
The Characterization of the Tobacco-Derived Wild Tomato Mosaic Virus by Employing Its Infectious DNA Clone
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plants Growth Conditions
2.2. Virus Samples COLLECTION and Preservation
2.3. Virus Detection by Western Blot and RT-PCR
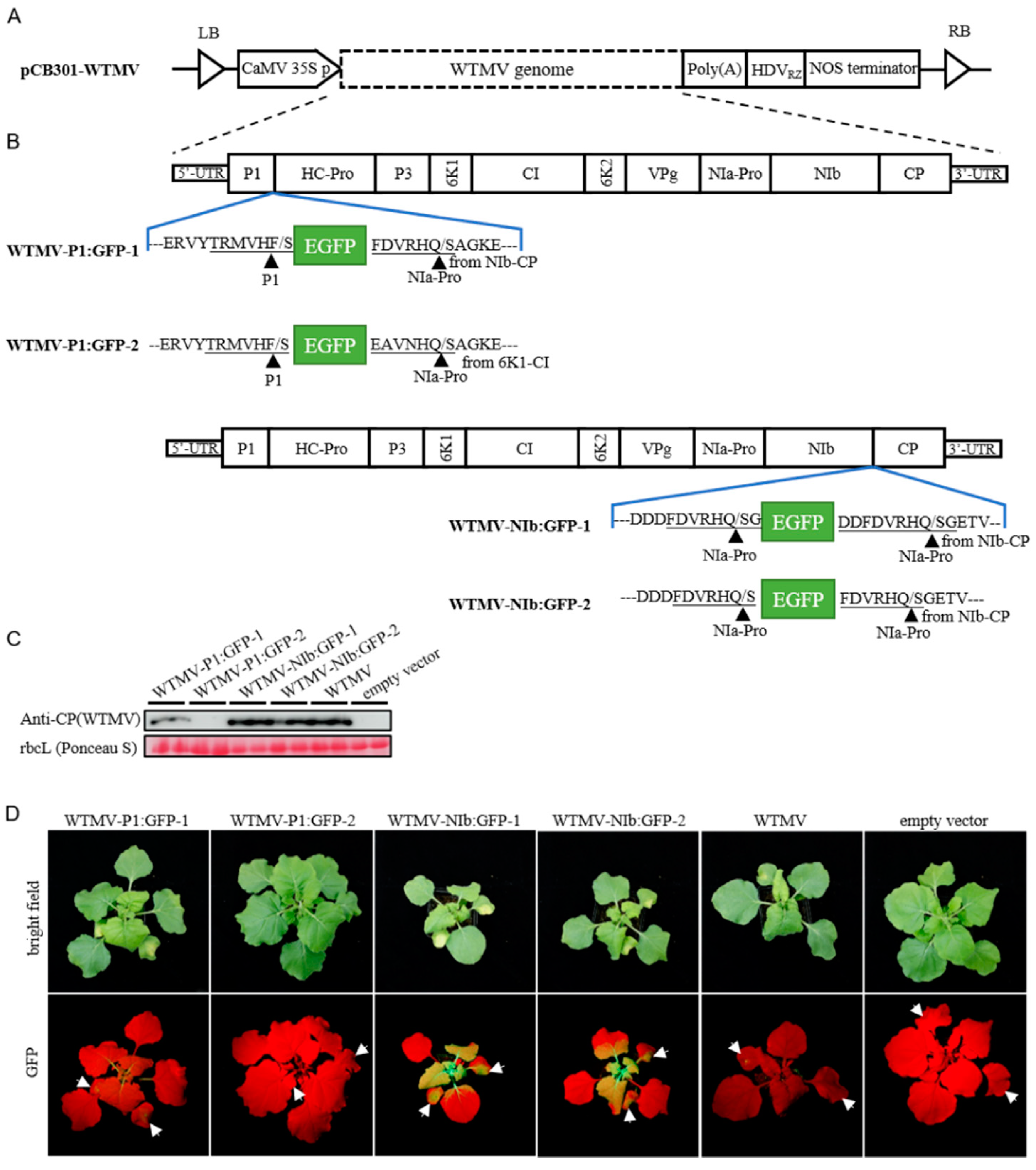
2.4. Infectious Clones Construction
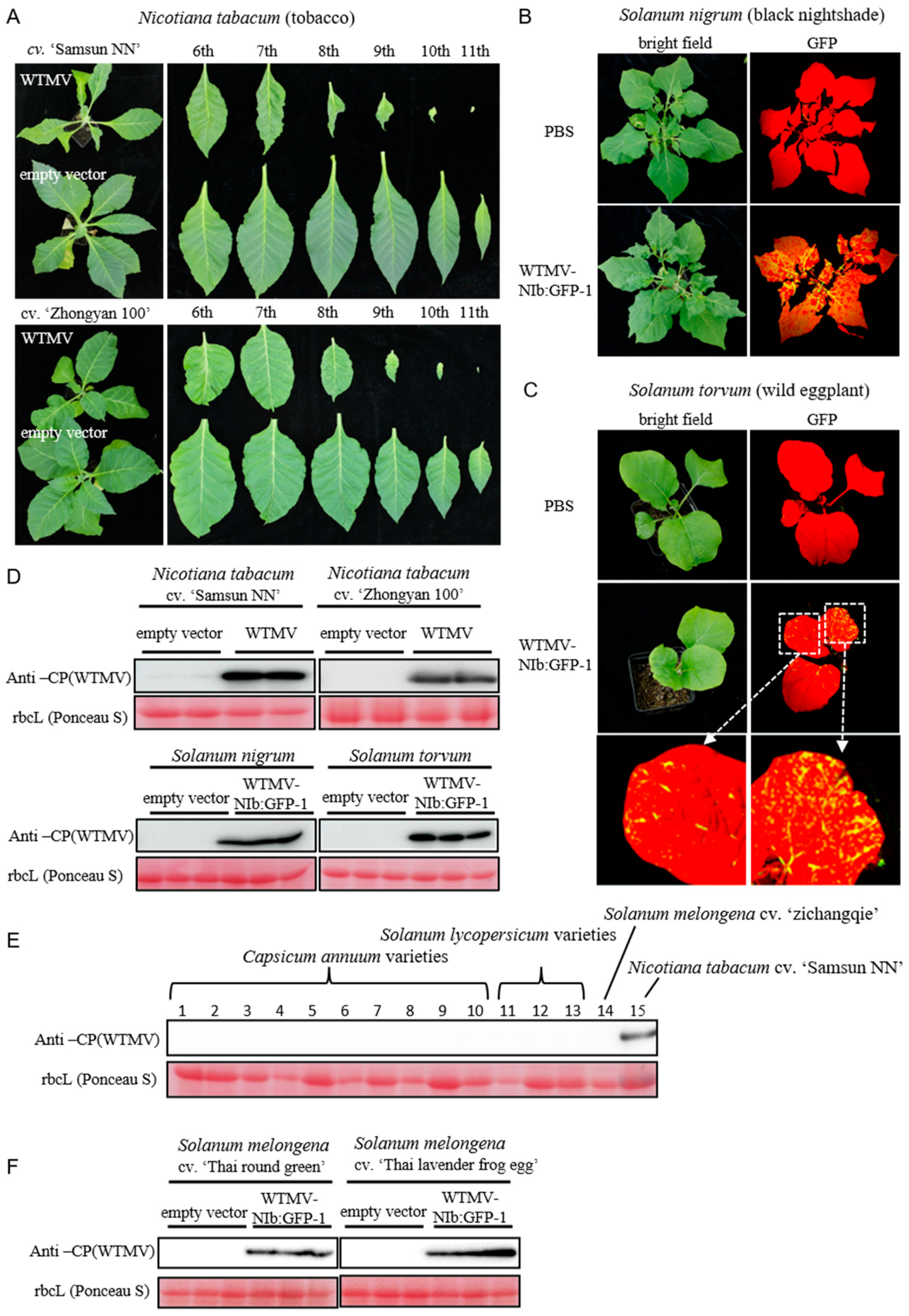
2.5. Plant Inoculation and Phenotyping
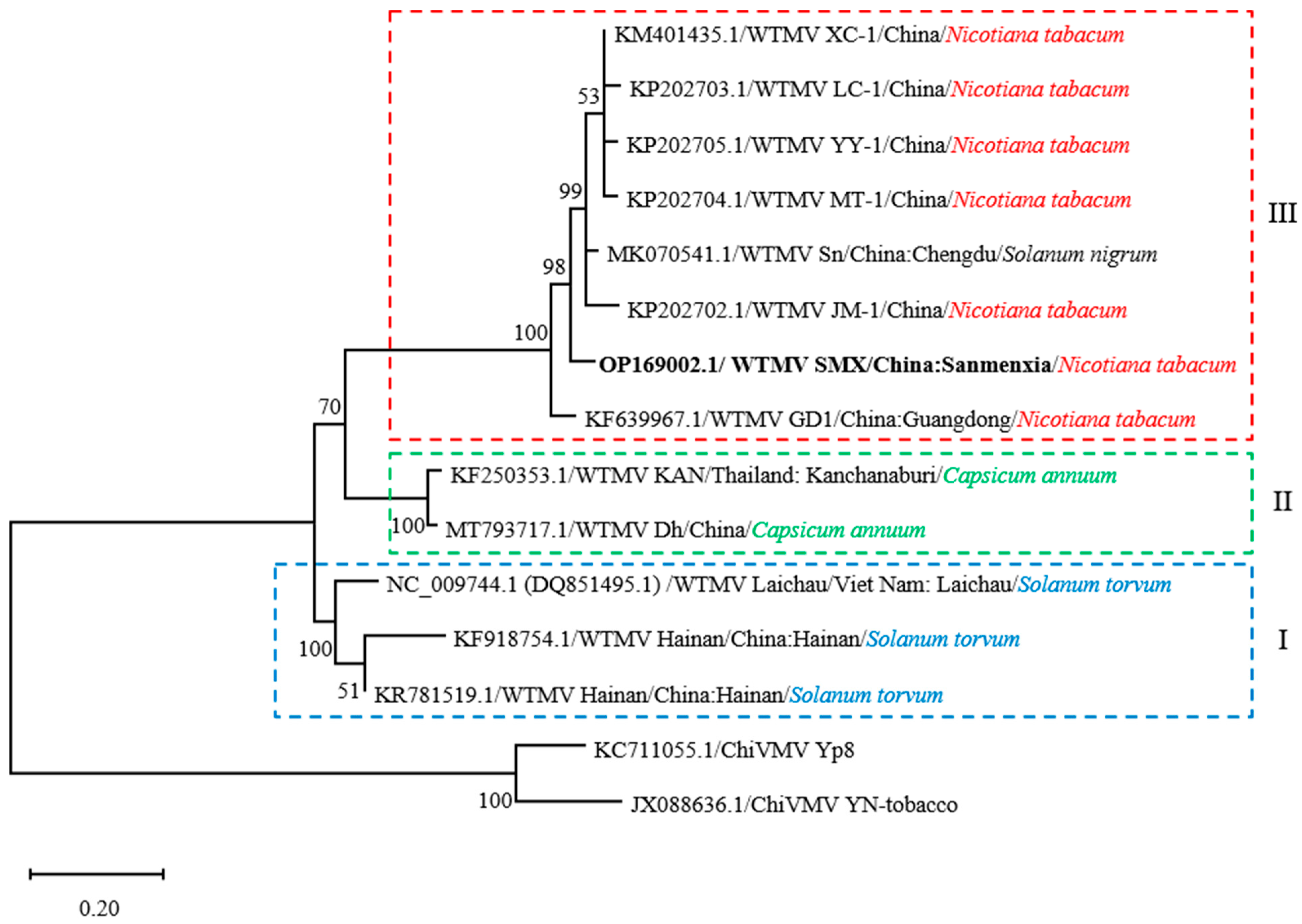
2.6. Phylogenetic Reconstruction
3. Results
3.1. Genome Sequence and Infectious DNA Clone of Tobacco-Derived WTMV
3.2. Evolutionary Relationship of WTMV Isolates
3.3. Infectivity of WTMV-SMX on Potential Hosts
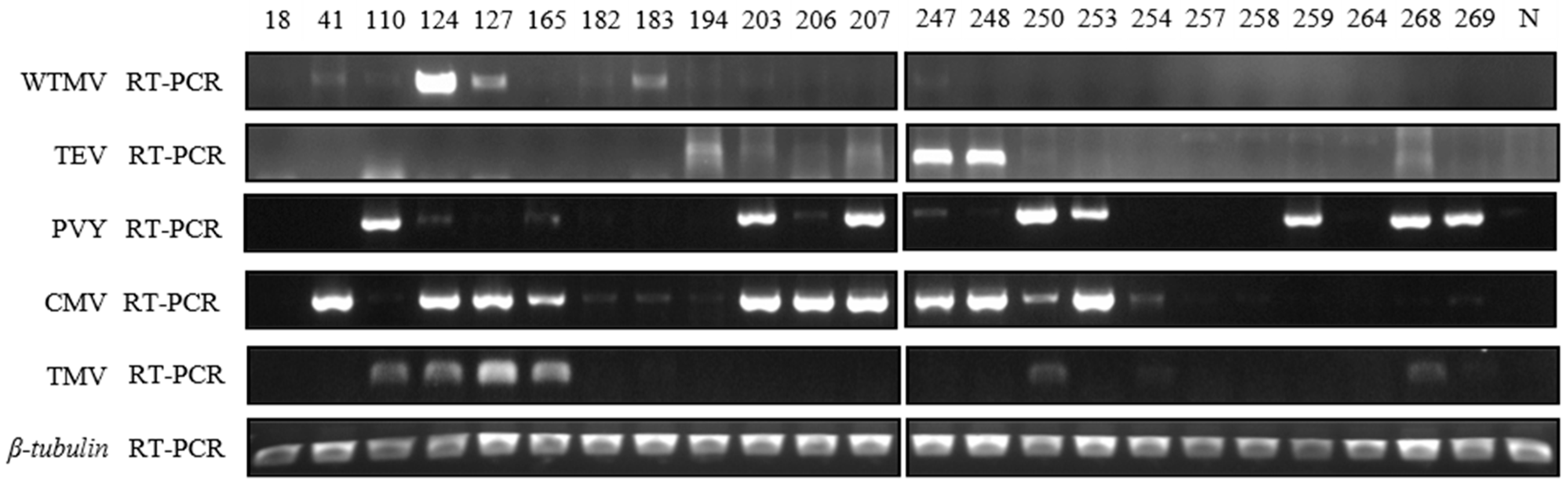
3.4. Field Survey of Major Tobacco Growing Areas in Henan Province of China in 2021
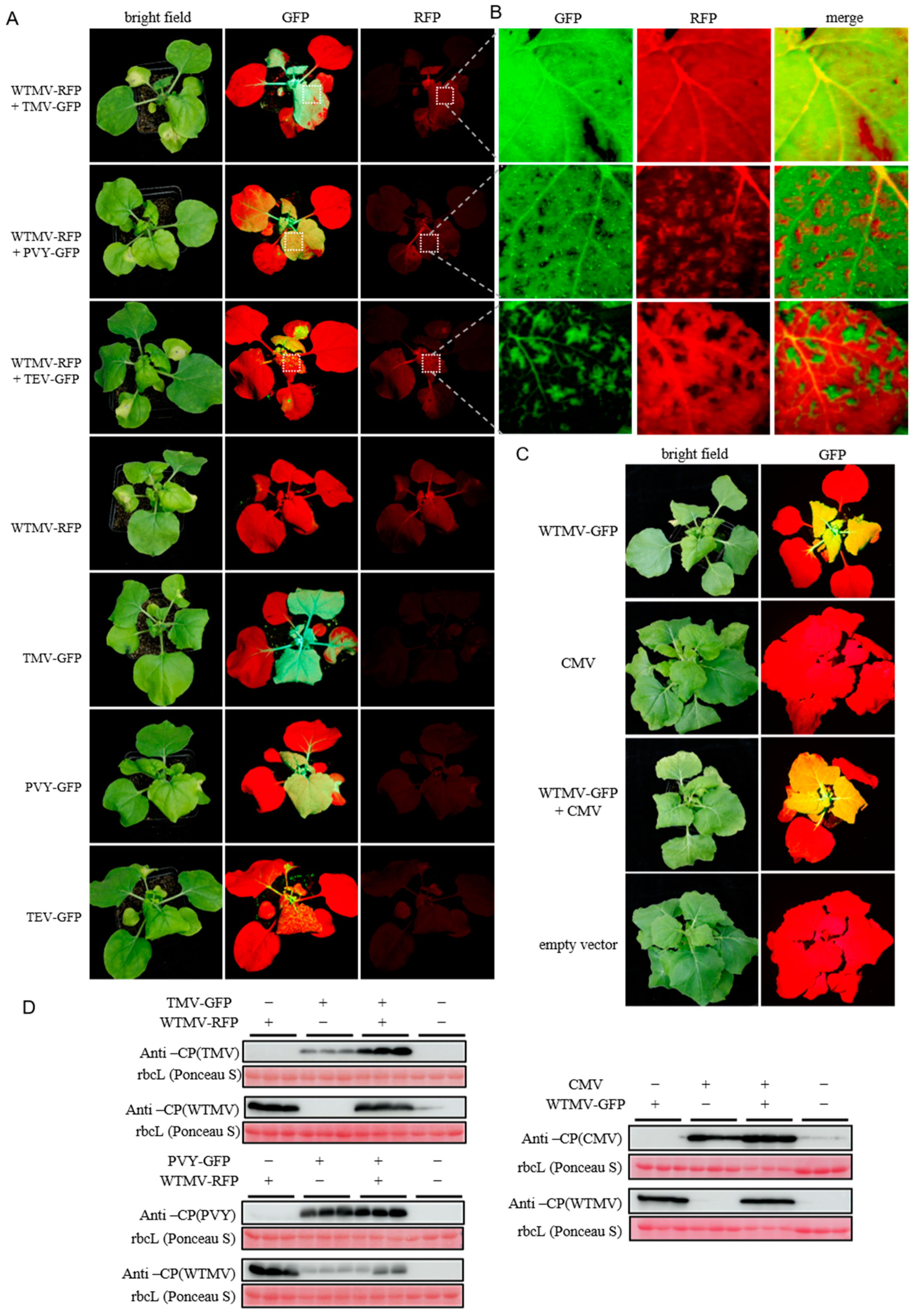
3.5. Coinfection of WTMV with Other Viruses Reveals SIE between Potyvirus Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Von Wettberg, E.; Davis, T.M.; Smykal, P. Editorial: Wild Plants as Source of New Crops. Front. Plant Sci. 2020, 11, 591554. [Google Scholar] [CrossRef] [PubMed]
- Hasiow-Jaroszewska, B.; Boezen, D.; Zwart, M.P. Metagenomic Studies of Viruses in Weeds and Wild Plants: A Powerful Approach to Characterise Variable Virus Communities. Viruses 2021, 13, 1939. [Google Scholar] [CrossRef] [PubMed]
- Susi, H.; Filloux, D.; Frilander, M.J.; Roumagnac, P.; Laine, A.L. Diverse and variable virus communities in wild plant populations revealed by metagenomic tools. PeerJ 2019, 7, e6140. [Google Scholar] [CrossRef]
- Harrison, J.P. The Evolution of the Colombian Tobacco Trade, to 1875. Hisp. Am. Hist. Rev. 1952, 32, 163–174. [Google Scholar] [CrossRef]
- Scholthof, K.B. Tobacco Mosaic Virus: The Beginning of Plant Virology. APSnet Features 2008. [Google Scholar] [CrossRef]
- Holmes, F.O. Indications of a New-World origin of tobacco-mosaic virus. Phytopathology 1951, 41, 341–349. [Google Scholar]
- Gibbs, A. Evolution and origins of tobamoviruses. Philos. Trans. R Soc. Lond. B Biol. Sci. 1999, 354, 593–602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gadhave, K.R.; Gautam, S.; Rasmussen, D.A.; Srinivasan, R. Aphid Transmission of Potyvirus: The Largest Plant-Infecting RNA Virus Genus. Viruses 2020, 12, 773. [Google Scholar] [CrossRef] [PubMed]
- Witt, A.; Luke, W.R.Q. Guide to the Naturalized and Invasive Plants of Eastern Africa; Centre for Agriculture and Bioscience International: Wallingford, UK, 2017. [Google Scholar] [CrossRef]
- Blakeslee, A.F. A graft-infectious disease of Datura resembling a vegetative mutation. J. Genet. 1921, 11, 17–36. [Google Scholar] [CrossRef]
- Anderson, C.W. Cassia tora, a leguminous host of Tobacco etch virus. Plant Dis. Report. 1954, 38, 736–738. [Google Scholar]
- Debrot, C. Cassia tora L., a natural host of the tobacco etch virus in Venezuela. Agron. Trop. 1974, 24, 21–26. [Google Scholar]
- McGovern, R.J.; Polston, J.E.; Mullahey, J.J. Solanum viarum: Weed reservoir of plant viruses in Florida. Int. J. Pest Manag. 1994, 40, 270–273. [Google Scholar] [CrossRef]
- Demski, J.W. The epidemiology of tobacco etch virus-infected Cassia obtusifolia in relation to pepper. Plant Dis. Report. 1979, 63, 647–650. [Google Scholar]
- Doolittle, S.P.; Alexander, L.J. Occurrence of Tobacco etch virus on greenhouse Tomatoes in Ohio. Plant Dis. Report. 1951, 35, 351–353. [Google Scholar]
- Wilfert Eckel, R.V.; Lampert, E.P. Effect of tobacco etch virus on the seasonal growth of flue-cured tobacco. Crop Prot. 1993, 12, 505–512. [Google Scholar] [CrossRef]
- Bai, J.; Li, S.; Li, C.; Wang, H.; Li, X.; Qiu, R.; Cheng, Y.; Zhou, S.; Song, R. Species and distribution of tobacco virus diseases in Henan Province. Tob. Sci. Technol. 2018, 51, 20–26. (In Chinese) [Google Scholar]
- Ha, C.; Coombs, S.; Revill, P.A.; Harding, R.M.; Vu, M.; Dale, J.L. Design and application of two novel degenerate primer pairs for the detection and complete genomic characterization of potyviruses. Arch. Virol. 2008, 153, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Fa, L.; Su, X.; Chen, Y.; Huang, J.; Yin, H.; Ma, G.; Wang, Y.; Zhang, J.; Zhang, Z.K.; et al. First report of wild tomato mosaic virus infecting chilli pepper in China. Plant Dis. 2021, 105, 3311. [Google Scholar] [CrossRef]
- Zhang, L.; Shang, J.; Jia, Q.; Gong, G.; Zhang, M.; Yang, W. The complete genome sequence of wild tomato mosaic virus isolated from Solanum nigrum reveals recombination in the P1 cistron. Arch. Virol. 2019, 164, 903–906. [Google Scholar] [CrossRef]
- Zhang, S.; Yu, N.; Wang, X.; Liang, J.; Xie, H.; Wang, J.; Zhang, Y.; Liu, Z. Natural Occurrence of Wild tomato mosaic virus in Wild Eggplant in China. J. Phytopathol. 2015, 163, 1023–1026. [Google Scholar] [CrossRef]
- Du, Z.G.; She, X.M.; Tang, Y.F.; He, Z.F.; Yang, J.G. First Report of Wild tomato mosaic virus Infecting Tobacco (Nicotiana tabacum) in China. Plant Dis. 2014, 98, 856. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Liu, W.; Yang, J.; Wang, F.; Shen, L.; Li, Y.; Sun, D.; Qian, Y. Complete Genome Sequence of a Novel Wild tomato mosaic virus Isolate Infecting Nicotiana tabacum in China. J. Phytopathol. 2016, 164, 686–690. [Google Scholar] [CrossRef]
- Bai, J.; Yang, X.; Li, Z.; Li, C.; Chen, Y.; Hu, J.; Liu, S.; Wang, X.; Li, D.; Li, S. A survey on viruses infecting tobacco in Henan Province by RNA-seq. Acta Phytopathol. Sin. 2021, 51, 658–662. (In Chinese) [Google Scholar]
- Zhang, W.; Qiu, Y.; Zhou, L.; Yin, J.; Wang, L.; Zhi, H.; Xu, K. Development of a Viral RdRp-Assisted Gene Silencing System and Its Application in the Identification of Host Factors of Plant (+)RNA Virus. Front. Microbiol. 2021, 12, 682921. [Google Scholar] [CrossRef]
- Sun, K.; Zhao, D.; Liu, Y.; Huang, C.; Zhang, W.; Li, Z. Rapid Construction of Complex Plant RNA Virus Infectious cDNA Clones for Agroinfection Using a Yeast-E. coli-Agrobacterium Shuttle Vector. Viruses 2017, 9, 332. [Google Scholar] [CrossRef] [Green Version]
- Yin, J.; Wang, L.; Jin, T.; Nie, Y.; Liu, H.; Qiu, Y.; Yang, Y.; Li, B.; Zhang, J.; Wang, D.; et al. A cell wall-localized NLR confers resistance to Soybean mosaic virus by recognizing viral-encoded cylindrical inclusion protein. Mol. Plant. 2021, 14, 1881–1900. [Google Scholar] [CrossRef]
- Nawaz-ul-Rehman, M.S.; Prasanth, K.R.; Xu, K.; Sasvari, Z.; Kovalev, N.; de Castro Martin, I.F.; Barajas, D.; Risco, C.; Nagy, P.D. Viral Replication Protein Inhibits Cellular Cofilin Actin Depolymerization Factor to Regulate the Actin Network and Promote Viral Replicase Assembly. PLoS Pathog. 2016, 12, e1005440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, K.; Nagy, P.D. Enrichment of Phosphatidylethanolamine in Viral Replication Compartments via Co-opting the Endosomal Rab5 Small GTPase by a Positive-Strand RNA Virus. PLoS Biol. 2016, 14, e2000128. [Google Scholar] [CrossRef] [Green Version]
- Schaad, M.C.; Jensen, P.E.; Carrington, J.C. Formation of plant RNA virus replication complexes on membranes: Role of an endoplasmic reticulum-targeted viral protein. EMBO J. 1997, 16, 4049–4059. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Wang, X.B.; Wu, Q.; Ito, T.; Cillo, F.; Li, W.X.; Chen, X.; Yu, J.L.; Ding, S.W. RNAi-mediated viral immunity requires amplification of virus-derived siRNAs in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2010, 107, 484–489. [Google Scholar] [CrossRef] [Green Version]
- Shivprasad, S.; Pogue, G.P.; Lewandowski, D.J.; Hidalgo, J.; Donson, J.; Grill, L.K.; Dawson, W.O. Heterologous sequences greatly affect foreign gene expression in tobacco mosaic virus-based vectors. Virology 1999, 255, 312–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capote, N.; Gorris, M.T.; Martinez, M.C.; Asensio, M.; Olmos, A.; Cambra, M. Interference between D and M Types of Plum pox virus in Japanese Plum Assessed by Specific Monoclonal Antibodies and Quantitative Real-Time Reverse Transcription-Polymerase Chain Reaction. Phytopathology 2006, 96, 320–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecoq, H.; Lemaire, J.M.; Wipf-Scheibel, C.J.P.D. Control of zucchini yellow mosaic virus in squash by cross protection. Plant Dis. 1991, 75, 208–211. [Google Scholar] [CrossRef]
- Valkonen, J.P.; Rajamaki, M.L.; Kekarainen, T. Mapping of viral genomic regions important in cross-protection between strains of a potyvirus. Mol. Plant Microbe Interact. 2002, 15, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Tatineni, S.; French, R. The Coat Protein and NIa Protease of Two Potyviridae Family Members Independently Confer Superinfection Exclusion. J. Virol. 2016, 90, 10886–10905. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yin, J.; Hong, X.; Luo, S.; Tan, J.; Zhang, Y.; Qiu, Y.; Latif, M.F.; Gao, T.; Yu, H.; Bai, J.; et al. The Characterization of the Tobacco-Derived Wild Tomato Mosaic Virus by Employing Its Infectious DNA Clone. Biology 2022, 11, 1467. https://doi.org/10.3390/biology11101467
Yin J, Hong X, Luo S, Tan J, Zhang Y, Qiu Y, Latif MF, Gao T, Yu H, Bai J, et al. The Characterization of the Tobacco-Derived Wild Tomato Mosaic Virus by Employing Its Infectious DNA Clone. Biology. 2022; 11(10):1467. https://doi.org/10.3390/biology11101467
Chicago/Turabian StyleYin, Jinlong, Xin Hong, Sha Luo, Jingquan Tan, Yuanming Zhang, Yanglin Qiu, Muhammad Faizan Latif, Tao Gao, Haijia Yu, Jingke Bai, and et al. 2022. "The Characterization of the Tobacco-Derived Wild Tomato Mosaic Virus by Employing Its Infectious DNA Clone" Biology 11, no. 10: 1467. https://doi.org/10.3390/biology11101467
APA StyleYin, J., Hong, X., Luo, S., Tan, J., Zhang, Y., Qiu, Y., Latif, M. F., Gao, T., Yu, H., Bai, J., Li, S., & Xu, K. (2022). The Characterization of the Tobacco-Derived Wild Tomato Mosaic Virus by Employing Its Infectious DNA Clone. Biology, 11(10), 1467. https://doi.org/10.3390/biology11101467