
Entomopathogenic Nematodes for the Management of Plum Curculio in Highbush Blueberry

 ,
,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Laboratory Experiments
2.1.1. Soils, Insects, Nematodes
2.1.2. Nematode Persistence in Different Soil Types
2.1.3. Nematodes Virulence of against C. nenuphar Larvae and Pupae
2.2. Field Experiment: 2020
2.3. Field Experiment: 2021
2.4. Statistical Analysis
3. Results
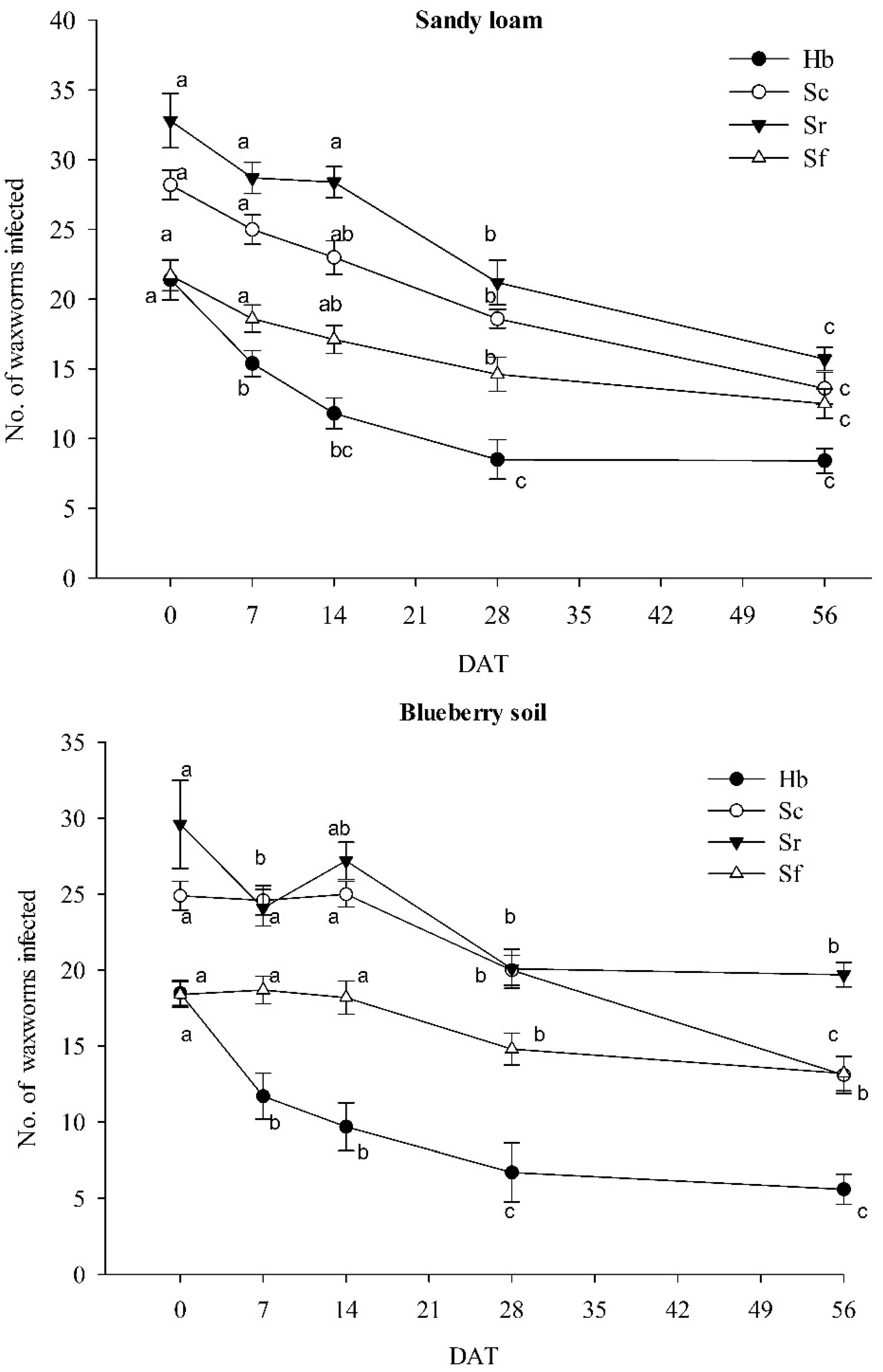
3.1. Nematode Persistence in Different Soil Types
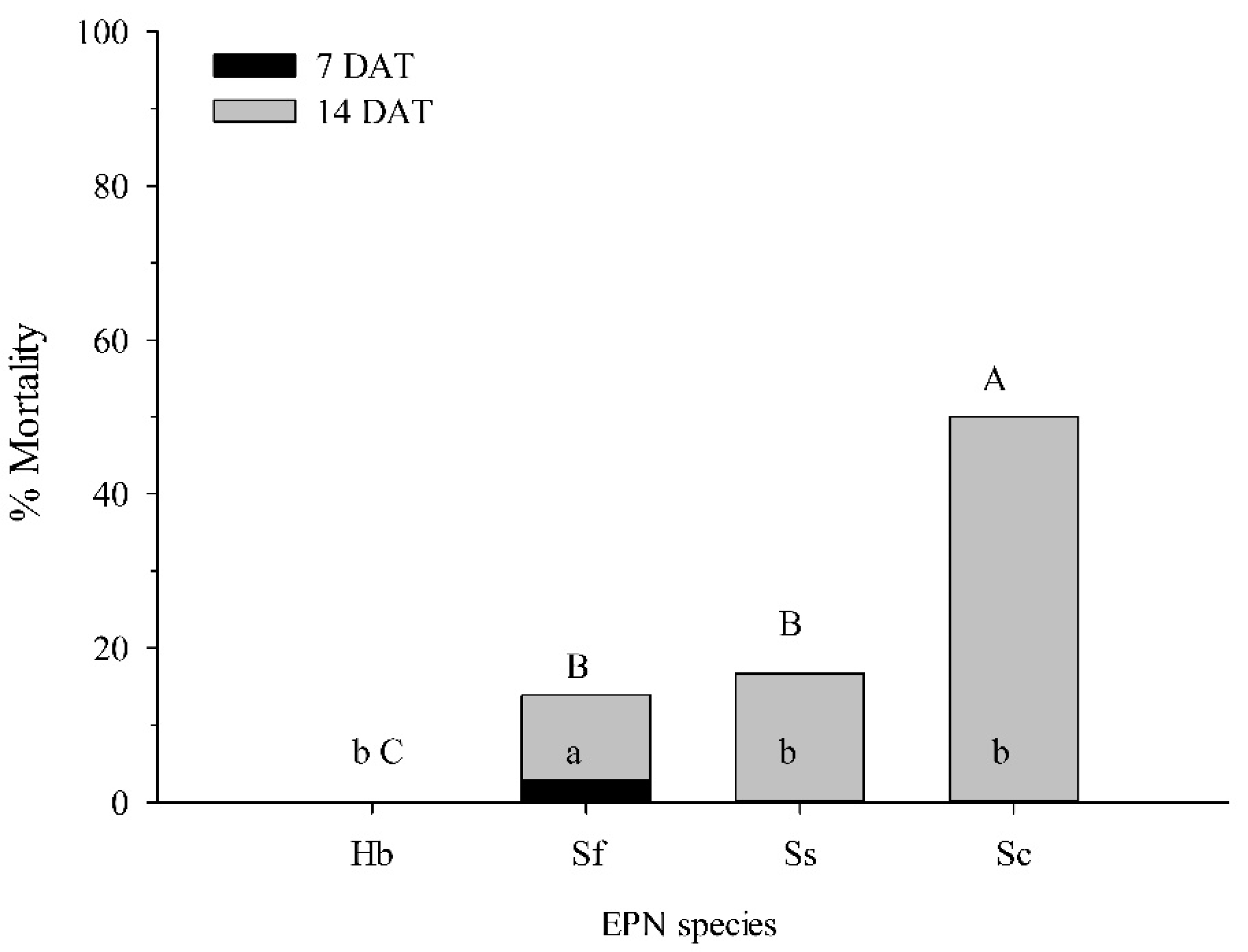
3.2. Virulence of EPNs to C. nenuphar Larvae
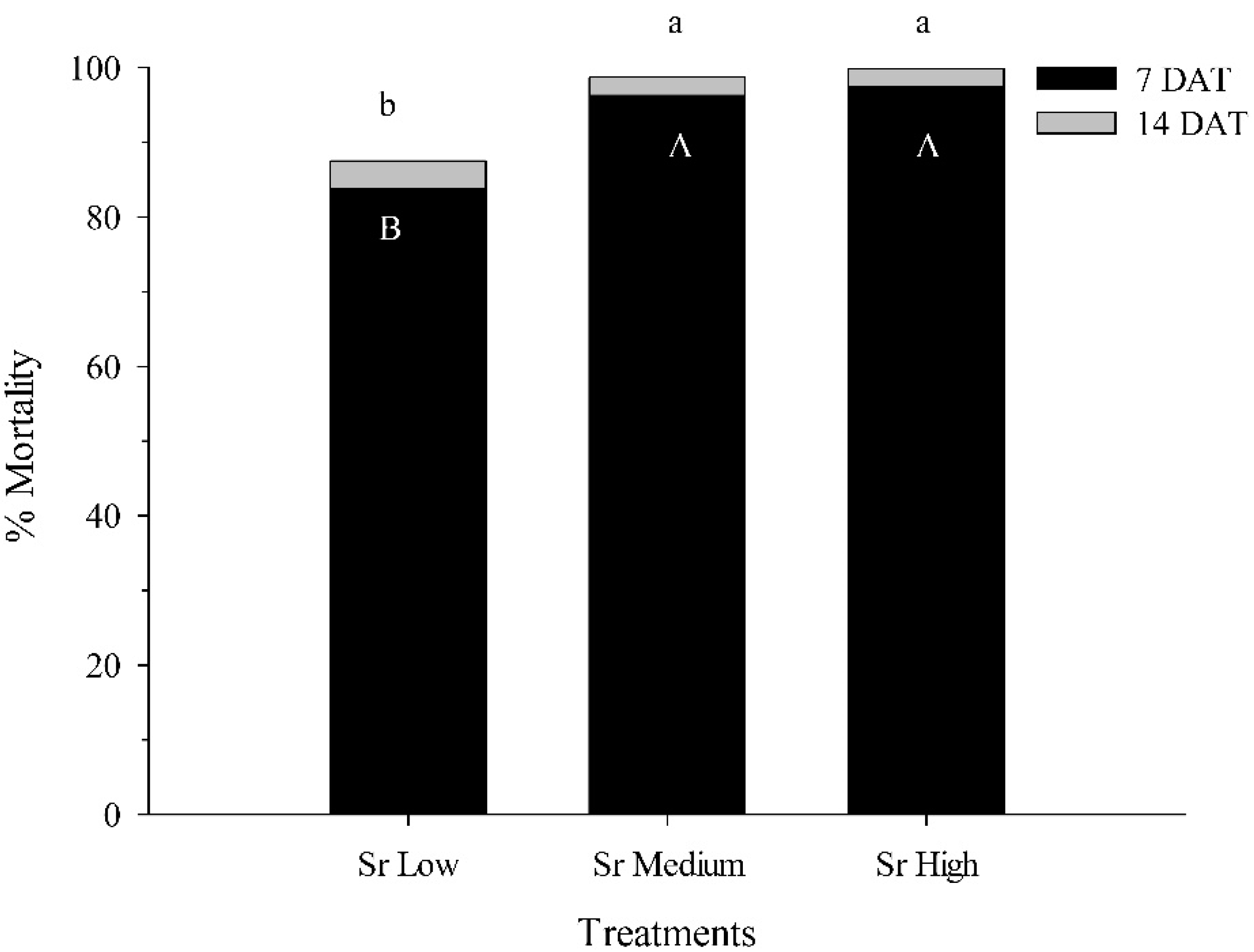
3.3. Virulence of EPNs to C. nenuphar Pupae
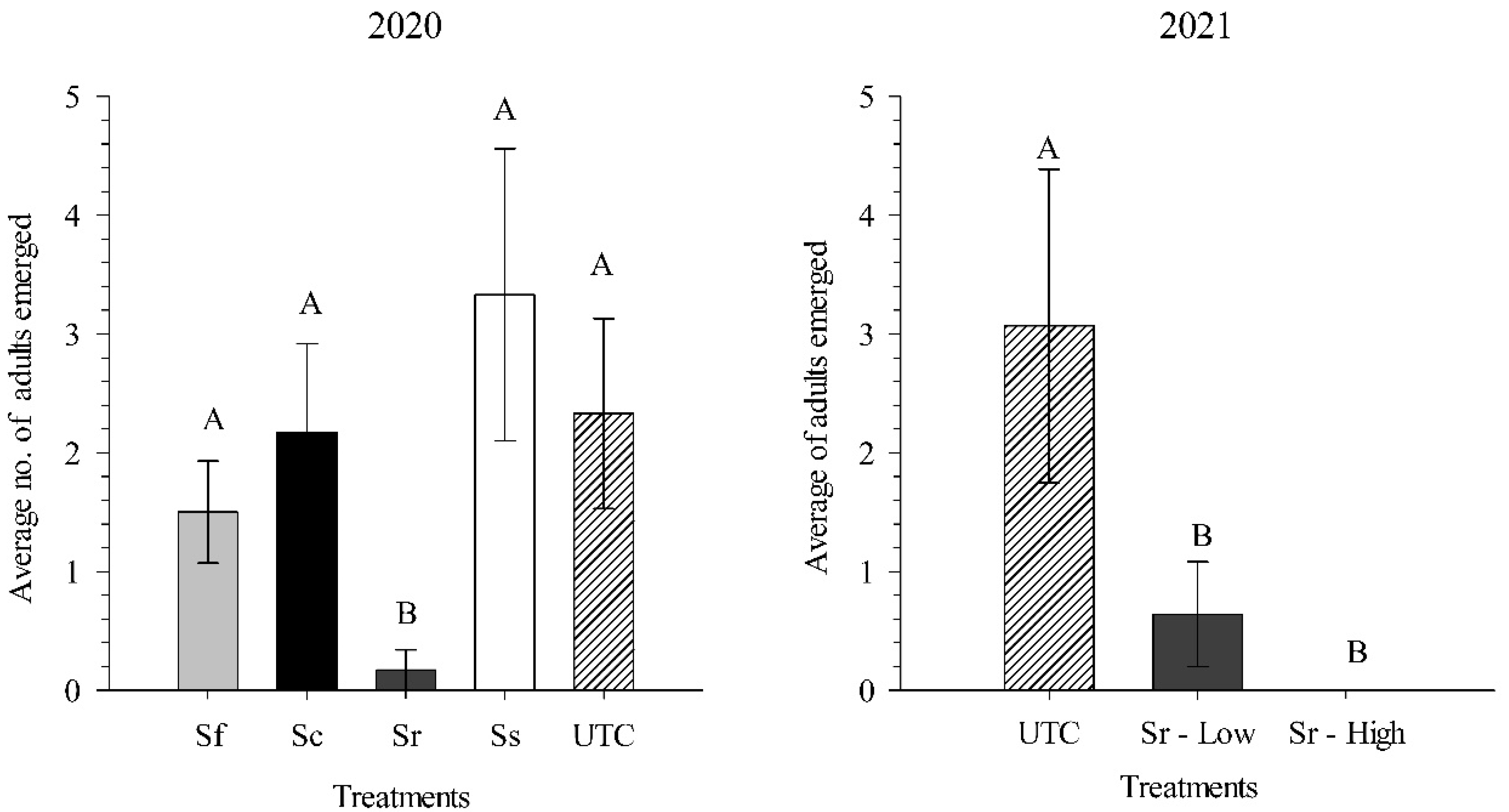
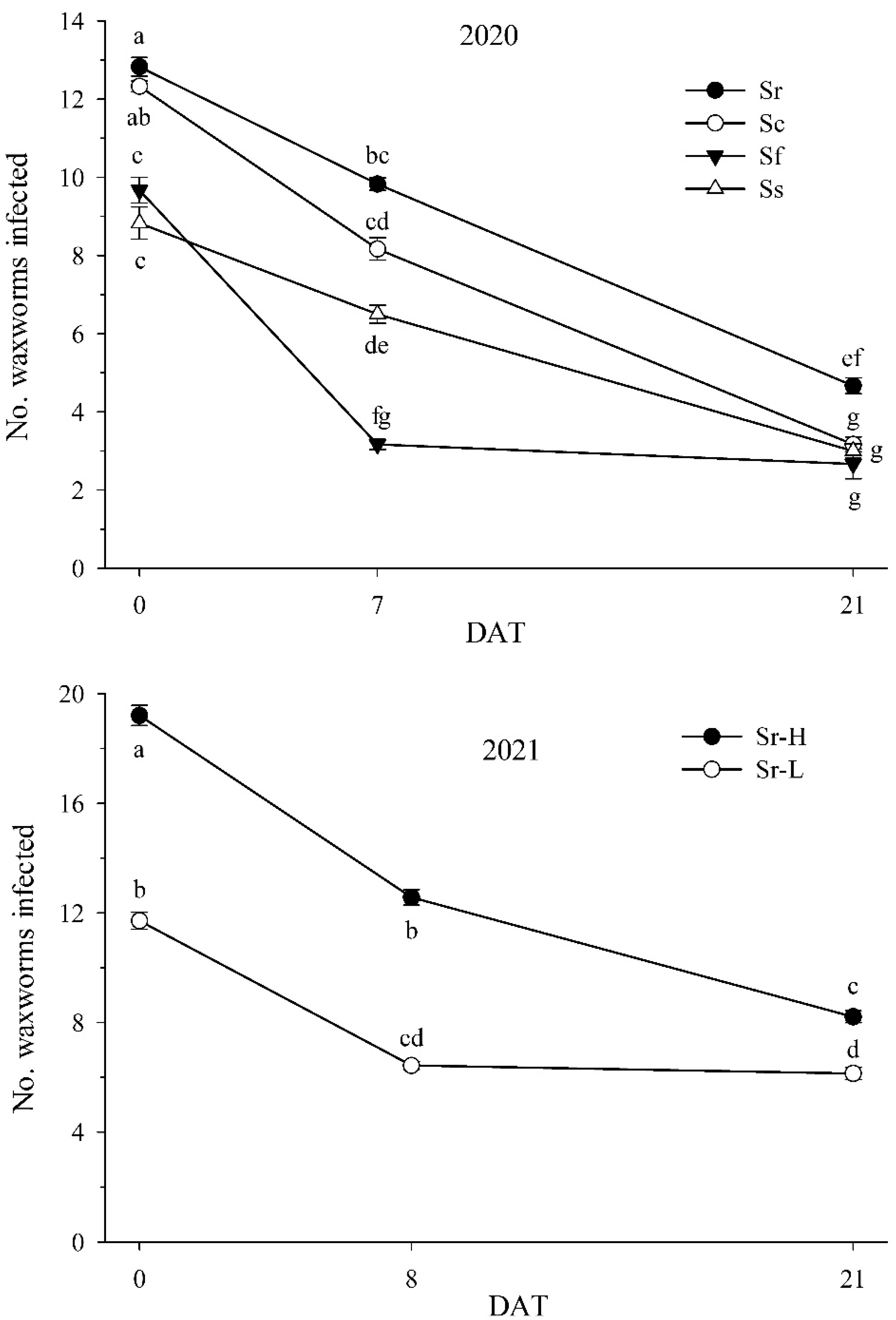
3.4. Field Experiments: 2020 and 2021
3.5. Nematode Field Persistence: 2020 and 2021
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Racette, G.; Chouinard, G.; Vincent, C.; Hill, S.B. Ecology and management of plum curculio, Conotrachelus nenuphar [Coleoptera: Curculionidae], in apple orchards. Phytoprotection 1992, 73, 85–100. [Google Scholar] [CrossRef] [Green Version]
- Horton, D.; Johnson, D. (Eds.) Southeastern Peach Grower’s Handbook; University of Georgia, Cooperative Extension Service: Athens, GA, USA, 2005. [Google Scholar]
- Leskey, T.C.; Chouinard, G.; Vincent, C. Monitoring and management of apple maggot fly and plum curculio: Honouring the legacy of Ronald J. Prokopy. In Biorational Tree Fruit Management; Aluja, M., Leskey, T.C., Vincent, C., Eds.; CABI Press: Oxfordshire, UK, 2009; p. 295. [Google Scholar]
- Antonelli, A.; Elsner, E.; Shanks, C. Arthropod Management. In Highbush Blueberry Production Guide; Northeast Regional Agricultural Engineering Service-55; Cooperative Extension: Ithaca, NY, USA, 1992; pp. 55–75. [Google Scholar]
- Levine, E.; Hall, F.R. Effect of feeding and oviposition by the plum curculio on apple and plum fruit abscission. J. Econ. Entomol. 1977, 70, 603–607. [Google Scholar] [CrossRef]
- Quaintance, A.L.; Jenne, E.L. The Plum Curculio (No. 103); US Department of Agriculture, Bureau of Entomology: Washington, DC, USA, 1912. [Google Scholar]
- Lampasona, T.P.; Rodriguez-Saona, C.; Leskey, T.C.; Nielsen, A.L. A Review of the biology, ecology, and management of plum curculio (Coleoptera: Curculionidae). J. Integr. Pest Manag. 2020, 11, 22. [Google Scholar] [CrossRef]
- Olthof, T.H.; Hagley, E.A.C. Laboratory studies of the efficacy of steinernematid nematodes against the plum curculio (Coleoptera: Curculionidae). J. Econ. Entomol. 1993, 86, 1078–1082. [Google Scholar] [CrossRef]
- Horton, D.; Brannen, P.; Bellinger, B.; Lockwood, D.; Ritchie, D. Southeastern Peach, Nectarine, and Plum Pest Management and Culture Guide, Bulletin 1171; University of Georgia: Athens, GA, USA, 2013. [Google Scholar]
- Shapiro-Ilan, D.I.; Mizell, R.F.; Campbell, J.F. Susceptibility of the plum curculio, Conotrachelus nenuphar, to entomopathogenic nematodes. J. Nematol. 2002, 34, 246–249. [Google Scholar] [PubMed]
- Shapiro-Ilan, D.I.; Mizell, R.F.; Cottrell, T.E.; Horton, D.L. Measuring field efficacy of Steinernema feltiae and Steinernema riobrave for suppression of plum curculio, Conotrachelus nenuphar, larvae. Biol. Control 2004, 30, 496–503. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Mizell III, R.F.; Cottrell, T.E.; Horton, D.L. Control of plum curculio, Conotrachelus nenuphar with entomopathogenic nematodes: Effects of application timing, alternate host plant, and nematode strain. Biol. Control 2008, 44, 207–215. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Leskey, T.C.; Wright, S.E. Virulence of entomopathogenic nematodes to plum curculio, Conotrachelus nenuphar: Effects of strain, temperature, and soil type. J. Nematol. 2011, 43, 187–195. [Google Scholar] [PubMed]
- Shapiro-Ilan, D.I.; Wright, S.E.; Tuttle, A.F.; Cooley, D.R.; Leskey, T.C. Using entomopathogenic nematodes for biological control of plum curculio, Conotrachelus nenuphar: Effects of irrigation and species in apple orchards. Biol. Control 2013, 67, 123–129. [Google Scholar] [CrossRef]
- Kaya, H.K.; Gaugler, R. Entomopathogenic nematodes. Annu. Rev. Entomol. 1993, 38, 181–206. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Duncan, L.W.; Lacey, L.A.; Han, R. Orchard crops. In Nematodes as Biological Control Agents; Grewal, P., Ehlers, R.-U., Shapiro-Ilan, D.I., Eds.; CABI Publishing: Wallingford, UK, 2005; pp. 215–230. [Google Scholar]
- Pereault, R.J.; Whalon, M.E.; Alston, D.G. Field efficacy of entomopathogenic fungi and nematodes targeting caged last-instar plum curculio (Coleoptera: Curculionidae) in Michigan cherry and apple orchards. Environ. Entomol. 2009, 38, 1126–1134. [Google Scholar] [CrossRef] [PubMed]
- Piñero, J.C.; Shapiro-Ilan, D.I.; Cooley, D.R.; Tuttle, A.F.; Eaton, A.; Drohan, P.; Leahy, K.; Zhang, A.; Hancock, T.; Wallingford, A.K.; et al. Toward the integration of an attract-and-kill approach with entomopathogenic nematodes to control multiple life stages of plum curculio (Coleoptera: Curculionidae). Insects 2020, 11, 375. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Bruck, D.J.; Lacey, L.A. Principles of epizootiology and microbial control. Insect Pathol. 2012, 2, 29–72. [Google Scholar]
- Koppenhöfer, A.M.; Fuzy, E.M. Effect of soil type on infectivity and persistence of the entomopathogenic nematodes Steinernema scarabaei, Steinernema glaseri, Heterorhabditis zealandica and Heterorhabditis bacteriophora. J. Invertebr. Pathol. 2006, 92, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, A.P. Filter-paper method for routine measurement of field water potential. J. Hydrol. 1981, 53, 355–360. [Google Scholar] [CrossRef]
- Amis, A.; Snow, J.J. Conotrachelus nenuphar. In Handbook of Insect Rearing; Singh, P., Moore, R.F., Eds.; Elsevier: Amsterdam, The Netherlands, 1985; pp. 227–236. [Google Scholar]
- Koppenhöfer, A.M.; Kaya, H.K.; Campbell, J.F.; Gaugler, R. Estimation of entomopathogenic nematode population density in soil by correlation between bait insect mortality and nematode penetration. Fund. Appl. Nematol. 1998, 21, 95–102. [Google Scholar]
- Polavarapu, S.; Koppenhöfer, A.M.; Barry, J.D.; Holdcraft, R.; Fuzy, E.M. Entomopathogenic nematodes and neonicotinoids for remedial control of oriental beetle, Anomala orientalis (Coleoptera: Scarabaeidae), in highbush blueberry. Crop Prot. 2007, 26, 1266–1271. [Google Scholar] [CrossRef]
- Koppenhöfer, A.M.; Fuzy, E.M. Long-term effects and persistence of Steinernema scarabaei applied for suppression of Anomala orientalis (Coleoptera: Scarabaeidae). Biol. Control 2009, 48, 63–72. [Google Scholar] [CrossRef]
- Kaya, H.K.; Stock, S.P. Techniques in insect nematology. In Manual of Techniques in Insect Pathology; Lacey, L.A., Ed.; Academic Press: San Diego, CA, USA, 1997; pp. 281–324. [Google Scholar]
- Koppenhöfer, A.M. Nematodes. In Field Manual of Techniques for the Application and Evaluation of Entomopathogens, 2nd ed.; Lacey, L., Kaya, H., Eds.; Kluwer: Dordrecht, The Netherlands, 2007; pp. 249–264. [Google Scholar]
- Baur, M.E.; Kaya, H.K. Persistence of entomopathogenic nematodes. In Factors Affecting the Survival of Entomopathogens; Baur, M.E., Fuxa, J.R., Eds.; Southern Cooperative Series Bulletin: Fayetteville, AR, USA, 2001. [Google Scholar]
- Koppenhöfer, A.M.; Fuzy, E.M. Soil moisture effects on infectivity and persistence of the entomopathogenic nematodes Steinernema scarabaei, S. glaseri, Heterorhabditis zealandica, and H. bacteriophora. Appl. Soil Ecol. 2007, 35, 128–139. [Google Scholar] [CrossRef]
- Alston, D.G.; Rangel, D.E.N.; Lacey, L.A.; Golez, H.G.; Kim, J.J.; Roberts, D.W. Evaluation of novel fungus and nematode isolates for control of Conotrachelus nenuphar (Coleoptera: Curculionidae) larvae. Biol. Control 2005, 35, 163–171. [Google Scholar] [CrossRef]
- Shapiro-Ilan, D.I.; Arthurs, S.P.; Lacey, L.A. Microbial control of arthropod pests of orchards in temperate climates. In Microbial Control of Insect and Mite Pests; Lacey, L.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 253–267. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IJs/larva | S. feltiae | S. carpocapsae | S. riobrave | S. scarabaei |
|---|---|---|---|---|
| 7 DAT | ||||
| 100 | 1.4 ± 1.4 Cb | 2.4 ± 2.4 Bc | 30 ± 8.1 Ac | 0.0 ± 0.0 Da |
| 200 | 1.4 ± 1.4 Cb | 8.6 ± 4.6 Bb | 55.7 ± 9.4 Ab | 0.0 ± 0.0 Da |
| 400 | 2.9 ± 2.9 Ca | 35.7 ± 10.5 Ba | 78.6 ± 7.4 Aa | 0.0 ± 0.0 Da |
| 14 DAT | ||||
| 100 | 34.3 ± 8.0 Bc | 41.4 ± 5.7 Bc | 94.3 ± 2.3 Ab | 7.1 ± 2.7 Cc |
| 200 | 41.4 ± 6.4 Bb | 55.7 ± 8.1 Bb | 95.7 ± 2.5 Aa | 27.1 ± 6.2 Cb |
| 400 | 62.9 ± 7.2 Ba | 61.4 ± 8.8 Ba | 98.6 ± 1.4 Aa | 37.1 ± 4.6 Ca |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sousa, A.L.; Rodriguez-Saona, C.; Holdcraft, R.; Kyryczenko-Roth, V.; Koppenhöfer, A.M. Entomopathogenic Nematodes for the Management of Plum Curculio in Highbush Blueberry. Biology 2022, 11, 45. https://doi.org/10.3390/biology11010045
Sousa AL, Rodriguez-Saona C, Holdcraft R, Kyryczenko-Roth V, Koppenhöfer AM. Entomopathogenic Nematodes for the Management of Plum Curculio in Highbush Blueberry. Biology. 2022; 11(1):45. https://doi.org/10.3390/biology11010045
Chicago/Turabian StyleSousa, Ana Luiza, Cesar Rodriguez-Saona, Robert Holdcraft, Vera Kyryczenko-Roth, and Albrecht M. Koppenhöfer. 2022. "Entomopathogenic Nematodes for the Management of Plum Curculio in Highbush Blueberry" Biology 11, no. 1: 45. https://doi.org/10.3390/biology11010045
APA StyleSousa, A. L., Rodriguez-Saona, C., Holdcraft, R., Kyryczenko-Roth, V., & Koppenhöfer, A. M. (2022). Entomopathogenic Nematodes for the Management of Plum Curculio in Highbush Blueberry. Biology, 11(1), 45. https://doi.org/10.3390/biology11010045