

Bacillus velezensis Strain GUMT319 Reshapes Soil Microbiome Biodiversity and Increases Grape Yields

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Plant Management
2.2. Physical and Chemical Properties Evaluation of the Grape
2.3. Soil Sampling
2.4. Determination of Soil Chemical Properties
2.5. 16S rDNA Sequencing and Bioinformatics
2.6. Statistical Analysis
3. Results
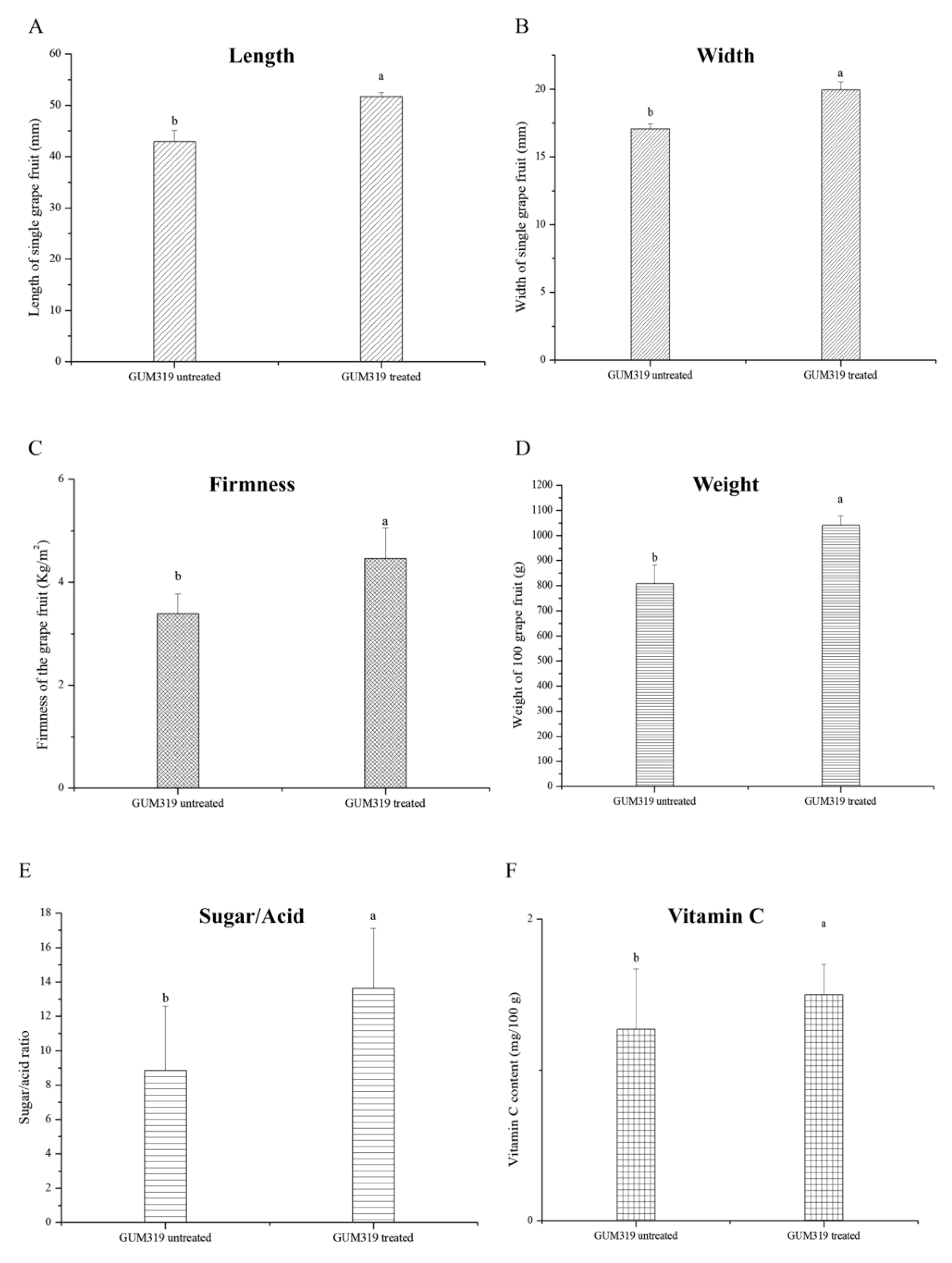
3.1. Effect of B. velezensis Strain GUMT319 on Grape Growth
3.2. Fruit Morphological and Architectural Parameters
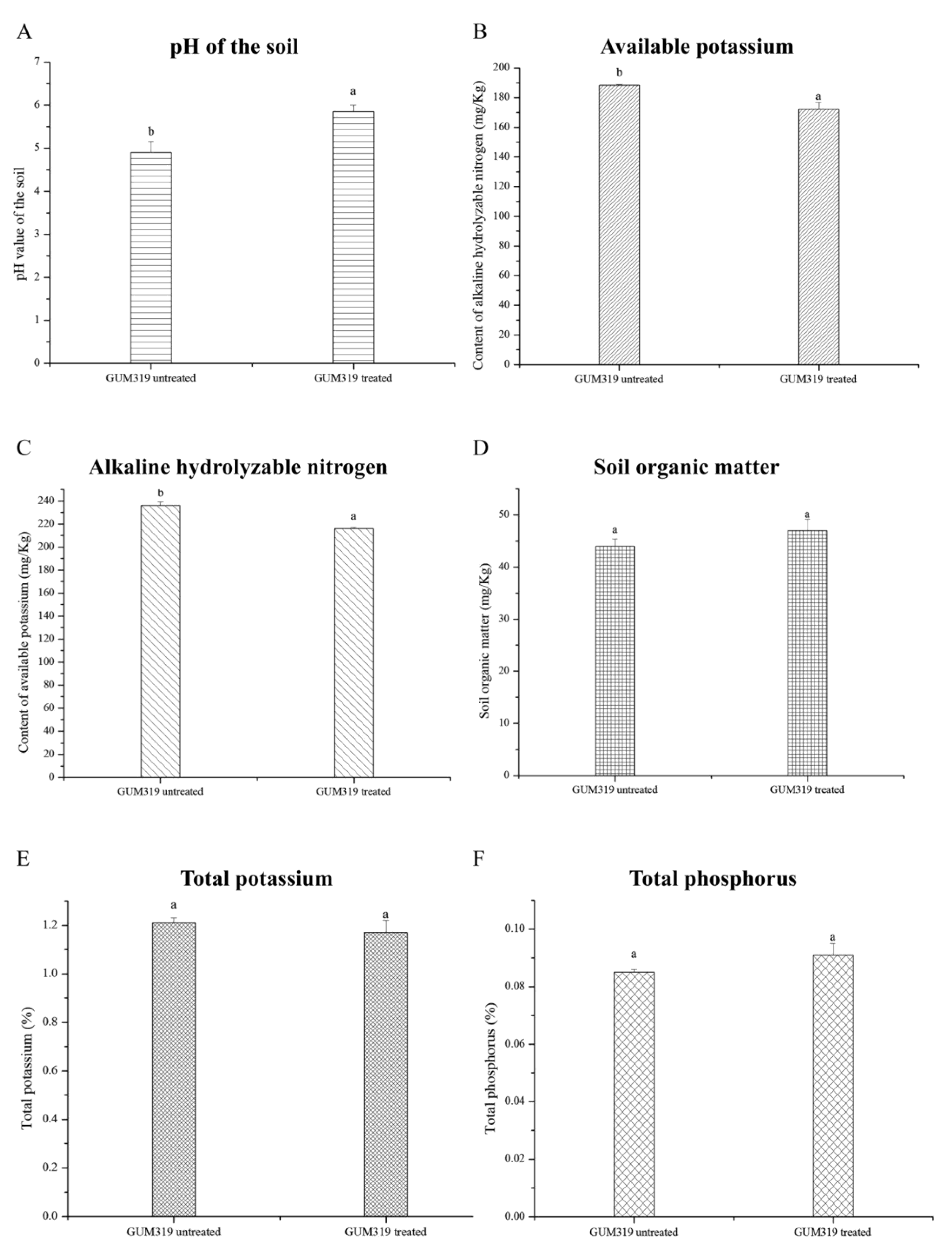
3.3. Changes in Soil Chemical Properties
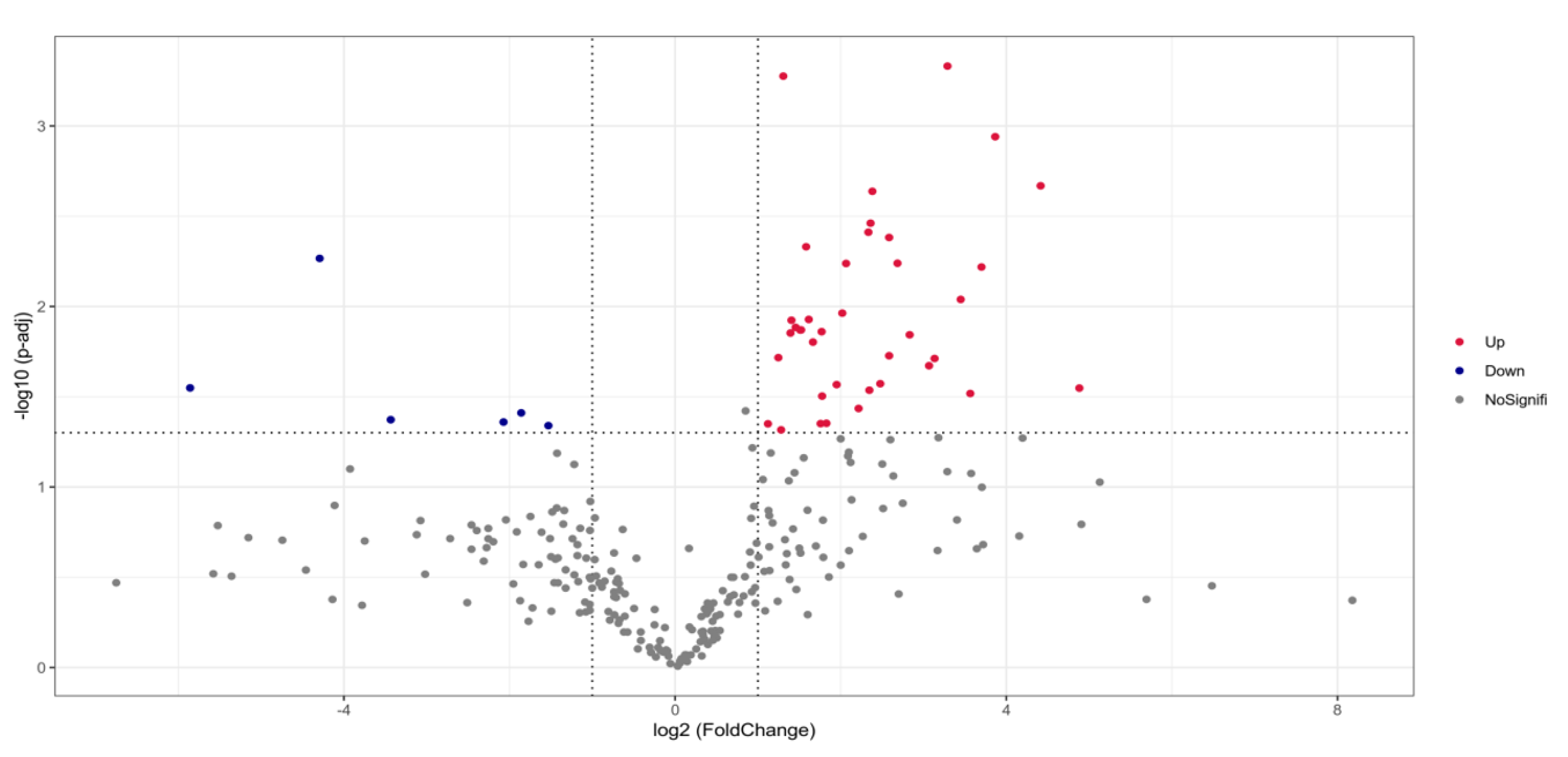
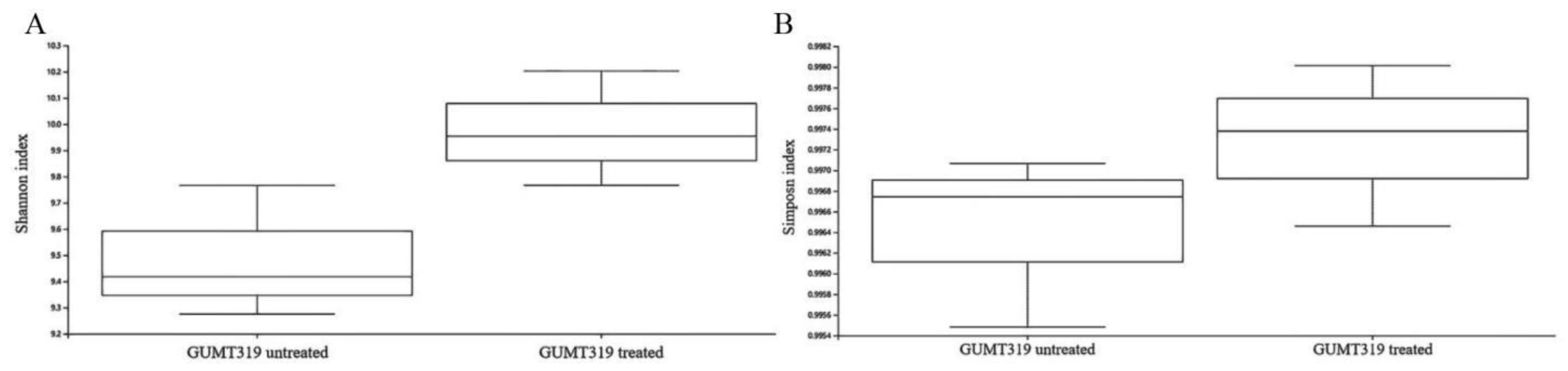
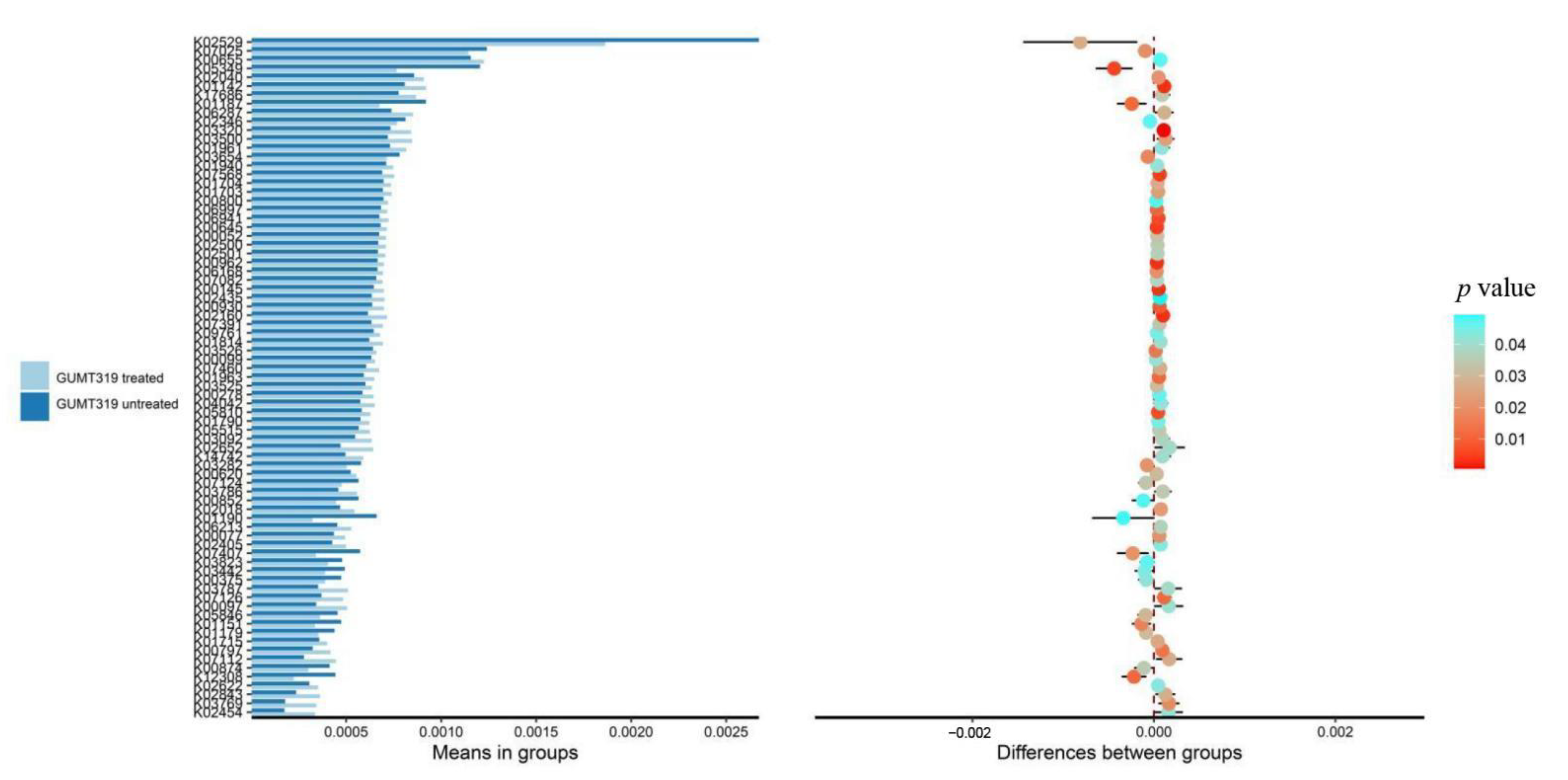
3.4. Effect of GUMT319 on the Soil Microbial Community Composition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Saeed, Q.; Xiukang, W.; Haider, F.U.; Kučerik, J.; Mumtaz, M.Z.; Holatko, J.; Naseem, M.; Kintl, A.; Ejaz, M.; Naveed, M.; et al. Rhizosphere Bacteria in Plant Growth Promotion, Biocontrol, and Bioremediation of Contaminated Sites: A Comprehensive Review of Effects and Mechanisms. Int. J. Mol. Sci. 2021, 22, 10529. [Google Scholar] [CrossRef] [PubMed]
- Peer, R.V. Induced Resistance and Phytoalexin Accumulation in Biological Control of Fusarium Wilt of Carnation by Pseudomonas sp. Strain WCS417r. Phytopathology 1991, 81, 728–734. [Google Scholar] [CrossRef]
- Wei, G. Induction of systemic resistance of cucumber to Colletotrichum orbiculare by select strains of plant growth-promoting rhizobacteria. Phytopathology 1991, 81, 1508–1512. [Google Scholar] [CrossRef]
- Khan, M.R.; Fischer, S.; Egan, D.; Doohan, F.M. Biological control of fusarium seedling blight disease of wheat and barley. Phytopathology 2006, 96, 386–394. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.C.; Cheung, K.C.; Luo, Y.M.; Wong, M.H. Effects of inoculation of plant growth-promoting rhizobacteria on metal uptake by Brassica juncea. Environ. Pollut. 2006, 140, 124–135. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, M.; Valderrama, B.; Arredondo-Peter, R.; Soberón, M.; Mora, J.; Hernández, G. Rhizobium etli Genetically Engineered for the Heterologous Expression of Vitreoscilla sp. Hemoglobin: Effects on Free-Living and Symbiosis. Mol. Plant-Microbe Interact. 1999, 12, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Elkhatib, H.A.; Gabr, S.M.; Mohamed, R.A.; Manaa, A.F. Impact of potassium fertilizer rates and potassium solubilizing bacteria inoculation on the growth, yield and quality of potato (Solanum tuberosum L.). Alex. Sci. Exch. J. 2019, 40, 127–142. [Google Scholar] [CrossRef]
- Sah, S.; Krishnani, S.; Singh, R. Pseudomonas mediated nutritional and growth promotional activities for sustainable food security. Curr. Res. Microb. Sci. 2021, 2, 100084. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.; Feng, S.; Wang, J.; Lu, W.; Zhang, W.; Chen, M.; Lin, M. Effect of inoculation with nitrogen-fixing bacterium Pseudomonas stutzeri A1501 on maize plant growth and the microbiome indigenous to the rhizosphere. Syst. Appl. Microbiol. 2019, 42, 248–260. [Google Scholar] [CrossRef] [PubMed]
- Kalayu, G. Phosphate Solubilizing Microorganisms: Promising Approach as Biofertilizers. Int. J. Agron. 2019, 2019, 4917256. [Google Scholar] [CrossRef]
- Sheng, X.F. Growth promotion and increased potassium uptake of cotton and rape by a potassium releasing strain of Bacillus edaphicus. Soil Biol. Biochem. 2005, 37, 1918–1922. [Google Scholar] [CrossRef]
- Saha, M.; Sarkar, S.; Sarkar, B.; Sharma, B.K.; Bhattacharjee, S.; Tribedi, P. Microbial siderophores and their potential applications: A review. Environ. Sci. Pollut. Res. 2016, 23, 3984–3999. [Google Scholar] [CrossRef] [PubMed]
- Subramanium, N.; Sundaram, L. Siderophore producing Pseudomonas spp. isolated from rhizospheric soil and enhancing iron content in Arachis hypogaea L. Plant. J. Agric. Technol. 2020, 16, 429–442. [Google Scholar]
- Tewari, S.; Arora, N.K. Role of salicylic acid from Pseudomonas aeruginosa PF23EPS+ in growth promotion of sunflower in saline soils infested with phytopathogen Macrophomina phaseolina. Environ. Sustain. 2018, 1, 49–59. [Google Scholar] [CrossRef]
- Hakim, S.; Naqqash, T.; Nawaz, M.S.; Laraib, I.; Siddique, M.J.; Zia, R.; Mirza, M.S.; Imran, A. Rhizosphere Engineering with Plant Growth-Promoting Microorganisms for Agriculture and Ecological Sustainability. Front. Sustain. Food Syst. 2021, 5, 617157. [Google Scholar] [CrossRef]
- Aloo, B.N.; Makumba, B.A.; Mbega, E.R. The potential of Bacilli rhizobacteria for sustainable crop production and environmental sustainability. Microbiol. Res. 2019, 219, 26–39. [Google Scholar] [CrossRef]
- Saxena, A.K.; Kumar, M.; Chakdar, H.; Anuroopa, N.; Bagyaraj, D.J. Bacillus species in soil as a natural resource for plant health and nutrition. J. Appl. Microbiol. 2020, 128, 1583–1594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Chen, L.; Wu, G.; Feng, H.; Zhang, G.; Shen, Q.; Zhang, R. Identification of Root-Secreted Compounds Involved in the Communication Between Cucumber, the Beneficial Bacillus amyloliquefaciens, and the Soil-Borne Pathogen Fusarium oxysporum. Mol. Plant Microbe. Interact. 2017, 30, 53–62. [Google Scholar] [CrossRef] [Green Version]
- Feng, H.; Zhang, N.; Fu, R.; Liu, Y.; Krell, T.; Du, W.; Shao, J.; Shen, Q.; Zhang, R. Recognition of dominant attractants by key chemoreceptors mediates recruitment of plant growth-promoting rhizobacteria. Environ. Microbiol. 2019, 21, 402–415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhury, S.P.; Hartmann, A.; Gao, X.; Borriss, R. Biocontrol mechanism by root-associated Bacillus amyloliquefaciens FZB42—A review. Front. Microbiol. 2015, 6, 780. [Google Scholar] [CrossRef] [Green Version]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491, Erratum in Front. Microbiol. 2019, 10, 1279. [Google Scholar] [CrossRef] [PubMed]
- Nazari, M.; Smith, D.L. A PGPR-Produced Bacteriocin for Sustainable Agriculture: A Review of Thuricin 17 Characteristics and Applications. Front. Plant Sci. 2020, 11, 916. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.; Tu, C.; Xie, Z.; Luo, Y.; Zhang, L.; Li, Z. The genome of Bacillus velezensis SC60 provides evidence for its plant probiotic effects. Microorganisms 2022, 10, 767. [Google Scholar] [CrossRef] [PubMed]
- Besset-Manzoni, Y.; Rieusset, L.; Joly, P.; Comte, G.; Prigent-Combaret, C. Exploiting rhizosphere microbial cooperation for developing sustainable agriculture strategies. Environ. Sci. Pollut. Res. 2018, 25, 29953–29970. [Google Scholar] [CrossRef] [PubMed]
- Gouda, S.; Kerry, R.G.; Das, G.; Paramithiotis, S.; Shin, H.-S.; Patra, J.K. Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 2018, 206, 131–140. [Google Scholar] [CrossRef]
- Behera, S.S.; Ray, R.C. Bioprospecting of cowdung microflora for sustainable agricultural, biotechnological and environmental applications. Curr. Res. Microb. Sci. 2020, 2, 100018. [Google Scholar] [CrossRef]
- Haskett, T.L.; Tkacz, A.; Poole, P.S. Engineering rhizobacteria for sustainable agriculture. ISME J. 2021, 15, 949–964. [Google Scholar] [CrossRef]
- Ding, H.; Mo, W.; Yu, S.; Cheng, H.; Peng, L.; Liu, Z. Whole genome sequence of Bacillus velezensis strain GUMT319: A potential biocontrol agent against tobacco black shank disease. Front. Microbiol. 2021, 12, 658113. [Google Scholar] [CrossRef]
- Van Reeuwijk, L.P. Procedure for Soil Analysis, 4th ed.; International Soil Reference Center Wageningen (ISRIC): Wageningen, The Netherlands, 1993. [Google Scholar]
- Li, Y.; Niu, W.; Dyck, M.; Wang, J.; Zou, X. Yields and nutritional of greenhouse tomato in response to different soil aeration volume at two depths of subsurface drip irrigation. Sci. Rep. 2016, 6, 39307. [Google Scholar] [CrossRef] [Green Version]
- Lv, J.; Sun, M.; Zhang, Y.; Chen, J.; Ge, Y.; Li, J. Comparative analysis of expression profiles of metacaspase (MC) genes between two apple (Malus domestica) cultivars with distinct ripening behavior. J. Food Biochem. 2022, 00, e14400. [Google Scholar] [CrossRef]
- Sayed Elnenaey, E.; Soliman, R. A sensitive colorimetric method for estimation of ascorbic acid. Talanta 1979, 26, 1164–1166. [Google Scholar] [CrossRef]
- Song, J.; Bi, J.; Chen, Q.; Wu, X.; Lyu, Y.; Meng, X. Assessment of sugar content, fatty acids, free amino acids, and volatile profiles in jujube fruits at different ripening stages. Food Chem. 2019, 270, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Qi, L.; Wang, B.; Medley, P.; Drake, J.; Vernon, J.; Ibeanusi, V.; Chen, G. Assess long-term As, Pb and Cr contamination and uptake by Eriocaulon decangulare in the Apalachicola National Forest. Sci. Total Environ. 2022, 838, 156040. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Ingram, J.S.I. Tropical soil biology and fertility: A handbook of methods C.A.B.; Oxford University Press: Oxford, UK, 1993; 221p. [Google Scholar]
- Hoang, H.T.; Le, D.H.; Le, T.T.H.; Nguyen, T.T.N.; Chu, H.H.; Nguyen, N.T. Metagenomic 16S rDNA amplicon data of microbial diversity of guts in Vietnamese humans with type 2 diabetes and nondiabetic adults. Data Brief 2020, 34, 106690. [Google Scholar] [CrossRef]
- An, N.; Wang, C.; Dou, X.; Liu, X.; Wu, J.; Cheng, Y. Comparison of 16S rDNA Amplicon Sequencing With the Culture Method for Diagnosing Causative Pathogens in Bacterial Corneal Infections. Transl. Vis. Sci. Technol. 2022, 11, 29. [Google Scholar] [CrossRef]
- Callahan, B.J.; Wong, J.; Heiner, C.; Oh, S.; Theriot, C.M.; Gulati, A.S.; McGill, S.K.; Dougherty, M.K. High-throughput amplicon sequencing of the full-length 16S rRNA gene with single-nucleotide resolution. Nucleic Acids Res. 2019, 47, e103. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Gregory Caporaso, J. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Li, B.; Zhang, X.; Guo, F.; Wu, W.; Zhang, T. Characterization of tetracycline resistant bacterial community in saline activated sludge using batch stress incubation with high-throughput sequencing analysis. Water Res. 2013, 47, 4207–4216. [Google Scholar] [CrossRef]
- Avershina, E.; Frisli, T.; Rudi, K. De novo Semi-alignment of 16S rRNA Gene Sequences for Deep Phylogenetic Characterization of Next Generation Sequencing Data. Microbes Environ. 2013, 28, 211–216. [Google Scholar] [CrossRef] [Green Version]
- Kruskal, J.B. Nonmetric multidimensional scaling: A numerical method. Psychometrika 1964, 29, 115–129. [Google Scholar] [CrossRef]
- Xu, Z.; Mandic-Mulec, I.; Zhang, H.; Liu, Y.; Sun, X.; Feng, H.; Xun, W.; Zhang, N.; Shen, Q.; Zhang, R. Antibiotic Bacillomycin D Affects Iron Acquisition and Biofilm Formation in Bacillus velezensis through a Btr-Mediated FeuABC-Dependent Pathway. Cell Rep. 2019, 29, 1192–1202.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adeleke, B.S.; Ayangbenro, A.S.; Babalola, O.O. Genomic Analysis of Endophytic Bacillus cereus T4S and Its Plant Growth-Promoting Traits. Plants 2021, 10, 1776. [Google Scholar] [CrossRef] [PubMed]
- Bharti, N.; Barnawal, D. Amelioration of salinity stress by PGPR: ACC deaminase and ROS scavenging enzymes activity. In PGPR Amelioration in Sustainable Agriculture; Singh, A.K., Kumar, A., Singh, P.K., Eds.; Woodhead Publishing: Cambridge, UK, 2019; pp. 85–106. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Steenkamp, P.A.; Labuschagne, N.; Dubery, I.A. Metabolic Profiling of PGPR-Treated Tomato Plants Reveal Priming-Related Adaptations of Secondary Metabolites and Aromatic Amino Acids. Metabolites 2020, 10, 210. [Google Scholar] [CrossRef]
- Mantzoukas, S.; Lagogiannis, I.; Mpousia, D.; Ntoukas, A.; Karmakolia, K.; Eliopoulos, P.A.; Poulas, K. Beauveria bassiana Endophytic Strain as Plant Growth Promoter: The Case of the Grape Vine Vitis vinifera. J. Fungi 2021, 7, 142. [Google Scholar] [CrossRef]
- Freschet, G.T.; Pagès, L.; Iversen, C.M.; Comas, L.H.; Rewald, B.; Roumet, C.; Klimešová, J.; Zadworny, M.; Poorter, H.; Postma, J.A.; et al. A starting guide to root ecology: Strengthening ecological concepts and standardising root classification, sampling, processing and trait measurements. N. Phytol. 2021, 232, 973–1122, Erratum in N. Phytol. 2022, 235, 372. [Google Scholar] [CrossRef]
- Malik, A.A.; Puissant, J.; Buckeridge, K.M.; Goodall, T.; Jehmlich, N.; Chowdhury, S.; Gweon, H.S.; Peyton, J.M.; Mason, K.E.; van Agtmaal, M.; et al. Land use driven change in soil pH affects microbial carbon cycling processes. Nat. Commun. 2018, 9, 3591. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, X.; Yang, F.; Bai, C.; Shi, Q.; Hu, S.; Tang, X.; Peng, L.; Ding, H. Bacillus velezensis Strain GUMT319 Reshapes Soil Microbiome Biodiversity and Increases Grape Yields. Biology 2022, 11, 1486. https://doi.org/10.3390/biology11101486
Chen X, Yang F, Bai C, Shi Q, Hu S, Tang X, Peng L, Ding H. Bacillus velezensis Strain GUMT319 Reshapes Soil Microbiome Biodiversity and Increases Grape Yields. Biology. 2022; 11(10):1486. https://doi.org/10.3390/biology11101486
Chicago/Turabian StyleChen, Xiangru, Fang Yang, Chunwei Bai, Qianrui Shi, Shan Hu, Xianying Tang, Lijuan Peng, and Haixia Ding. 2022. "Bacillus velezensis Strain GUMT319 Reshapes Soil Microbiome Biodiversity and Increases Grape Yields" Biology 11, no. 10: 1486. https://doi.org/10.3390/biology11101486
APA StyleChen, X., Yang, F., Bai, C., Shi, Q., Hu, S., Tang, X., Peng, L., & Ding, H. (2022). Bacillus velezensis Strain GUMT319 Reshapes Soil Microbiome Biodiversity and Increases Grape Yields. Biology, 11(10), 1486. https://doi.org/10.3390/biology11101486