
Thermococcus bergensis sp. nov., a Novel Hyperthermophilic Starch-Degrading Archaeon

 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Isolation and Cultivation
2.2. Sequencing and Bioinformatics
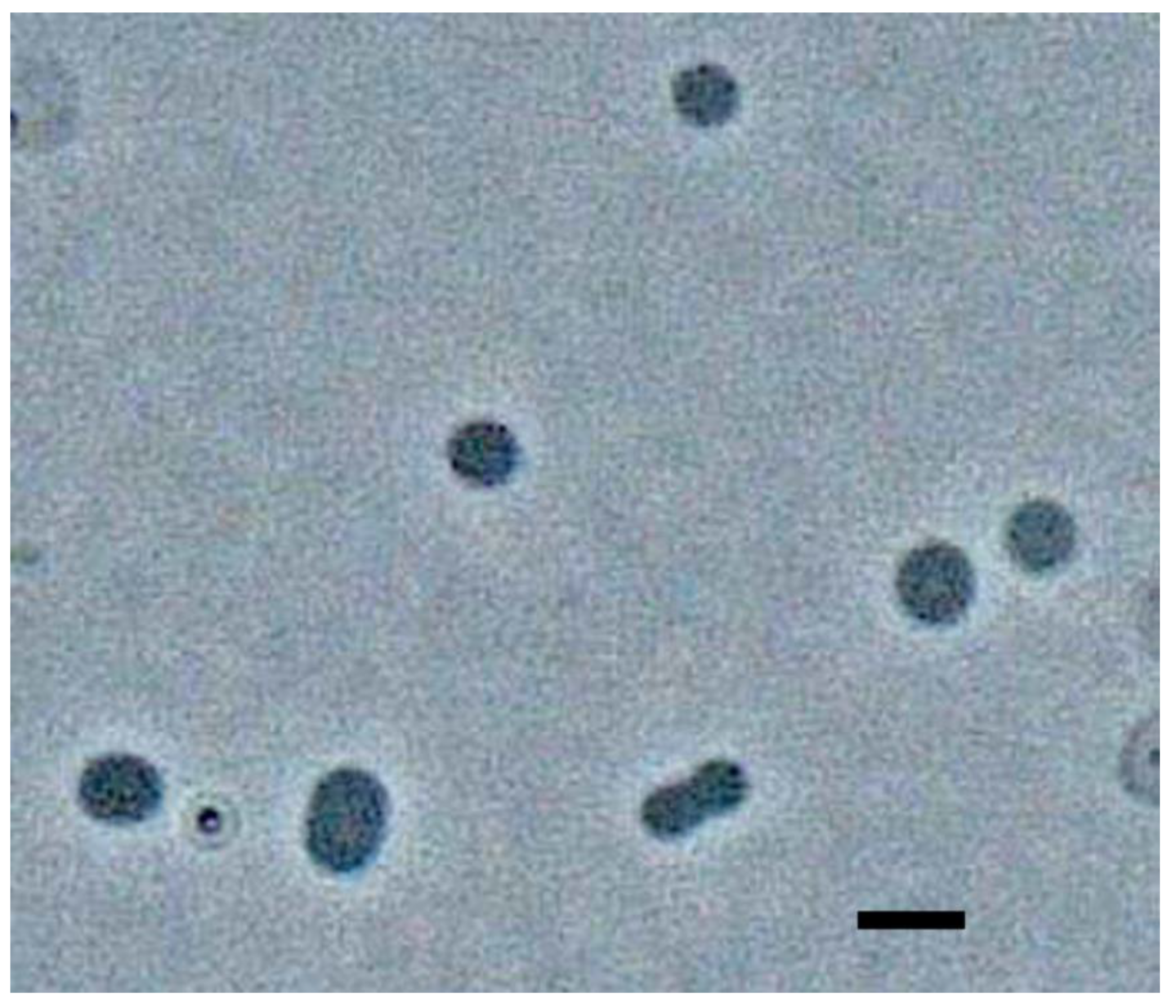
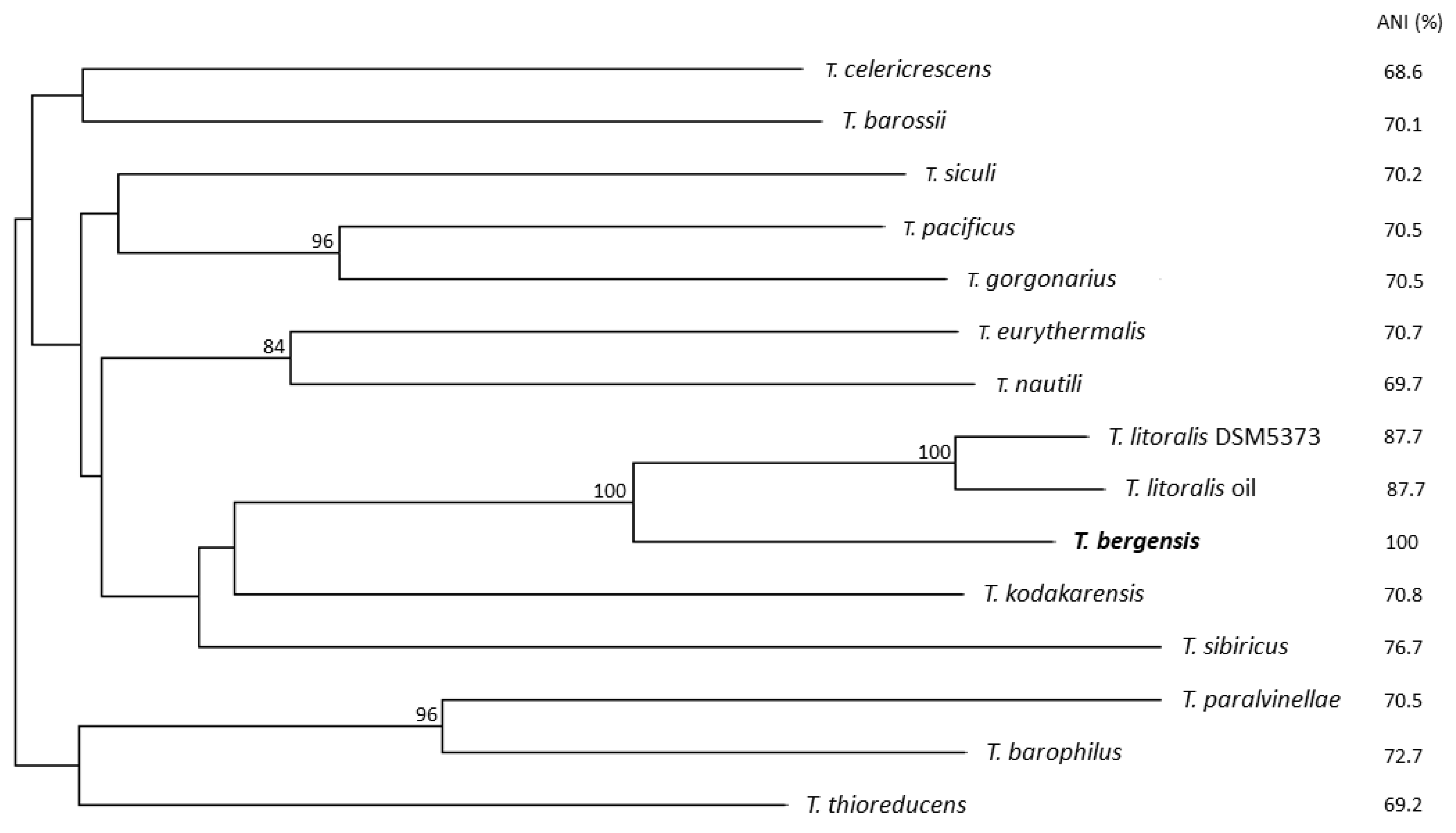
3. Results and Discussion
4. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Prieur, D. Hydrothermal Vents: Prokaryotes in Deep Sea Hydrothermal Vents. In Encyclopedia of Environmental Microbiology; Bitton, G., Ed.; John Wiley & Sons, Inc.: New York, NY, USA, 2003; pp. 1617–1628. [Google Scholar] [CrossRef]
- Zillig, W.; Holz, I.; Janekovic, D.; Schäfer, W.; Reiter, W.D. The Archaebacterium Thermococcus-Celer Represents, a Novel Genus within the Thermophilic Branch of the Archaebacteria. Syst. Appl. Microbiol. 1983, 4, 88–94. [Google Scholar] [CrossRef]
- Fiala, G.; Stetter, K.O. Pyrococcus furiosus sp. nov. represents a novel genus of marine heterotrophic archaebacteria growing optimally at 100 °C. Arch. Microbiol. 1986, 145, 56–61. [Google Scholar] [CrossRef]
- Takai, K.; Sugai, A.; Itoh, T.; Horikoshi, K. Palaeococcus ferrophilus gen. nov., sp. nov., a barophilic, hyperthermophilic archaeon from a deep-sea hydrothermal vent chimney. Int. J. Syst. Evol. Microbiol. 2000, 50, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Jolivet, E.; Corre, E.; L’Haridon, S.; Forterre, P.; Prieur, D. Thermococcus marinus sp. nov. and Thermococcus radiotolerans sp. nov., two hyperthermophilic archaea from deep-sea hydrothermal vents that resist ionizing radiation. Extremophiles 2004, 8, 219–227. [Google Scholar] [CrossRef]
- Gonzalez, J.M.; Sheckells, D.; Viebahn, M.; Krupatkina, D.; Borges, K.M.; Robb, F.T. Thermococcus waiotapuensis sp. nov., an extremely thermophilic archaeon isolated from a freshwater hot spring. Arch. Microbiol. 1999, 172, 95–101. [Google Scholar] [CrossRef]
- Ronimus, R.S.; Reysenbach, A.L.; Musgrave, D.R.; Morgan, H.W. The phylogenetic position of the Thermococcus isolate AN1 based on 16S rRNA gene sequence analysis: A proposal that AN1 represents a new species, Thermococcus zilligii sp. nov. Arch. Microbiol. 1997, 168, 245–248. [Google Scholar] [CrossRef]
- Miroshnichenko, M.L.; Hippe, H.; Stackebrandt, E.; Kostrikina, N.A.; Chernyh, N.A.; Jeanthon, C.; Nazina, T.N.; Belyaev, S.S.; Bonch-Osmolovskaya, E.A. Isolation and characterization of Thermococcus sibiricus sp. nov. from a Western Siberia high-temperature oil reservoir. Extremophiles 2001, 5, 85–91. [Google Scholar] [CrossRef]
- Cai, M.; Nie, Y.; Chi, C.Q.; Tang, Y.Q.; Li, Y.; Wang, X.B.; Liu, Z.S.; Yang, Y.F.; Zhou, J.Z.; Wu, X.L. Crude oil as a microbial seed bank with unexpected functional potentials. Sci. Rep. 2015, 5, 16057. [Google Scholar] [CrossRef]
- Magot, M.; Ollivier, B.; Patel, B.K. Microbiology of petroleum reservoirs. Antonie Van Leeuwenhoek 2000, 77, 103–116. [Google Scholar] [CrossRef]
- Pannekens, M.; Kroll, L.; Muller, H.; Mbow, F.T.; Meckenstock, R.U. Oil reservoirs, an exceptional habitat for microorganisms. New Biotechnol. 2019, 49, 1–9. [Google Scholar] [CrossRef]
- Dahle, H.; Birkeland, N.K. Thermovirga lienii gen. nov., sp. nov., a novel moderately thermophilic, anaerobic, amino-acid-degrading bacterium isolated from a North Sea oil well. Int. J. Syst. Evol. Microbiol. 2006, 56, 1539–1545. [Google Scholar] [CrossRef] [Green Version]
- Dahle, H.; Garshol, F.; Madsen, M.; Birkeland, N.K. Microbial community structure analysis of produced water from a high-temperature North Sea oil-field. Anton Leeuw Int. J. G 2008, 93, 37–49. [Google Scholar] [CrossRef]
- Stetter, K.O. Archaeoglobus fulgidus gen. nov, sp. nov.: A new taxon of extremely thermophilic archaebacteria. Syst. Appl. Microbiol. 1988, 10, 172–173. [Google Scholar] [CrossRef]
- Beeder, J.; Nilsen, R.K.; Rosnes, J.T.; Torsvik, T.; Lien, T. Archaeoglobus fulgidus Isolated from Hot North-Sea-Oil Field Waters. Appl. Environ. Microbol. 1994, 60, 1227–1231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labes, A.; Schönheit, P. Sugar utilization in the hyperthermophilic, sulfate-reducing archaeon Archaeoglobus fulgidus strain 7324: Starch degradation to acetate and CO2 via a modified Embden-Meyerhof pathway and acetyl-CoA synthetase (ADP-forming). Arch. Microbiol. 2001, 176, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Birkeland, N.K.; Schönheit, P.; Poghosyan, L.; Fiebig, A.; Klenk, H.P. Complete genome sequence analysis of Archaeoglobus fulgidus strain 7324 (DSM 8774), a hyperthermophilic archaeal sulfate reducer from a North Sea oil field. Stand. Genom. Sci. 2017, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balch, W.E.; Fox, G.E.; Magrum, L.J.; Woese, C.R.; Wolfe, R.S. Methanogens: Reevaluation of a unique biological group. Microbiol. Rev. 1979, 43, 260–296. [Google Scholar] [CrossRef] [PubMed]
- Lien, T.; Madsen, M.; Rainey, F.A.; Birkeland, N.K. Petrotoga mobilis sp. nov., from a North Sea oil-production well. Int. J. Syst. Bacteriol. 1998, 48, 1007–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shurigin, V.; Hakobyan, A.; Panosyan, H.; Egamberdieva, D.; Davranov, K.; Birkeland, N.K. A glimpse of the prokaryotic diversity of the Large Aral Sea reveals novel extremophilic bacterial and archaeal groups. MicrobiologyOpen 2019, 8, e00850. [Google Scholar] [CrossRef] [Green Version]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [Green Version]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A Comprehensive, Accurate, and Fast Distance-Based Phylogeny Inference Program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farris, J.S. Estimating Phylogenetic Trees from Distance Matrices. Am. Nat. 1972, 106, 645. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Petty, N.K.; Ben Zakour, N.L.; Beatson, S.A. BLAST Ring Image Generator (BRIG): Simple prokaryote genome comparisons. BMC Genom. 2011, 12, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labes, A.; Schönheit, P. Unusual starch degradation pathway via cyclodextrins in the hyperthermophilic sulfate-reducing archaeon Archaeoglobus fulgidus strain 7324. J. Bacteriol. 2007, 189, 8901–8913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aalen, N.; Steen, I.H.; Birkeland, N.K.; Lien, T. Purification and properties of an extremely thermostable NADP(+)-specific glutamate dehydrogenase from Archaeoglobus fulgidus. Arch. Microbiol. 1997, 168, 536–539. [Google Scholar] [CrossRef]
- Gebhard, S. Zuckerstoffwechsel von Hyperthermophilen Archaea. Master’s Thesis, University of Kiel, Kiel, Germany, 2003. [Google Scholar]
- Labes, A.; Schönheit, P. ADP-dependent glucokinase from the hyperthermophilic sulfate-reducing archaeon Archaeoglobus fulgidus strain 7324. Arch. Microbiol. 2003, 180, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Hansen, T.; Schönheit, P. ADP-dependent 6-phosphofructokinase, an extremely thermophilic, non-allosteric enzyme from the hyperthermophilic, sulfate-reducing archaeon Archaeoglobus fulgidus strain 7324. Extremophiles 2004, 8, 29–35. [Google Scholar] [CrossRef]
- Johnsen, U.; Hansen, T.; Schönheit, P. Comparative analysis of pyruvate kinases from the hyperthermophilic archaea Archaeoglobus fulgidus, Aeropyrum pernix, and Pyrobaculum aerophilum and the hyperthermophilic bacterium Thermotoga maritima—Unusual regulatory properties in hyperthermophilic archaea. J. Biol. Chem. 2003, 278, 25417–25427. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| T. bergensis T7324T | T. litoralis DSM 5473T | |
|---|---|---|
| GenBank accession | JABFNK000000000 | CP006670.1 |
| Number of contigs | 5 (draft) | 1 (complete) |
| Number of base pairs | 2,077,832 | 2,215,172 |
| GC% | 44.1 | 43.1 |
| Number of rRNAs | 2, 1, 1 (5S, 16S, 23S) | 2, 1, 1 (5S, 16S, 23S) |
| Number of genes (total) | 2370 | 2575 |
| Number of CRISPRs | 6 arrays, 129 spacers * | 7 arrays, 251 spacers * |
| Enzyme | Reference | Locus Tag (in T. bergensis) |
|---|---|---|
| Cyclodextrin gluconotransferase (CGTase) | [27] | GQS78_01870 |
| Maltodextrin–phosphorylase (MalP)C | [27] | GQS78_09095 |
| Phosphoglucomutase (PGM) | [27] | GQS78_06950 |
| ADP-dependent glucokinase | [30] | GQS78_06970 |
| ADP-dependent phosphofructokinase | [31] | GQS78_11830 |
| Pyruvate kinase | [32] | GQS78_00030 |
| Glutamate dehydrogenase | [28] | GQS78_10575 |
| Glyceraldehyde-3-phosphate ferredoxin oxidoreductase | [29] | GQS78_03205 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Birkeland, N.-K.; Bunk, B.; Spröer, C.; Klenk, H.-P.; Schönheit, P. Thermococcus bergensis sp. nov., a Novel Hyperthermophilic Starch-Degrading Archaeon. Biology 2021, 10, 387. https://doi.org/10.3390/biology10050387
Birkeland N-K, Bunk B, Spröer C, Klenk H-P, Schönheit P. Thermococcus bergensis sp. nov., a Novel Hyperthermophilic Starch-Degrading Archaeon. Biology. 2021; 10(5):387. https://doi.org/10.3390/biology10050387
Chicago/Turabian StyleBirkeland, Nils-Kåre, Boyke Bunk, Cathrin Spröer, Hans-Peter Klenk, and Peter Schönheit. 2021. "Thermococcus bergensis sp. nov., a Novel Hyperthermophilic Starch-Degrading Archaeon" Biology 10, no. 5: 387. https://doi.org/10.3390/biology10050387
APA StyleBirkeland, N. -K., Bunk, B., Spröer, C., Klenk, H. -P., & Schönheit, P. (2021). Thermococcus bergensis sp. nov., a Novel Hyperthermophilic Starch-Degrading Archaeon. Biology, 10(5), 387. https://doi.org/10.3390/biology10050387