S. Prevalence of CTX-M resistance marker and integrons among Escherichia coli and Klebsiella pneumoniae isolates of clinical origin. Lett. Appl. Microbiol. 2016, 62, 419–427. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Isolates (%) * | No. of Isolates (%) | Chicken vs. Human | ||||||
|---|---|---|---|---|---|---|---|---|
| Chickens | Humans | |||||||
| C (n = 100) | Ce (n = 60) | TC (n = 160) | Cw (n = 22) | Hc (n = 90) | TH (n = 112) | X2 | p-Value | |
| MDR-K. pneumoniae (100) | 9 (9) | 10 (16.7) | 19 (11.9) | 5 (22.7) | 13 (14.4) | 18 (16.1) | 0.9 | 0.3 |
| ESBL-K. pneumoniae (100) | 9 (9) | 10 (16.7) | 19 (11.9) | 5 (22.7) | 13 (14.4) | 18 (16.1) | 0.9 | 0.3 |
| CR-K. pneumoniae (45.9) | 1 (1) | 2 (3.3) | 3 (1.9) | 4 (18.2) | 10 (11.1) | 14 (12.5) | 12.7 | >0.001 |
| CTR-K. pneumoniae (18.9) | 1 (1) | 1 (1.7) | 2 (1.25) | 1 (4.5) | 4 (4.4) | 5 (4.5) | 2.7 | 0.1 |
| ESBL and CR-K. pneumoniae (45.9) | 1 (1) | 2 (3.3) | 3 (1.9) | 4 (18.2) | 10 (11.1) | 14 (12.5) | 12.7 | >0.001 |
| ESBL and CTR-K. pneumoniae (18.9) | 1(1) | 1 (1.7) | 2 (1.25) | 1 (4.5) | 4 (4.4) | 5 (4.5) | 2.7 | 0.1 |
| CR and CTR-K. pneumoniae (13.5) | 0 | 0 | 0 | 1 (4.5) | 4 (4.4) | 5 (4.5) | - | - |
| ESBL, CR and CTR-K. pneumoniae (13.5) | 0 | 0 | 0 | 1 (4.5) | 4 (4.4) | 5 (4.5) | - | - |
| Pattern No. | Source | Code No. | Antimicrobial Resistance Pattern | MAR Index | Colistin MIC (mg/L) | Genotype | ERIC Fingerprint |
|---|---|---|---|---|---|---|---|
| (Band Size) | |||||||
| 1 | Hc | 34 | Amp, AMC, CAZ, FEP, IPM, NA, CIP, AK, TE, C, ATM, AZM, SXT, F, CT | 0.94 | 16 | blaSHV, blaTEM, blaVIM, blaNDM-1, blaIMP, mcr-1 | E12 (109, 312, 423, 878, 1000, 1225) |
| 2 | Cw | 22 | Amp, AMC, CAZ, FEP, IPM, NA, CIP, AK, TE, C, AZM, SXT, F, CT | 0.88 | 8 | blaSHV, blaVIM, mcr-1 | E7 (872) |
| 3 | Cw | 23 | Amp, AMC, CAZ, FEP, NA, IPM, CIP, TE, C, AZM, SXT, F | 0.75 | 1 | blaSHV, blaNDM-1 | E7 (872) |
| 4 | C | 6 | Amp, AMC, CAZ, FEP, NA, TE, SXT, F | 0.5 | 0.25 | blaSHV | E7 (878) |
| 5 | Hc | 28 | Amp, AMC, CAZ, FEP, CN, IPM, NA, CIP, AK, TE, ATM, AZM, SXT, F, CT | 0.94 | 32 | blaSHV, blaCTX-M1, blaVIM, mcr-1 | E6 (298, 428, 860, 1148) |
| 6 | Hc | 29 | Amp, AMC, CAZ, FEP, CN, NA, CIP, AK, TE, C, ATM, AZM, SXT, F | 0.88 | 0.5 | blaTEM | E22 (157, 305, 401) |
| 7 | C | 4 | Amp, AMC, CAZ, FEP, CN, IPM, NA, AK, TE, C, ATM, SXT, F | 0.81 | 0.25 | blaTEM, blaNDM-1 | E24 (159, 412) |
| 8 | Hc | 25 | Amp, AMC, CAZ, FEP, CN, IPM, NA, CIP, AK, ATM, AZM, SXT, F | 0.81 | 1 | blaTEM, blaNDM-1, blaIMP | E24 (150, 380) |
| 9 | Ce | 12 | Amp, AMC, CAZ, FEP, IPM, NA, CIP, AK, TE, C, AZM, SXT, F | 0.81 | 0.5 | blaSHV, blaCTX-M1, blaNDM-1, blaIMP | E9 (127, 199, 294, 352, 436, 489, 748) |
| 10 | Ce | 15 | Amp, AMC, CAZ, FEP, IPM, NA, CIP, AK, TE, C, ATM, SXT, F | 0.81 | 0.25 | blaSHV, blaVIM | E20 (116, 168, 312) |
| 11 | Hc | 30 | Amp, AMC, CAZ, FEP, CN, IPM, NA, CIP, AK, TE, C, AZM, SXT | 0.81 | 2 | blaSHV, blaVIM, blaNDM-1 | E3 (144, 169, 320, 340, 432, 1205, 1346) E14 (107) |
| Cw | 20 | 1 | |||||
| 12 | Hc | 32 | Amp, AMC, CAZ, FEP, CN, IPM, NA, CIP, AK, ATM, AZM, SXT, F | 0.81 | 2 | blaSHV, blaCTX-M1, blaVIM, blaNDM-1 | E14 (117) |
| 13 | Hc | 26 | Amp, AMC, CAZ, FEP, CN, IPM, NA, CIP, AK, ATM, AZM, SXT, F, CT | 0.88 | 16 | blaSHV, blaOXA-1, blaVIM, mcr-1 | E19 (159, 244, 307, 429, 680) |
| 14 | Hc | 31 | Amp, AMC, CAZ, FEP, CN, NA, CIP, AK, ATM, TE, AZM, SXT, F | 0.81 | 1 | blaSHV | E5 (1187, 1307) |
| 15 | Ce | 19 | Amp, AMC, CAZ, FEP, NA, TE, CIP, C, AZM, SXT, F | 0.69 | 2 | blaSHV | E5 (1205, 1353) |
| 16 | C | 7 | Amp, AMC, CAZ, FEP, CN, NA, CIP, TE, C, AZM, SXT, F | 0.75 | 2 | blaSHV | E5 (1196, 1320) E5 (1214, 1353) E5 (1200,1333) E5 (1214, 1353) |
| C | 3 | 2 | |||||
| Ce | 14 | 1 | |||||
| Cw | 24 | 2 | |||||
| 17 | C | 5 | Amp, AMC, CAZ, FEP, NA, CIP, AK, TE, C, AZM, SXT, F | 0.75 | 0.5 | blaSHV, blaTEM | E23 (157) |
| 18 | Ce | 10 | Amp, AMC, NA, AK, TE, SXT, F | 0.44 | 0.5 | blaTEM | E23 (159) |
| Ce | 11 | 1 | |||||
| 19 | Hc | 37 | Amp, AMC, CAZ, FEP, CN, IPM, NA, CIP, ATM, TE, C, AZM, SXT | 0.81 | 1 | blaSHV, blaTEM, blaCTX-M1, blaVIM, blaNDM-1, blaIMP | E11 (188, 306) |
| 20 | Ce | 17 | Amp, AMC, CAZ, FEP, NA, CIP, AK, TE, C, SXT, F, CT | 0.75 | 4 | blaSHV, mcr-1 | E4 (1360) |
| 21 | Hc | 27 | Amp, AMC, CAZ, FEP, NA, AK, CN, CIP, ATM, AZM, SXT, F | 0.75 | 2 | blaTEM | E18 (160, 254, 301, 369, 424, 703, 884, 1197) |
| 22 | C | 2 | Amp, AMC, CAZ, FEP, TE, AK, C, ATM, AZM, SXT, F | 0.69 | 1 | blaSHV | E1 (679) |
| 23 | C | 9 | Amp, AMC, CAZ, FEP, NA, TE, C, CN, SXT, F, CT | 0.69 | 8 | blaTEM, blaCTX-M1, blaOXA-1, mcr-1 | E8 (891) |
| 24 | Hc | 35 | Amp, AMC, CAZ, IPM, FEP, NA, TE, CIP, C, AZM, SXT | 0.69 | 2 | blaCTX-M1, blaVIM, blaIMP | E10 (107, 188, 377, 492) |
| 25 | Ce | 18 | Amp, AMC, CAZ, FEP, NA, TE, AK, C, SXT, F | 0.63 | 1 | blaCTX-M1 | E17 (189, 266, 331, 373, 488, 565, 623, 964) |
| 26 | Hc | 33 | Amp, AMC, CAZ, FEP, NA, IPM, CIP, TE, C, ATM, AZM, SXT, F, CT | 0.88 | 16 | blaTEM, blaOXA-1, blaVIM, blaNDM-1, mcr-1 | E15 (167, 211, 316) |
| 27 | Cw | 21 | Amp, AMC, FEP, NA, IPM, CIP, TE, AZM, C, SXT | 0.63 | 0.25 | blaOXA-1, blaVIM, blaNDM-1 | E16 (396, 767, 1357) |
| 28 | Hc | 36 | Amp, AMC, CAZ, FEP, NA, IPM, CIP, TE, C, SXT | 0.63 | 2 | blaTEM, blaCTX-M1, blaVIM, blaNDM-1 | E13 (107, 878) |
| 29 | Ce | 13 | Amp, AMC, CAZ, NA, AK, C, SXT, F | 0.5 | 0.5 | blaTEM | E21(170, 320, 459) |
| 30 | Ce | 16 | Amp, AMC, CIP, NA, AK, ATM, SXT, F | 0.5 | 1 | blaTEM, blaOXA | E21(169, 320, 462) |
| 31 | C | 8 | AMP, NA, TE | 0.19 | 0.25 | blaOXA-1 | E21(170, 320, 466) |
| 32 | C | 1 | Amp, AMC, CAZ, FEB, TE, C, F | 0.44 | 1 | blaSHV, blaCTX-M1 | E2 (297, 409, 700, 1158) |
| No. | Isolates Group and Genotypes | Total | Chickens | Humans | ||||
|---|---|---|---|---|---|---|---|---|
| C | Ce | TC | Cw | Hc | TH | |||
| ESBL-producing isolates | ||||||||
| 1 | blaSHV | 8 (21.6) | 4 | 2 | 6 (75) | 1 | 1 | 2 (25) |
| 2 | blaSHV, blaCTX-M1 | 1 (2.7) | 1 | 1 (100) | ||||
| 3 | blaSHV, blaTEM | 1 (2.7) | 1 | 1 (100) | ||||
| 4 | blaTEM | 5 (13.5) | 3 | 3 (60) | 2 | 2 (40) | ||
| 5 | blaTEM, blaOXA-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 6 | blaCTX-M1 | 1 (2.7) | 1 | 1 (100) | ||||
| 7 | blaOXA-1 | 1 (2.7) | 1 | 1 (100) | ||||
| ESBL- and CR-producing isolates | ||||||||
| 8 | blaSHV, blaVIM | 1 (2.7) | 1 | 1 (100) | ||||
| 9 | blaSHV, blaNDM-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 10 | blaSHV, blaVIM, blaNDM-1 | 2 (5.4) | 1 | 1 | 2 (100) | |||
| 11 | blaSHV, blaCTX-M1, blaVIM, blaNDM-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 12 | blaSHV, blaCTX-M1, blaNDM-1, blaIMP | 1 (2.7) | 1 | 1 (100) | ||||
| 13 | blaSHV, blaTEM, blaCTX-M1, blaVIM, blaNDM-1, blaIMP | 1 (2.7) | 1 | |||||
| 14 | blaTEM, blaNDM-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 15 | blaTEM, blaNDM-1, blaIMP | 1 (2.7) | 1 | 1 (100) | ||||
| 16 | blaTEM, blaCTX-M1, blaVIM, blaNDM-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 17 | blaCTX-M1, blaVIM, blaIMP | 1 (2.7) | 1 | 1 (100) | ||||
| 18 | blaOXA, blaVIM, blaNDM-1 | 1 (2.7) | 1 | 1 (100) | ||||
| ESBL-producing and CTR isolates | ||||||||
| 19 | blaSHV, mcr-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 20 | blaTEM, blaCTX-M1, blaOXA-1, mcr-1 | 1 (2.7) | 1 | 1 (100) | ||||
| ESBL-, CR- producing and CTR isolates | ||||||||
| 21 | blaSHV, blaVIM, mcr-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 22 | blaSHV, blaCTX-M1, blaVIM, mcr-1 | 1 (2.7) | 1 | 1 (100) | ||||
| 23 | blaSHV, blaOXA-1, blaVIM, mcr-1 | 1 (2.7) | 1 | |||||
| 24 | blaSHV, blaTEM, blaVIM, blaNDM-1, blaIMP, mcr-1 | 1 (2.7) | 1 | |||||
| 25 | blaTEM, blaOXA-1, blaVIM, blaNDM-1, mcr-1 | 1 (2.7) | 1 | 1 (100) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elmonir, W.; Abd El-Aziz, N.K.; Tartor, Y.H.; Moustafa, S.M.; Abo Remela, E.M.; Eissa, R.; Saad, H.A.; Tawab, A.A. Emergence of Colistin and Carbapenem Resistance in Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae Isolated from Chickens and Humans in Egypt. Biology 2021, 10, 373. https://doi.org/10.3390/biology10050373
Elmonir W, Abd El-Aziz NK, Tartor YH, Moustafa SM, Abo Remela EM, Eissa R, Saad HA, Tawab AA. Emergence of Colistin and Carbapenem Resistance in Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae Isolated from Chickens and Humans in Egypt. Biology. 2021; 10(5):373. https://doi.org/10.3390/biology10050373
Chicago/Turabian StyleElmonir, Walid, Norhan K. Abd El-Aziz, Yasmine H. Tartor, Samar M. Moustafa, Etab M. Abo Remela, Radwa Eissa, Hosam A. Saad, and Ahmed Abdel Tawab. 2021. "Emergence of Colistin and Carbapenem Resistance in Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae Isolated from Chickens and Humans in Egypt" Biology 10, no. 5: 373. https://doi.org/10.3390/biology10050373
APA StyleElmonir, W., Abd El-Aziz, N. K., Tartor, Y. H., Moustafa, S. M., Abo Remela, E. M., Eissa, R., Saad, H. A., & Tawab, A. A. (2021). Emergence of Colistin and Carbapenem Resistance in Extended-Spectrum β-Lactamase Producing Klebsiella pneumoniae Isolated from Chickens and Humans in Egypt. Biology, 10(5), 373. https://doi.org/10.3390/biology10050373