
Enigmatic Evolutionary History of Porphobilinogen Deaminase in Eukaryotic Phototrophs


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
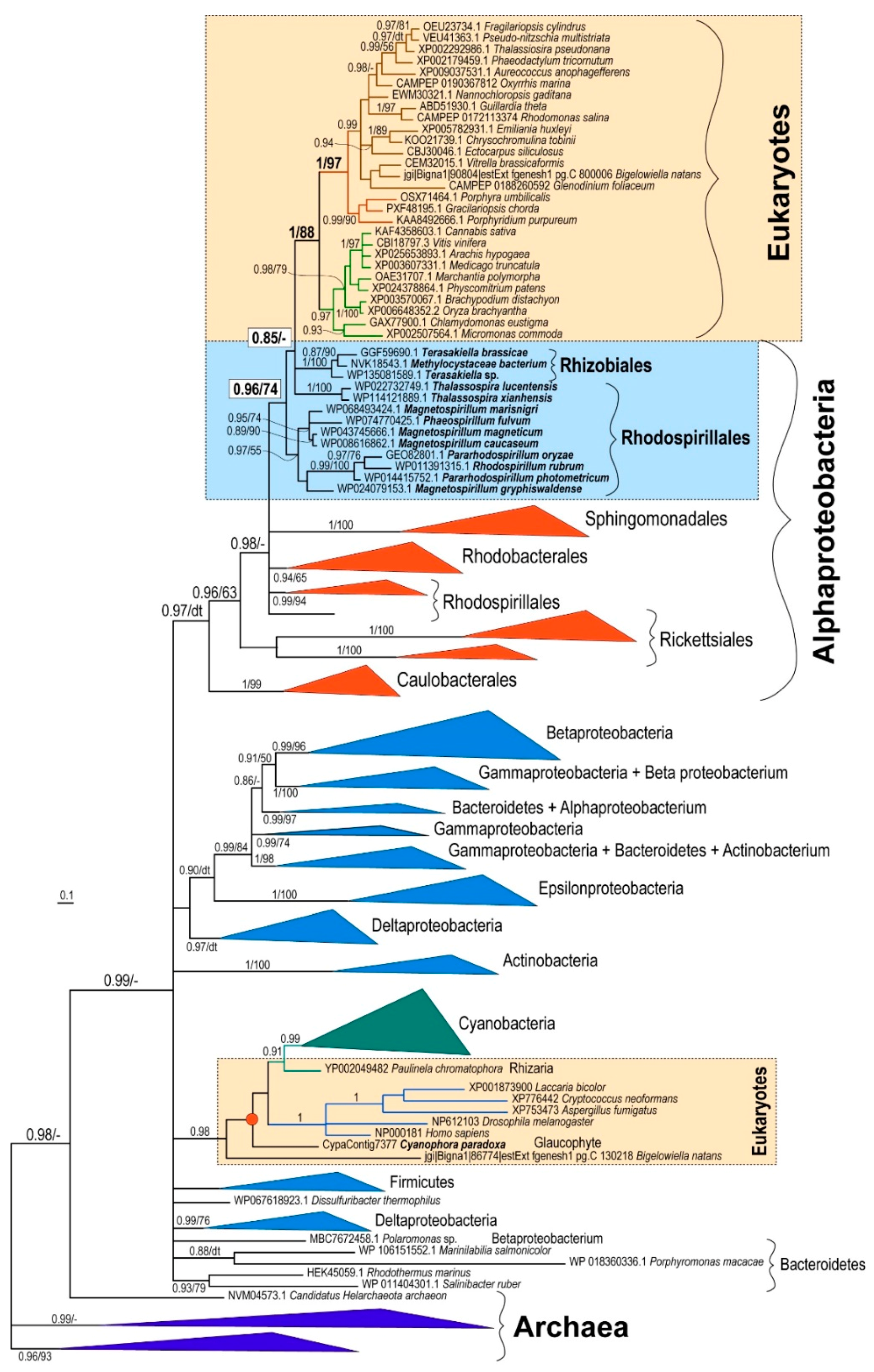
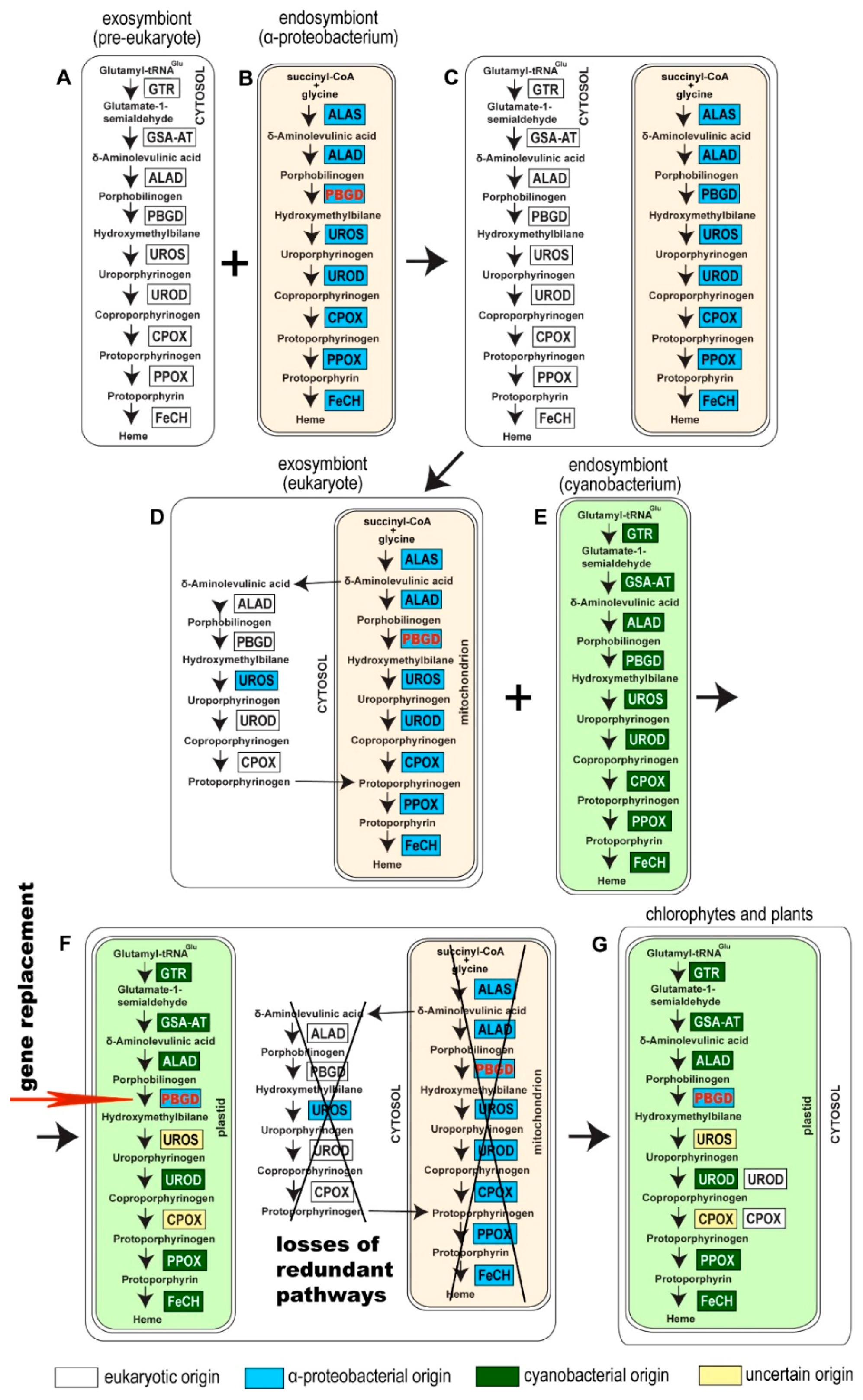
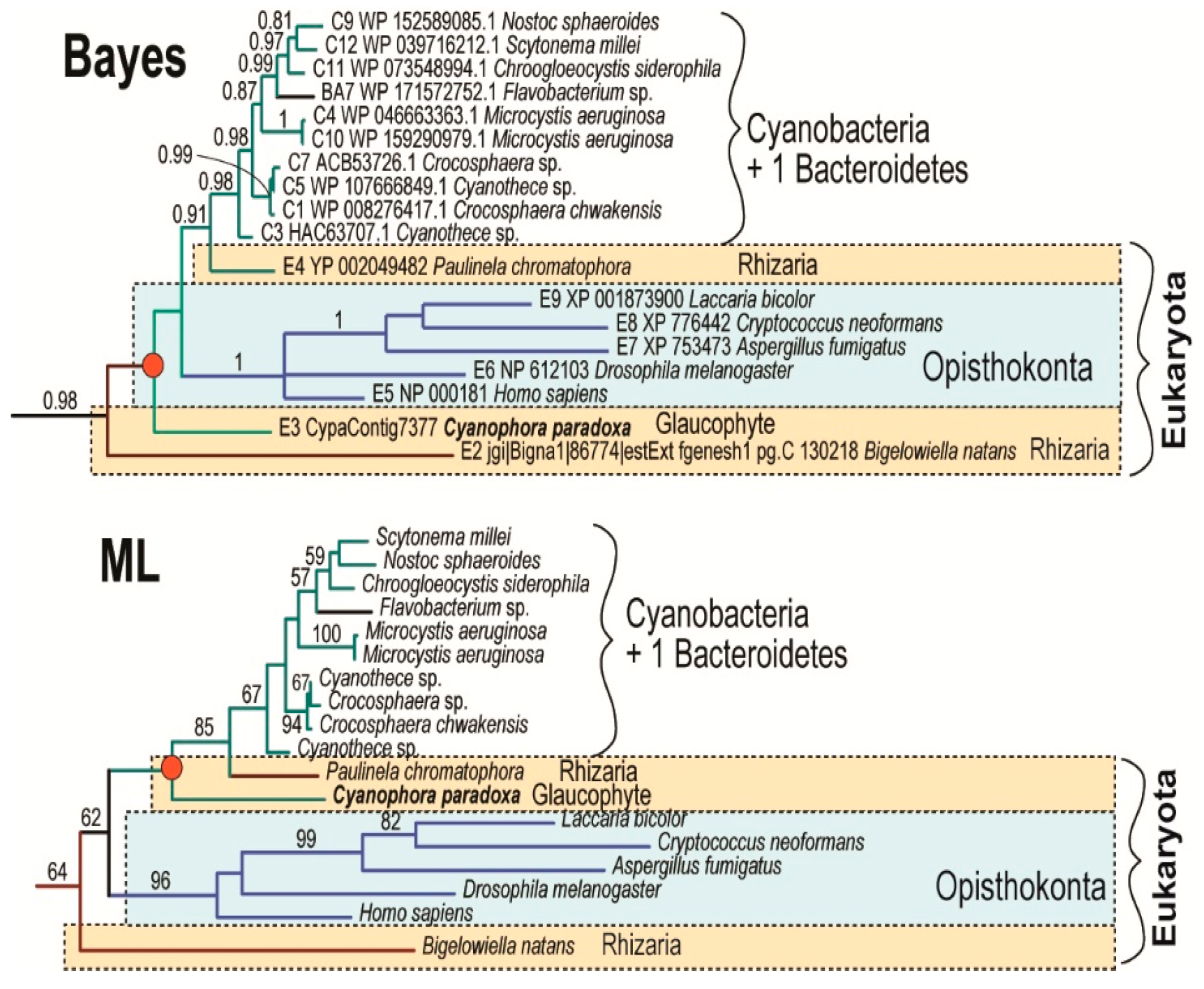
3. Results
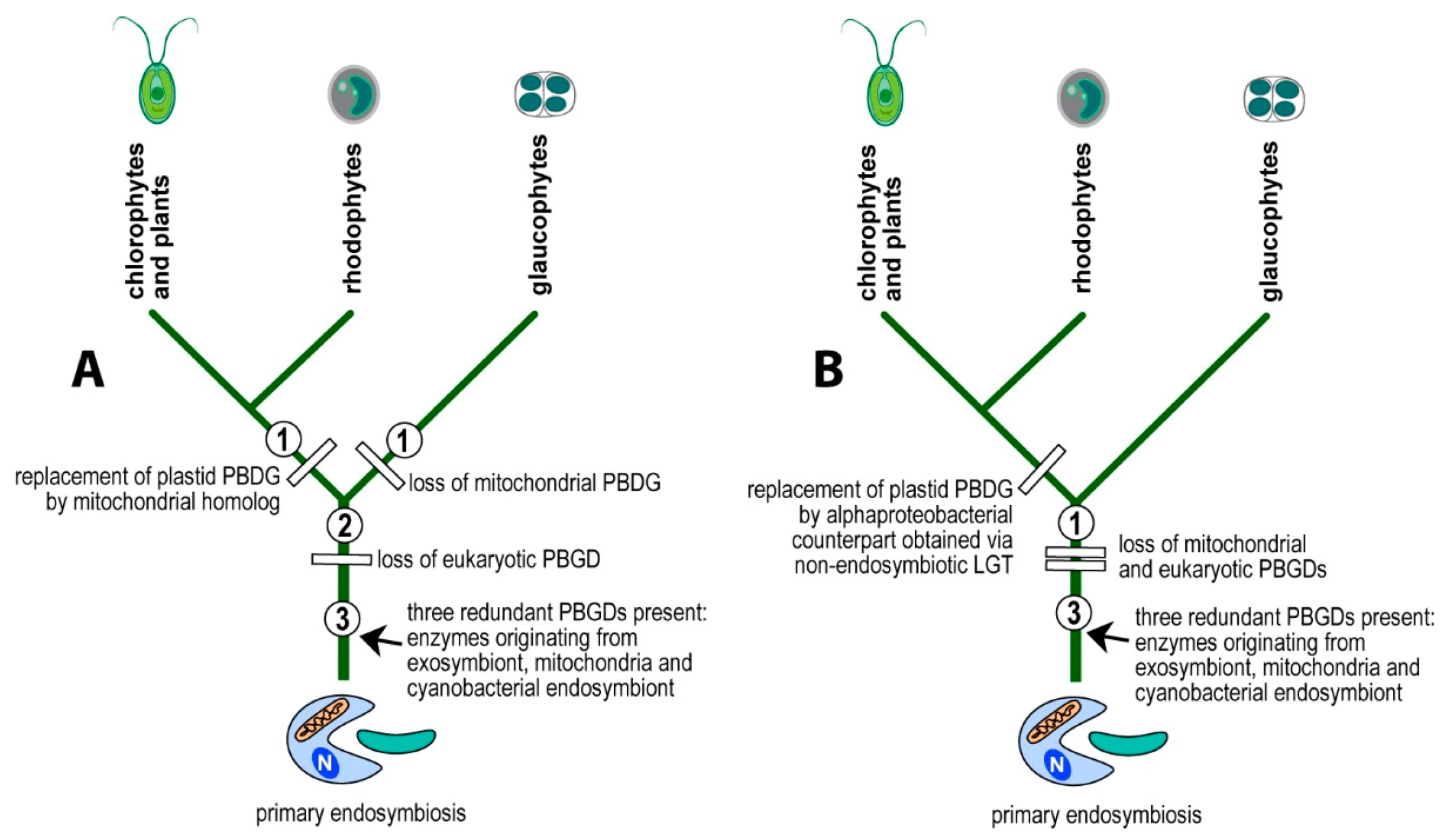
4. Discussion
5. Conclusions
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kořený, L.; Sobotka, R.; Kovářová, J.; Gnipová, A.; Flegontov, P.; Horváth, A.; Oborník, M.; Ayala, J.F.; Lukeš, J. Aerobic kinetoplastid flagellate Phytomonas does not require heme for viability. Proc. Natl. Acad. Sci. USA 2012, 109, 3808–3813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oborník, M.; Green, B.R. Mosaic origin of the heme biosynthesis pathway in photosynthetic eukaryotes. Mol. Biol. Evol. 2005, 22, 2343–2353. [Google Scholar] [CrossRef] [Green Version]
- Kořený, L.; Oborník, M. Sequence evidence for the presence of two tetrapyrrole pathways in Euglena gracilis. Genome Biol. Evol. 2011, 3, 359–364. [Google Scholar] [CrossRef] [Green Version]
- Cihlář, J.; Füssy, Z.; Horák, A.; Oborník, M. Evolution of the tetrapyrrole biosynthetic pathway in secondary algae: Conservation, redundancy and replacement. PLoS ONE 2016, 11, e0166338. [Google Scholar] [CrossRef]
- Cihlář, J.; Füssy, Z.; Oborník, M. Evolution of tetrapyrrole pathway in eukaryotic phototrophs. Adv. Bot. Res. 2018, 90, 273–309. [Google Scholar]
- Matsuo, E.; Inagaki, Y. Patterns in evolutionary origins of heme, chlorophyll a and isopentenyl diphosphate biosynthetic pathways suggest non-photosynthetic periods prior to plastid replacements in dinoflagellates. PeerJ 2018, 6, e5345. [Google Scholar] [CrossRef] [Green Version]
- Kořený, L.; Oborník, M.; Lukeš, J. Make it, take it, or leave it: Heme metabolism of parasites. PLoS Pathog. 2013, 9, e1003088. [Google Scholar] [CrossRef] [Green Version]
- Kořený, L.; Sobotka, R.; Janouškovec, J.; Keeling, P.J.; Oborník, M. Tetrapyrrole synthesis of photosynthetic chromerids is likely homologous to the unusual pathway of apicomplexan parasites. Plant Cell 2011, 23, 3454–3462. [Google Scholar] [CrossRef] [Green Version]
- Janouškovec, J.; Tikhonenkov, D.V.; Burki, F.; Howe, A.T.; Kolísko, M.; Mylnikov, A.P.; Keeling, P.J. Factors mediating plastid dependency and the origins of parasitism in apicomplexans and their close relatives. Proc. Natl. Acad. Sci. USA 2015, 112, 10200–10207. [Google Scholar] [CrossRef] [Green Version]
- Gawryluk, R.M.R.; Tikhonenkov, D.V.; Hehenberger, E.; Husník, F.; Mylnikov, A.P.; Keeling, P.J. Non-photosynthetic predators are sister to red algae. Nature 2019, 572, 240. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1793. [Google Scholar] [CrossRef] [Green Version]
- Gouy, M.; Guindon, S.; Gascuel, O. SeaView version 4: A multiplatform graphical user interface for sequence alignment and phylogenetic tree building. Mol. Biol. Evol. 2010, 27, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Keane, T.M.; Creevey, C.J.; Pentony, M.M.; Naughton, T.J.; McInerney, J.O. Assessment of methods for amino acid matrix selection and their use on empirical data shows that ad hoc assumptions for choice of matrix are not justified. BMC Evol. Biol. 2006, 6, 29. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML v8.2.X Manual; Heidelberg Institute of Teoretical Studies: Heidelberg, Germany, 2016. [Google Scholar]
- Lartillot, N.; Rodrigue, N.; Stubs, D.; Richer, J. PhyloBayes 3: A Bayesian software package for phylogenetic reconstruction and molecular dating. Syst. Biol. 2013, 62, 611–615. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Gómez, S.; Hess, S.; Burger, G.; Lang, B.F.; Susko, E.; Slamovits, C.H.; Roger, A.J. An updated phylogeny of the Alphaproteobacteria reveals that the parasitic Rickettsiales and Holosporales have independent origins. eLife 2019, 8, e42535. [Google Scholar] [CrossRef]
- Zaremba-Niedzwiedzka, K.; Caceres, E.F.; Saw, J.H.; Bäckström, D.; Juzokaite, L.; Vancaester, E.; Seitz, K.W.; Anantharaman, K.; Starnawski, P.; Kjelsden, K.U.; et al. Asgard archaea illuminate the origin of eukaryotic cellular complexity. Nature 2017, 541, 353–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Storbeck, S.; Rolfes, S.; Raux-Deery, E.; Warren, M.J.; Jahn, D.; Layer, G. A novel pathway for the biosynthesis of heme in Archaea: Genome-based bioinformatic predictions and experinmental evidence. Archaea 2010, 2010, 175050. [Google Scholar]
- Sato, T.; Haruyuki, A. Novel metabolic pathways in Archaea. Curr. Opin. Microbiol. 2011, 14, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Andersson, A.G.; Zomorodipour, A.; Andersson, J.O.; Sicheritz-Pontén, T.; Alsmark, U.C.; Powodski, R.M.; Näslund, A.K.; Eriksson, A.S.; Winkler, H.H.; Kurland, C.G. The genome of Rickettsia prowazekii and the origin of mitochondria. Nature 1998, 369, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, D.A.; Creevey, C.J.; McInerney, J.O. Genome phylogenies indicate a meaningful α-proteobacterial phylogeny and support a grouping of the mitochondria with the Rickettsiales. Mol. Biol. Evol. 2006, 23, 74–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, K.P.; Sobral, B.W.; Dickerman, A.W. A robust species tree for Alphaproteobacteria. J. Bacteriol. 2007, 189, 4578–4586. [Google Scholar] [CrossRef] [Green Version]
- Gray, M.W. Mitochondrial evolution. Cold Spring Harb. Persp. Biol. 2012, 4, a011403. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wu, M. An integrated phylogenomic approach toward pinpointing the origin of mitochondria. Sci. Rep. 2015, 5, 7949. [Google Scholar] [CrossRef] [Green Version]
- Roger, A.J.; Muñoz-Gómez, S.; Kamikawa, R. The origin and diversification of mitochondria. Curr. Biol. 2017, 27, R1177–R1192. [Google Scholar] [CrossRef] [Green Version]
- Atteia, A.; Adrait, A.; Brugière, S.; Tardif, M.; van Lis, R.; Deusch, O.; Dagan, T.; Kuth, L.; Gontero, B.; Martin, W.; et al. A proteomic survey of Chlamydomonas reinhardtii mitochondria sheds light on the metabolic plasticity of the organelle and on the nature of the α-proteobacterial mitochondrial ancestor. Mol. Biol. Evol. 2009, 26, 1533–1548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esser, C.; Ahmadinejad, N.; Wiegand, C.; Rotte, C.; Sebastiani, F.; Gelius-Dietrich, G.; Henze, K.; Kretschmann, E.; Richly, E.; Leiser, D.; et al. A genome phylogeny of mitochondria among α-proteobacteria and a predominantly eubacterial ancestry of yeasts nuclear genes. Mol. Biol. Evol. 2004, 21, 1643–1660. [Google Scholar] [CrossRef] [PubMed]
- Abhishek, A.; Bavishi, A.; Bavishi, A.; Choudhary, M. Bacterial genome chimaerism and the origin of mitochondria. Can. J. Microbiol. 2011, 57, 49–61. [Google Scholar] [CrossRef]
- Carvalho, D.S.; Andrade, R.F.S.; Pihno, S.T.R.; Góes-Neto, A.; Lobãlo, T.C.P.; Bomfim, G.C.; El-Hani, C.N. What are the evolutionary origins of mitochondria? A complex network approach. PLoS ONE 2015, 10, e0134988. [Google Scholar] [CrossRef]
- Martijn, J.; Vosseberg, J.; Guy, L.; Offre, P.; Etterna, T.J.G. Deep mitochondrial origin outside the sampled alphaproteobacterial. Nature 2018, 557, 101–105. [Google Scholar] [CrossRef]
- Dawkins, R. The Selfish Gene; Oxford University Press: Oxford, UK, 1976. [Google Scholar]
- Sobotka, R.; McLean, S.; Zuberova, M.; Hunter, C.N.; Tichy, M. The C-terminal extension of ferrochelatase is critical for enzyme activity and for functioning of the tetrapyrrole pathway in Synechocystis strain PCC 6803. J. Bacteriol. 2008, 190, 2086–2095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazderník, M.; Mareš, J.; Pilný, J.; Sobotka, R. The anthena-like domain of the cyanobacterial ferrochelatase can bind chlorophyll and carotenoids in an energy-dissipative configuration Binding of pigments on the ferrochelatase CAB domain. J. Biol. Chem. 2019, 294, 11131–11143. [Google Scholar] [CrossRef] [PubMed]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oborník, M. Enigmatic Evolutionary History of Porphobilinogen Deaminase in Eukaryotic Phototrophs. Biology 2021, 10, 386. https://doi.org/10.3390/biology10050386
Oborník M. Enigmatic Evolutionary History of Porphobilinogen Deaminase in Eukaryotic Phototrophs. Biology. 2021; 10(5):386. https://doi.org/10.3390/biology10050386
Chicago/Turabian StyleOborník, Miroslav. 2021. "Enigmatic Evolutionary History of Porphobilinogen Deaminase in Eukaryotic Phototrophs" Biology 10, no. 5: 386. https://doi.org/10.3390/biology10050386
APA StyleOborník, M. (2021). Enigmatic Evolutionary History of Porphobilinogen Deaminase in Eukaryotic Phototrophs. Biology, 10(5), 386. https://doi.org/10.3390/biology10050386