
Eating More and Fighting Less: Social Foraging Is a Potential Advantage for Successful Expansion of Bird Source Populations
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Husbandry
2.2. Procedures
2.2.1. Novel Food Preparation
2.2.2. Experiment 1: Solitary Foraging of Novel Food
2.2.3. Experiment 2: Group Foraging and Competition for Novel Food
2.3. Behavioral Measurements
2.4. Statistical Analyses
3. Results
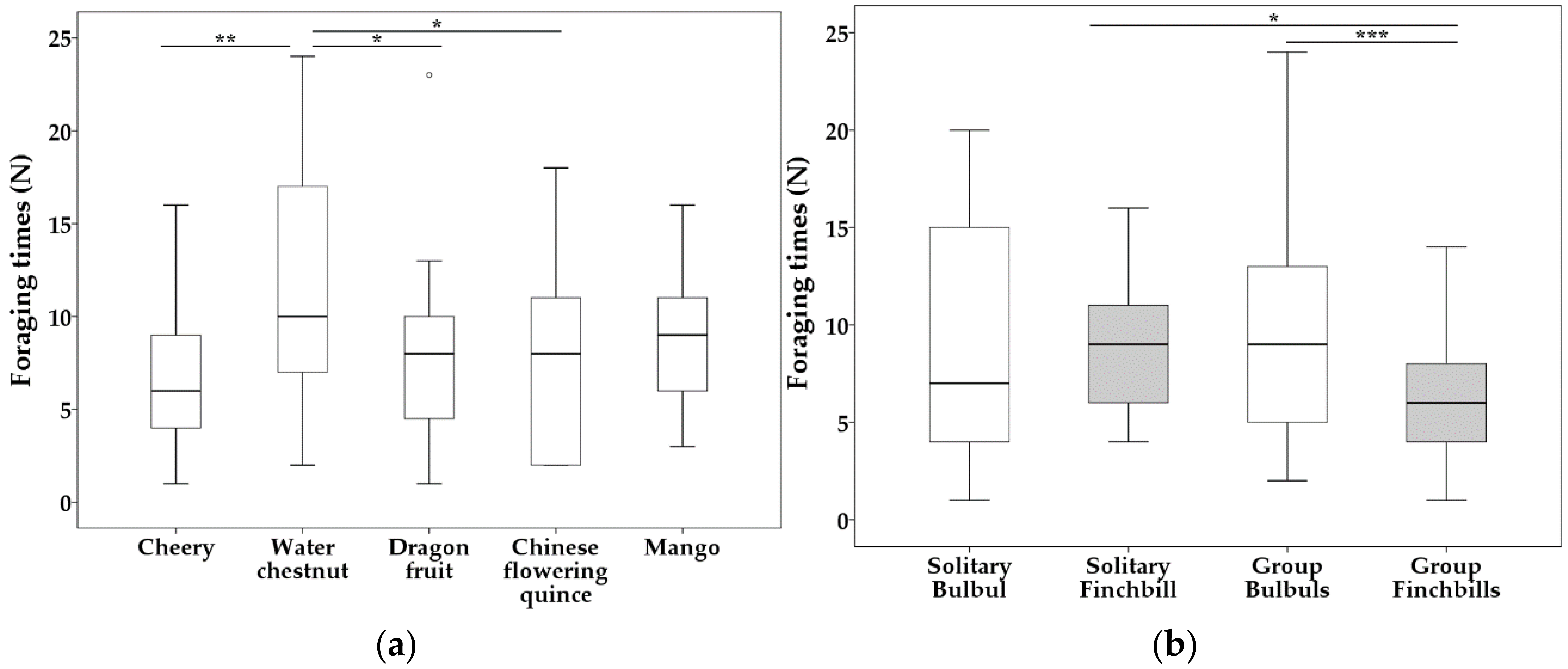
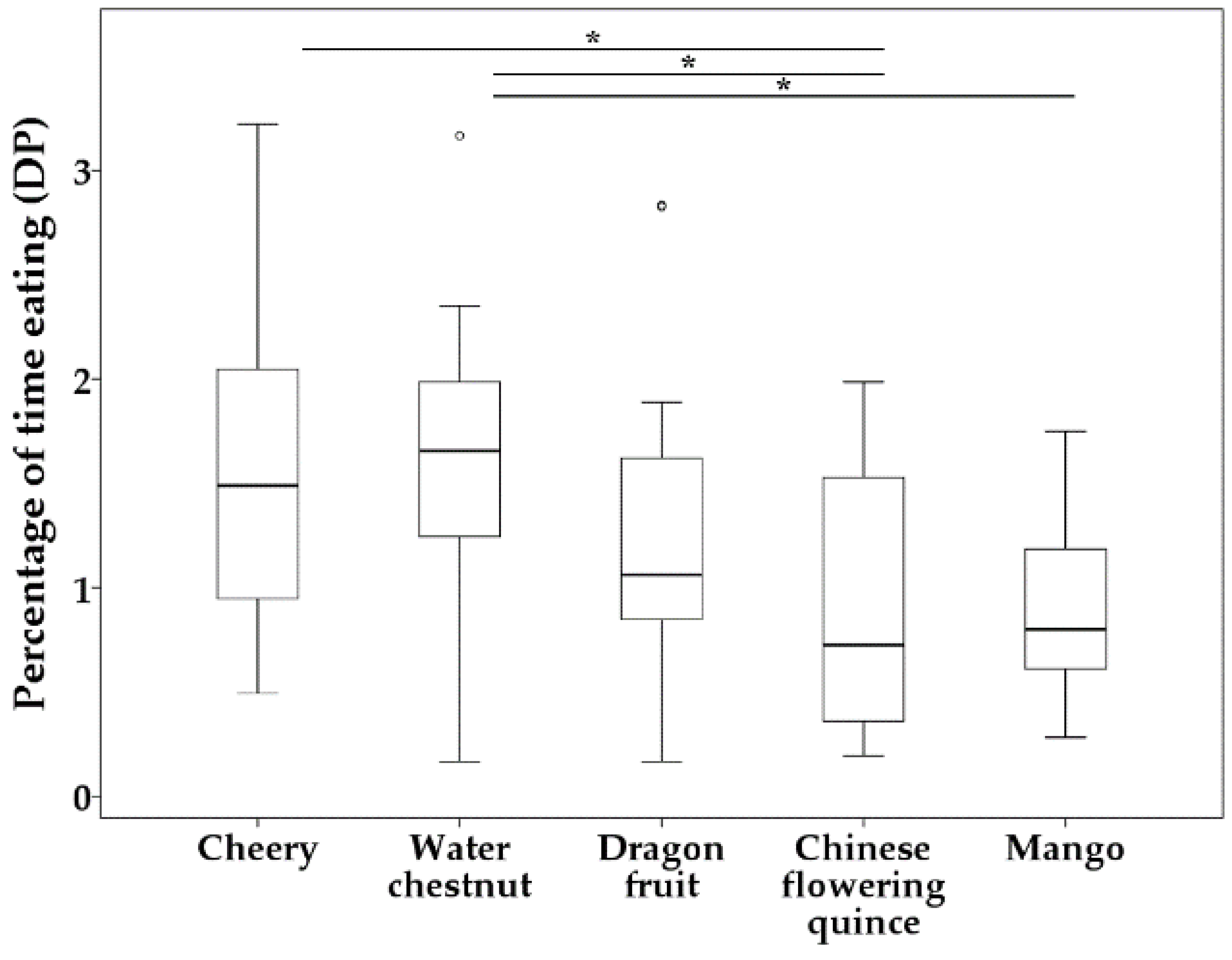
3.1. Variations in Foraging Behavior among Solitary and Group Bulbuls and Finchbills
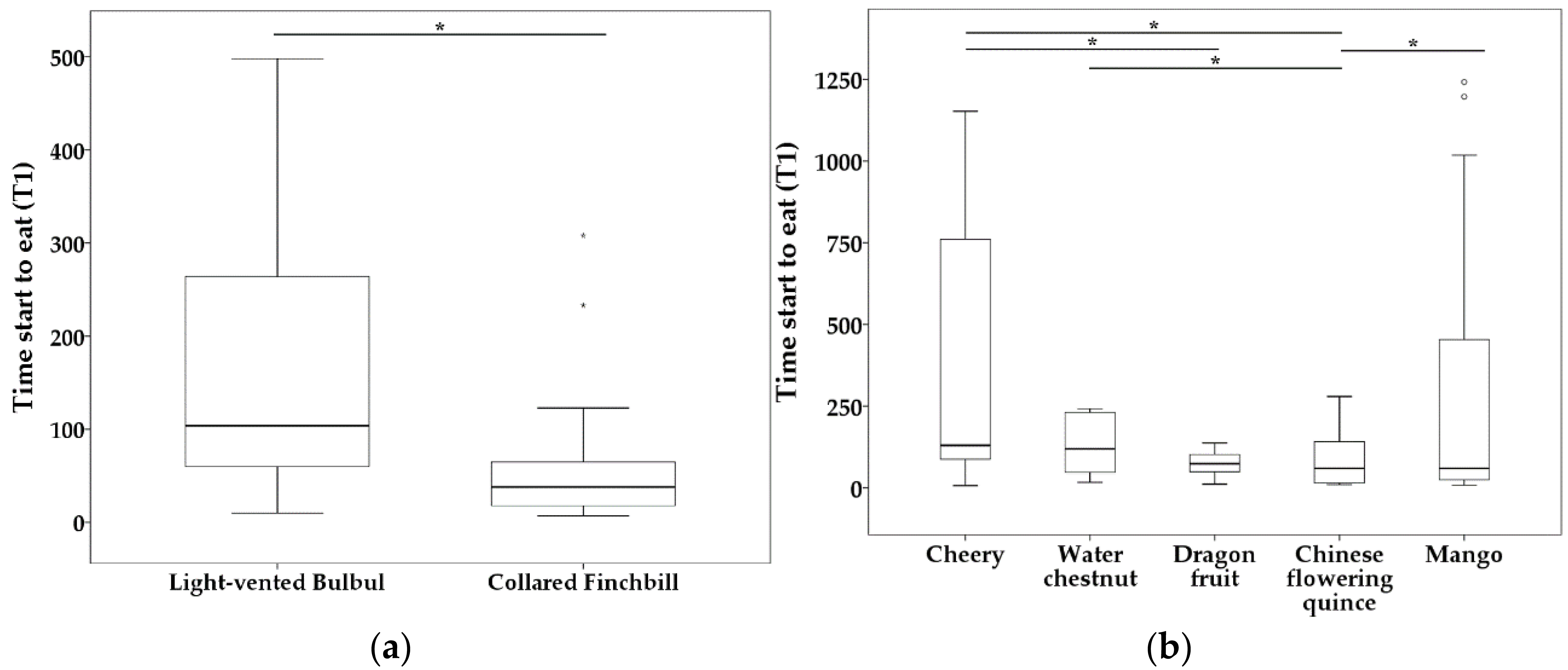
3.2. Comparison of Foraging Behavior between Bulbuls and Finchbills
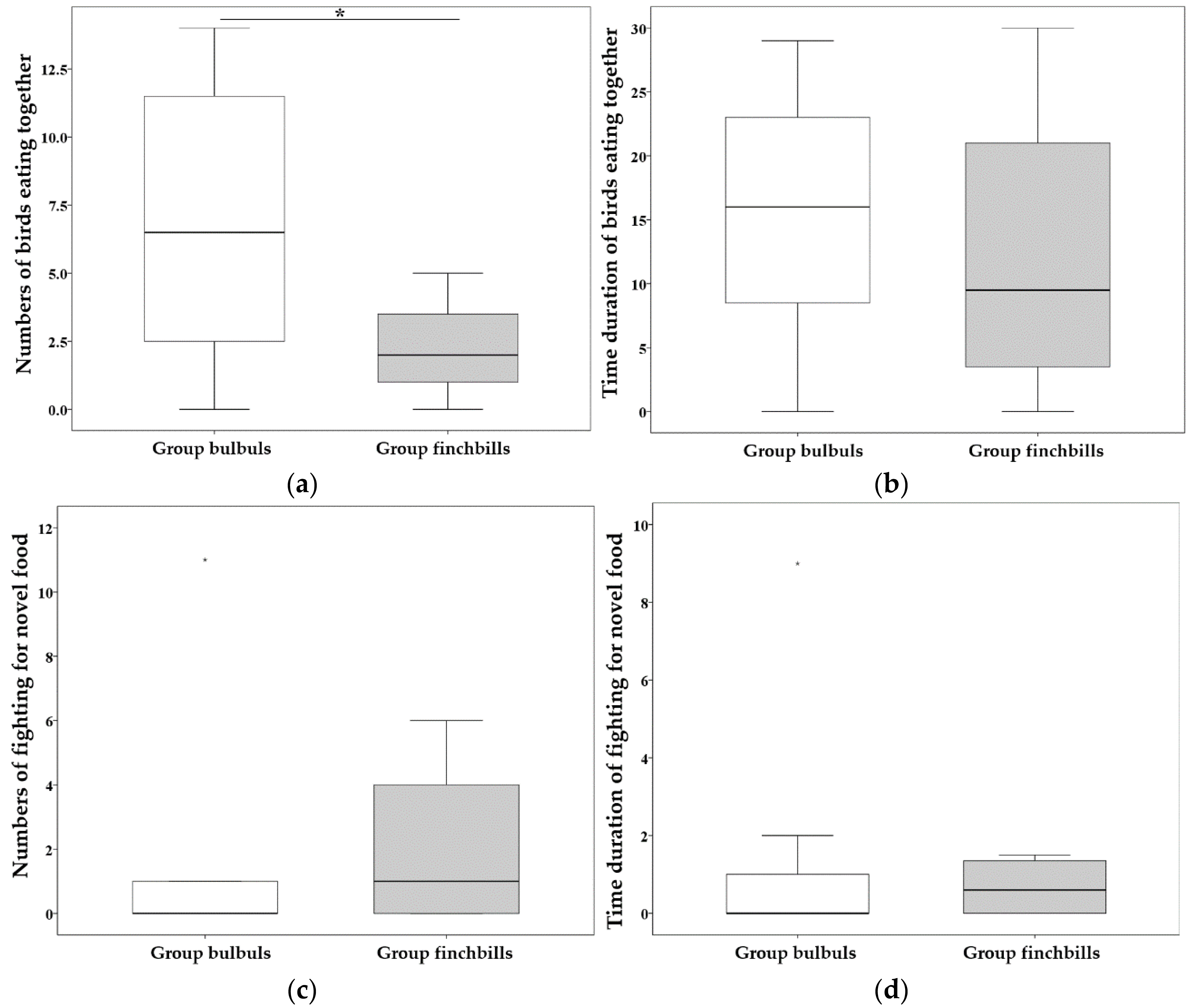
3.3. Changes from Solitary to Group Context
3.4. Intraspecific Food Competition
4. Discussion
4.1. Finchbills Foraged More Novel Food in a Solitary Than Social Context
4.2. Bulbuls Consumed More Novel Food after Transferring from Solitary to Group Foraging
4.3. Bulbuls Shared More but Competed Less in Social Context
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burger, C.; Nord, A.; Nilsson, J.-Å.; Gilot-Fromont, E.; Both, C. Fitness consequences of northward dispersal as possible adaptation to climate change, using experimental translocation of a migratory passerine. PLoS ONE 2013, 8, e83176. [Google Scholar]
- Kamp, J.; Oppel, S.; Ananin, A.A.; Durnev, Y.A.; Gashev, S.N.; Hölzel, N.; Mishchenko, A.L.; Pessa, J.; Smirenski, S.M.; Strelnikov, E.G. Global population collapse in a superabundant migratory bird and illegal trapping in China. Conserv. Biol. 2015, 29, 1684–1694. [Google Scholar] [CrossRef] [PubMed]
- Holway, D.A.; Suarez, A.V. Animal behavior: An essential component of invasion biology. Trends Ecol. Evol. 1999, 14, 328–330. [Google Scholar] [CrossRef]
- Kolar, C.S.; Lodge, D.M. Progress in invasion biology: Predicting invaders. Trends Ecol. Evol. 2001, 16, 199–204. [Google Scholar] [CrossRef]
- Wright, T.F.; Eberhard, J.R.; Hobson, E.A.; Avery, M.L.; Russello, M.A. Behavioral flexibility and species invasions: The adaptive flexibility hypothesis. Ethol. Ecol. Evol. 2010, 22, 393–404. [Google Scholar] [CrossRef]
- Chapple, D.G.; Simmonds, S.M.; Wong, B.B.M. Can behavioral and personality traits influence the success of unintentional species introductions? Trends Ecol. Evol. 2012, 27, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Machovsky-Capuska, G.E.; Senior, A.M.; Zantis, S.P.; Barna, K.; Cowieson, A.J.; Pandya, S.; Pavard, C.; Shiels, M.; Raubenheimer, D. Dietary protein selection in a free-ranging urban population of common myna birds. Behav. Ecol. 2016, 27, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Wood-Gush, D.; Vestergaard, K. The seeking of novelty and its relation to play. Anim. Behav. 1991, 42, 599–606. [Google Scholar] [CrossRef]
- Wood-Gush, D.G.; Vestergaard, K. Inquisitive exploration in pigs. Anim. Behav. 1993, 45, 185–187. [Google Scholar] [CrossRef]
- Mettke-Hofmann, C.; Winkler, H.; Leisler, B. The significance of ecological factors for exploration and neophobia in parrots. Ethology 2002, 108, 249–272. [Google Scholar] [CrossRef]
- Marples, N.M.; Mappes, J. Can the dietary conservatism of predators compensate for positive frequency dependent selection against rare, conspicuous prey? Evol. Ecol. 2011, 25, 737–749. [Google Scholar] [CrossRef]
- Martin, L.B.; Lisa, F. A taste for novelty in invading house sparrows. Passer Domesticus. Behav. Ecol. 2005, 16, 702–707. [Google Scholar] [CrossRef]
- Duncan, R.P.; Sol, B.D. The ecology of bird introductions. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 71–98. [Google Scholar] [CrossRef]
- Romanuk, T.N.; Zhou, Y.; Brose, U.; Berlow, E.L.; Williams, R.J.; Martinez, N.D. Predicting invasion success in complex ecological networks. Philos. Trans. R Soc. Lond. B Biol. Sci. 2019, 364, 1743–1754. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuliozi, B.; Fracasso, G.; Hoi, H.; Griggio, M. House sparrows’ (Passer domesticus) behaviour in a novel environment is modulated by social context and familiarity in a sex-specific manner. Front. Zool. 2018, 15, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Oers, K.; Klunder, M.; Drent, P.J. Context dependence of personalities: Risk-taking behavior in a social and a nonsocial situation. Behav. Ecol. 2005, 16, 716–723. [Google Scholar] [CrossRef] [Green Version]
- Hofmann, H.A.; Beery, A.K.; Blumstein, D.T.; Couzin, I.D.; Earley, R.L.; Hayes, L.D.; Hurd, P.L.; Lacey, E.A.; Phelps, S.M.; Solomon, N.G. An evolutionary framework for studying mechanisms of social behavior. Trends Ecol. Evol. 2014, 29, 581–589. [Google Scholar] [CrossRef] [Green Version]
- Jones, T.B.; Aplin, L.M.; Devost, I.; Morand-Ferron, J. Individual and ecological determinants of social information transmission in the wild. Anim. Behav. 2017, 129, 93–101. [Google Scholar] [CrossRef]
- Vasquez, R.A.; Kacelnik, A. Foraging rate versus sociality in the starling Sturnus vulgaris. Proc. Royal Soc. B 2000, 267, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Liker, A.; Bokony, V. Larger groups are more successful in innovative problem solving in house sparrows. Proc. Natl. Acad. Sci. USA 2009, 106, 7893–7898. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, A.; Hausberger, M.; Clergeau, P. Flexibility in European starlings’ use of social information: Experiments with decoys in different populations. Anim. Behav. 2010, 80, 965–973. [Google Scholar] [CrossRef]
- Vall-Llosera, M.; Llimona, F.; de Cáceres, M.; Sales, S.; Sol, D. Competition, niche opportunities and the successful invasion of natural habitats. Biol. Invasions 2016, 18, 1–12. [Google Scholar] [CrossRef]
- Podulka, S.; Rohrbaugh, R.W.; Bonney, R. Handbook of Bird Biology; Princeton University Press: Princeton, NJ, USA, 2004; p. 296. [Google Scholar]
- Gruber, J.; Brown, G.; Whiting, M.J.; Shine, R. Behavioural divergence during biological invasions: A study of cane toads (Rhinella marina) from contrasting environments in Hawaii. Royal Soc. Open Sci. 2018, 5, 180197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, J.; Brown, G.; Whiting, M.J.; Shine, R. Is the behavioural divergence between range-core and range-edge populations of cane toads (Rhinella marina) due to evolutionary change or developmental plasticity? Royal Soc. Open Sci. 2017, 4, 170789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gruber, J.; Brown, G.; Whiting, M.J.; Shine, R. Geographic divergence in dispersal-related behaviour in cane toads from range-front versus range-core populations in Australia. Behav. Ecol. Sociobiol. (Print) 2017, 71, 1–7. [Google Scholar] [CrossRef]
- Chuang, A.; Peterson, C.R. Expanding population edges: Theories, traits, and trade-offs. Glob. Chang. Biol. 2016, 22, 494–512. [Google Scholar] [CrossRef]
- Lapiedra, O.; Chejanovski, Z.; Kolbe, J.J. Urbanization and biological invasion shape animal personalities. Glob. Chang. Biol. 2017, 23, 592–603. [Google Scholar] [CrossRef]
- The CBR Checklist of Birds of China v5.0. China Bird Report, 16 November 2017.
- Xu, X.Y.; Lin, Y.; Yi, J.F.; Zhou, F.; Yu, L.J. Distribution change of red-whiskered bulbul (Pycnonotus jocosus) in China over past 40 years. Chinese J. Zool. 2018, 53, 17–25. [Google Scholar]
- Xing, X.Y.; Alstrom, P.; Yang, X.J.; Lei, F.M. Recent northward range expansion promotes song evolution in a passerine bird, the light-vented bulbul. J. Evol. Biol. 2013, 26, 867–877. [Google Scholar] [CrossRef]
- Wang, X.P.; Du, M.; Sun, L.X.; Li, J.L. New distribution of Chinese bulbuls (Pycnonotus sinensis) in Lüshun of Dalian, China. Chinese Zool. Res. 2005, 26, 95. [Google Scholar]
- Piao, J.A.; Piao, J.; Jiang, T.Y. Bird survey of Light-vented bulbul in Dalian, China. Chinese J. Liaoning Univ. 2012, 39, 361–364. [Google Scholar]
- Liu, J.; Cheng, H.Y.; Bai, C.; Zhan, T.T.; Zhang, J.; Wang, Y.; Liu, H.Y.; Mao, Y.; Cui, D.Y. New distribution record of collared finchbill (Spizixos semitorques) in Beijing city. Chinese J. Wildl. 2022, 43, 2. [Google Scholar]
- Fan, J.G.; Wang, P.H.; Hou, J.H. Collared finchbill Spizixos semitorques found in Xingtai city, Hebei province. Chinese J. Zool. 2020, 55, 1. [Google Scholar]
- Del Hoyo, J.; Elliott, A.; Christie, D.A. Handbook of the Birds of the World. Volume 10. Cuckoo-Shrikes to Thrushes; Lynx Edicions: Barcelona, Spain, 2005; pp. 149–151. [Google Scholar]
- Cassey, P. Life history and ecology influences establishment success of introduced land birds. Biol. J. Linn. Soc. Lond. 2002, 76, 465–480. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, T.M.; Cassey, P.; Lockwood, J.L. The role of species traits in the establishment success of exotic birds. Glob. Chang. Biol. 2010, 15, 2852–2860. [Google Scholar] [CrossRef]
- Menzel, E.W.; Menzel, A. Cognitive, Developmental and social aspects of responsiveness to novel objects in a Family group of Marmosets (Saguinus fuscicollis). Behaviour 1979, 70, 251–279. [Google Scholar] [CrossRef]
- Podsakoff, P.M.; Mackenzie, S.B.; Lee, J.Y.; Podsakoff, N.P. Common method biases in behavioral research: A critical review of the literature and recommended remedies. J. Appl. Psychol. 2003, 88, 879–903. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2014. [Google Scholar]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 8, 1–27. [Google Scholar]
- Peneaux, C.; Machovsky-Capuska, G.E.; Raubenheimer, D.; Lermite, F.; Rousseau, C.; Ruhan, T.; Rodger, J.C.; Griffin, A.S. Tasting novel foods and selecting nutrient content in a highly successful ecological invader, the common myna. J. Avian Biol. 2017, 48, 1432–1440. [Google Scholar] [CrossRef]
- Levine, J.M. Biological invasions. Curr. Biol. 2008, 18, R57–R60. [Google Scholar] [CrossRef] [Green Version]
- Cavalli, M.; Baladrón, A.V.; Isacch, J.P.; Martínez, G.; Bó, M.S. Prey selection and food habits of breeding burrowing owls (Athene cunicularia) in natural and modified habitats of Argentine pampas. Emu 2014, 114, 184–188. [Google Scholar] [CrossRef]
- Coleman, S.L.; Mellgren, R.L. Neophobia when feeding alone or in flocks in zebra finches, Taeniopygia guttata. Anim. Behav. 1994, 48, 903–907. [Google Scholar] [CrossRef]
- Stöwe, M.; Bugnyar, T.; Loretto, M.C.; Schloegl, C.; Range, F.; Kotrschal, K. Novel object exploration in ravens (Corvus corax): Effects of social relationships. Behav. Process. 2006, 73, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Stöwe, M.; Bugnyar, T.; Heinrich, B.; Kotrschal, K. Effects of group size on approach to novel objects in ravens (Corvus corax). Ethology 2006, 112, 1079–1088. [Google Scholar] [CrossRef]
- Aplin, L.M.; Farine, D.R.; Morand-Ferron, J.; Cockburn, A.; Thornton, A.; Sheldon, B.C. Experimentally induced innovations lead to persistent culture via conformity in wild birds. Nature 2015, 518, 538–541. [Google Scholar] [CrossRef] [Green Version]
- Stöwe, M.; Kotrschal, K. Behavioural phenotypes may determine whether social context facilitates or delays novel object exploration in ravens (Corvus corax). J. Ornithol. 2007, 148, 179–184. [Google Scholar] [CrossRef]
- Pereira, P.F.; Godinho, C.; Vila-Viçosa, M.J.O.; Mota, P.G.; Lourenço, R. Competitive advantages of the red-billed leiothrix (Leiothrix lutea) invading a passerine community in Europe. Biol. Invasions 2017, 19, 1–10. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fixed Effect | Comparison | Estimate | 5%CI | 95%CI | p Value |
|---|---|---|---|---|---|
| Sex | Male vs. Female | 1.648 | −0.751 | 4.047 | 0.176 |
| Species | Light-vented Bulbul vs. Collared Finchbill | 0.807 | −1.632 | 3.246 | 0.512 |
| Social Context | Solitary vs. Group | 1.718 | −1.060 | 4.496 | 0.222 |
| Novel Food Type | a vs. b | −4.478 | −7.582 | −1.374 | 0.005 |
| a vs. c | −0.405 | −2.895 | 2.085 | 0.747 | |
| a vs. d | −1.008 | −3.508 | 1.491 | 0.425 | |
| a vs. e | −2.338 | −4.992 | 0.317 | 0.084 | |
| b vs. c | 4.076 | 0.890 | 7.263 | 0.013 | |
| b vs. d | 3.459 | 0.277 | 6.642 | 0.033 | |
| b vs. e | 1.892 | −1.407 | 5.191 | 0.257 | |
| c vs. d | −0.603 | −3.226 | 2.020 | 0.649 | |
| c vs. e | −1.933 | −4.660 | 0.795 | 0.162 | |
| d vs. e | −1.330 | −4.055 | 1.396 | 0.335 | |
| Sex × Social Context | Male: Solitary vs. Group | 2.049 | −1.576 | 5.673 | 0.264 |
| Female: Solitary vs. Group | 1.434 | −2.468 | 5.335 | 0.467 | |
| Social Context × Sex | Solitary: Male vs. Female | 1.980 | −2.884 | 6.843 | 0.421 |
| Group: Male vs. Female | 1.365 | −0.484 | 3.214 | 0.146 | |
| Species × Social Context | Light-vented Bulbul: Solitary vs. Group | −1.784 | −5.038 | 1.469 | 0.279 |
| Collared Finchbill: Solitary vs. Group | 4.938 | 0.561 | 9.316 | 0.028 | |
| Social Context × Species | Solitary: Light-vented Bulbul vs. Collared Finchbill | −2.928 | −7.844 | 1.989 | 0.240 |
| Group: Light-vented Bulbul vs. Collared Finchbill | 3.795 | 1.817 | 5.773 | <0.001 | |
| Random Effect | variance | ±SE | |||
| Individual Identity | □ | 0.325 | 0.050 | □ | □ |
| Fixed Effect | Comparison | Estimate | 5%CI | 95%CI | p Value |
|---|---|---|---|---|---|
| Sex | Male vs. Female | −33.644 | −165.862 | 98.573 | 0.614 |
| Species | Light-vented Bulbul vs. Collared Finchbill | 166.747 | 17.030 | 316.465 | 0.029 |
| Social Context | Solitary vs. Group | −86.333 | −213.983 | 41.318 | 0.182 |
| Novel Food Type | a vs. b | 142.194 | −131.106 | 415.494 | 0.304 |
| a vs. c | 291.266 | 45.710 | 536.823 | 0.013 | |
| a vs. d | 313.216 | 67.853 | 558.579 | 0.013 | |
| a vs. e | 140.774 | −130.333 | 411.882 | 0.305 | |
| b vs. c | 149.072 | −2.524 | 300.669 | 0.054 | |
| b vs. d | 171.022 | 23.214 | 318.831 | 0.024 | |
| b vs. e | −1.420 | −193.027 | 190.188 | 0.988 | |
| c vs. d | 0.547 | −50.153 | 94.053 | 0.547 | |
| c vs. e | −150.492 | −302.676 | 1.692 | 0.053 | |
| d vs. e | −172.442 | −322.695 | −22.188 | 0.026 | |
| Sex × Social Context | Male: Solitary vs. Group | 4.094 | −136.622 | 144.810 | 0.954 |
| Female: Solitary vs. Group | −203.603 | −434.561 | 27.354 | 0.083 | |
| Social Context × Sex | Solitary: Male vs. Female | 43.602 | −128.504 | 215.709 | 0.616 |
| Group: Male vs. Female | −164.095 | −355.588 | 27.398 | 0.092 | |
| Species × Social Context | Light-vented Bulbul: Solitary vs. Group | 24.470 | −266.387 | 315.326 | 0.868 |
| Collared Finchbill: Solitary vs. Group | −120.031 | −262.203 | 22.141 | 0.097 | |
| Social Context × Species | Solitary: Light-vented Bulbul vs. Collared Finchbill | 222.697 | −55.667 | 501.062 | 0.115 |
| Group: Light-vented Bulbul vs. Collared Finchbill | 78.197 | −74.240 | 230.635 | 0.311 | |
| Random Effect | variance | ±SE | |||
| Individual Identity | □ | 1.729 | 0.265 | □ | □ |
| Fixed Effect | Comparison | Estimate | 5%CI | 95%CI | p Value |
|---|---|---|---|---|---|
| Sex | Male vs. Female | −0.124 | −0.633 | 0.384 | 0.628 |
| Species | Light-vented Bulbul vs. Collared Finchbill | 0.077 | −0.578 | 0.424 | 0.760 |
| Social Context | Solitary vs. Group | 0.385 | −0.188 | 0.958 | 0.185 |
| Novel Food Type | a vs. b | −0.104 | −0.792 | 0.583 | 0.763 |
| a vs. c | 0.192 | −0.489 | 0.873 | 0.576 | |
| a vs. d | 0.660 | 0.075 | 1.225 | 0.027 | |
| a vs. e | 0.555 | −0.015 | 1.126 | 0.056 | |
| b vs. c | 0.297 | −0.379 | 0.973 | 0.385 | |
| b vs. d | 0.754 | 0.186 | 1.323 | 0.010 | |
| b vs. e | 0.660 | 0.095 | 1.224 | 0.022 | |
| c vs. d | 0.458 | −0.110 | 1.025 | 0.113 | |
| c vs. e | 0.363 | −0.197 | 0.923 | 0.201 | |
| d vs. e | −0.095 | −0.513 | 0.324 | 0.655 | |
| Sex × Social Context | Male: Solitary vs. Group | 0.465 | −0.189 | 1.119 | 0.161 |
| Female: Solitary vs. Group | 0.297 | −0.628 | 1.221 | 0.525 | |
| Social Context × Sex | Solitary: Male vs. Female | −0.028 | −1.063 | 1.007 | 0.957 |
| Group: Male vs. Female | −0.196 | −0.596 | 0.204 | 0.332 | |
| Species × Social Context | Light-vented Bulbul: Solitary vs. Group | 0.285 | −0.433 | 1.003 | 0.432 |
| Collared Finchbill: Solitary vs. Group | 0.490 | −0.375 | 1.356 | 0.263 | |
| Social Context × Species | Solitary: Light-vented Bulbul vs. Collared Finchbill | −0.193 | −1.191 | 0.805 | 0.702 |
| Group: Light-vented Bulbul vs. Collared Finchbill | 0.012 | −0.431 | 0.455 | 0.956 | |
| Random Effect | variance | ±SE | |||
| Individual Identity | □ | 0.358 | 0.055 | □ | □ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Wang, X.; Lu, J.; Li, L.; Li, D.; Xing, X.; Lei, F. Eating More and Fighting Less: Social Foraging Is a Potential Advantage for Successful Expansion of Bird Source Populations. Biology 2022, 11, 1496. https://doi.org/10.3390/biology11101496
Li X, Wang X, Lu J, Li L, Li D, Xing X, Lei F. Eating More and Fighting Less: Social Foraging Is a Potential Advantage for Successful Expansion of Bird Source Populations. Biology. 2022; 11(10):1496. https://doi.org/10.3390/biology11101496
Chicago/Turabian StyleLi, Xiang, Xiaochen Wang, Jinyao Lu, Li Li, Dongming Li, Xiaoying Xing, and Fumin Lei. 2022. "Eating More and Fighting Less: Social Foraging Is a Potential Advantage for Successful Expansion of Bird Source Populations" Biology 11, no. 10: 1496. https://doi.org/10.3390/biology11101496
APA StyleLi, X., Wang, X., Lu, J., Li, L., Li, D., Xing, X., & Lei, F. (2022). Eating More and Fighting Less: Social Foraging Is a Potential Advantage for Successful Expansion of Bird Source Populations. Biology, 11(10), 1496. https://doi.org/10.3390/biology11101496