



Plasma Amino Acids and Acylcarnitines Are Associated with the Female but Not Male Adolescent Swimmer’s Performance: An Integration between Mass Spectrometry and Complex Network Approaches

 , , , , and
, , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
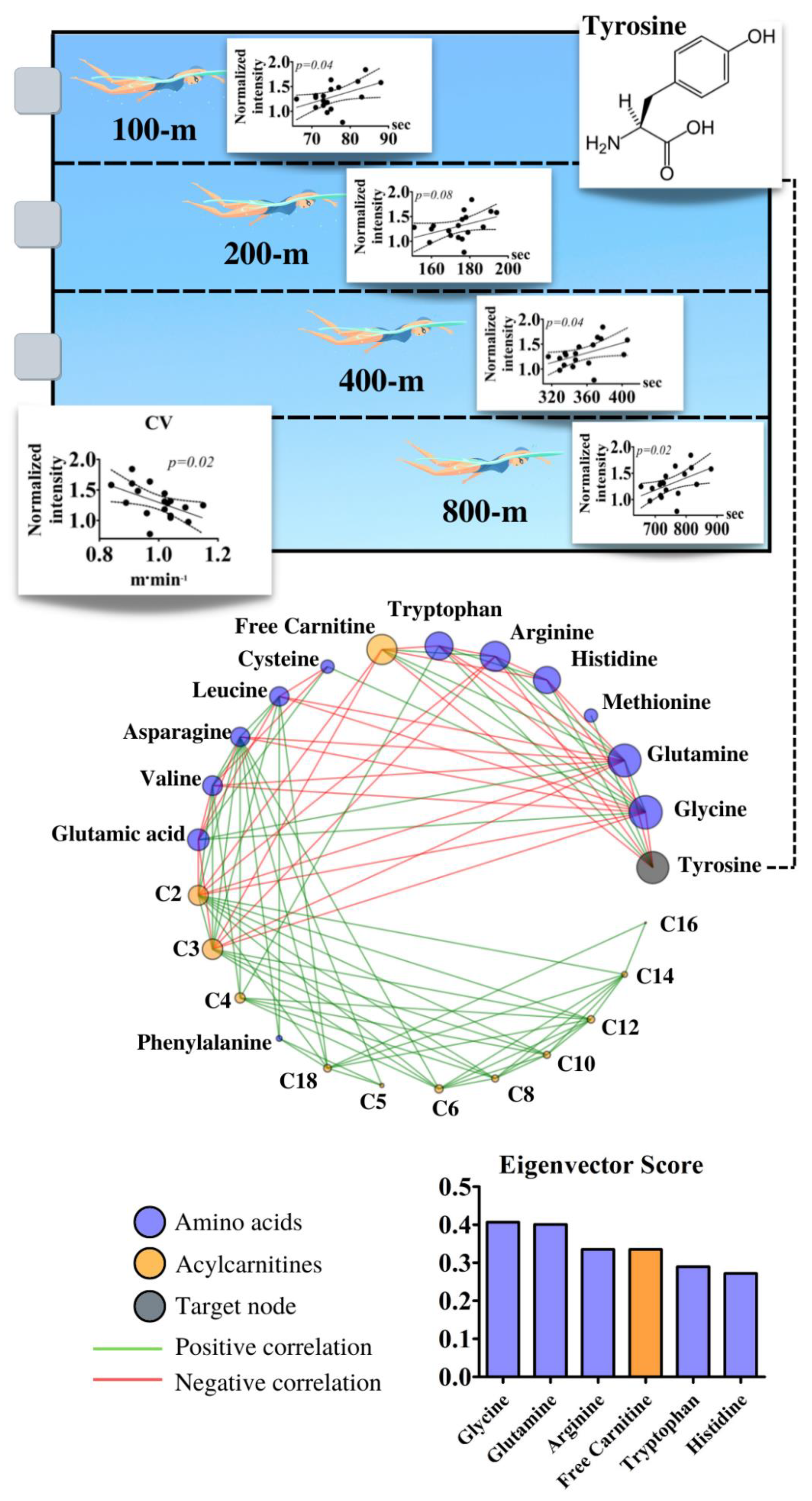{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
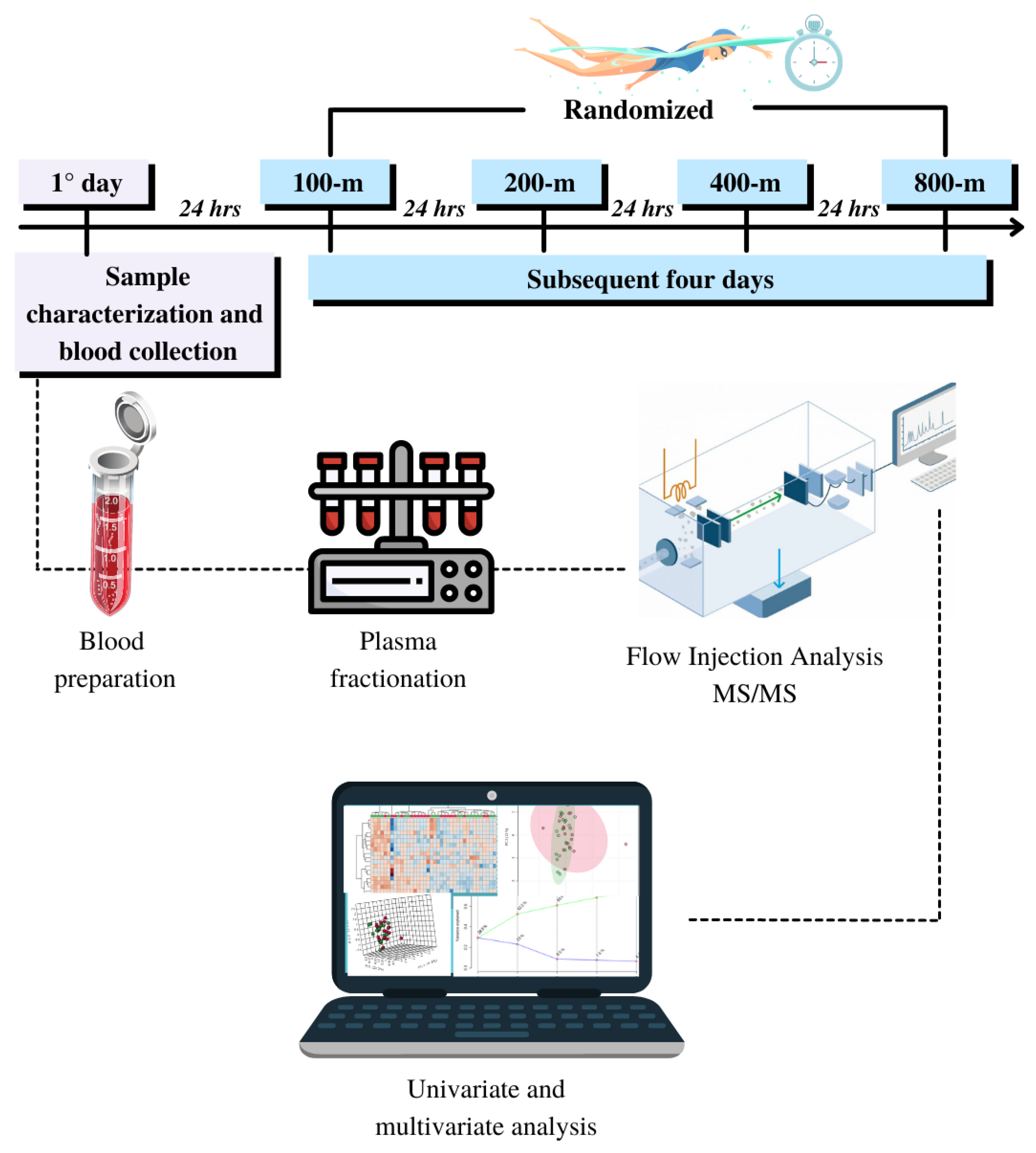
2.2. Experimental Design
2.3. Metabolomic Analysis
2.4. Tandem MS Analysis
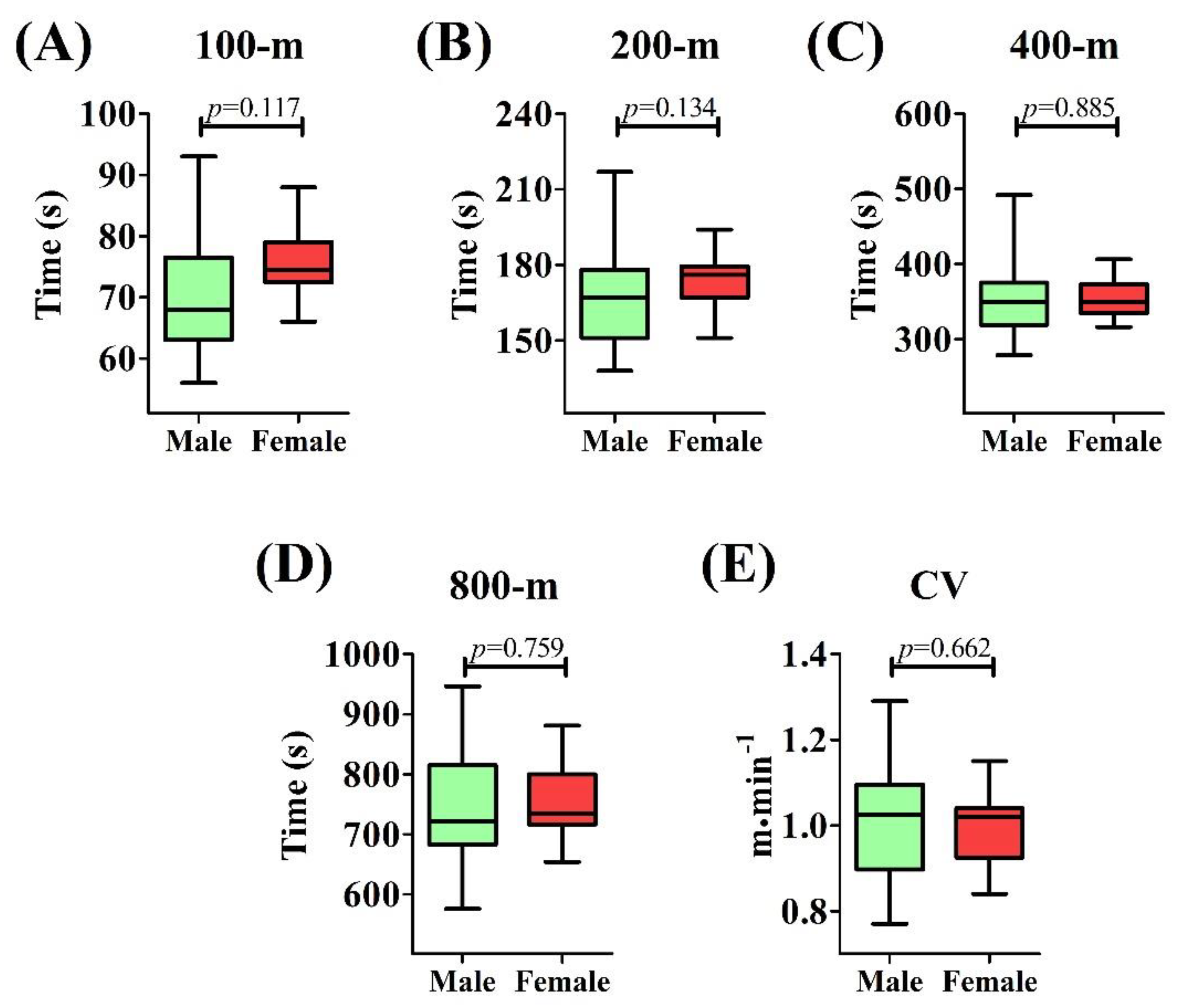
2.5. Performance Trials and Critical Velocity Assessment
2.6. Univariate and Multivariate Analysis
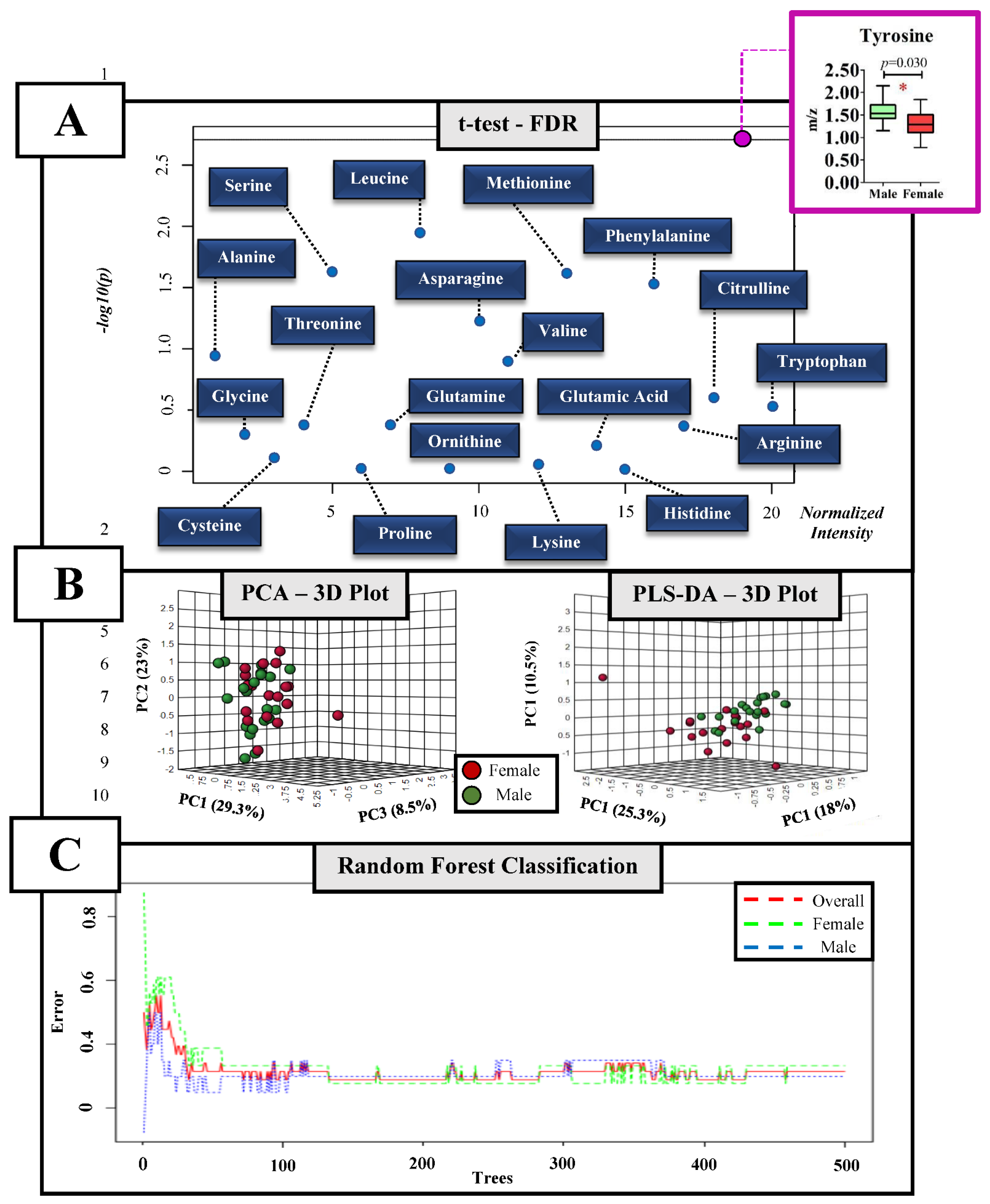
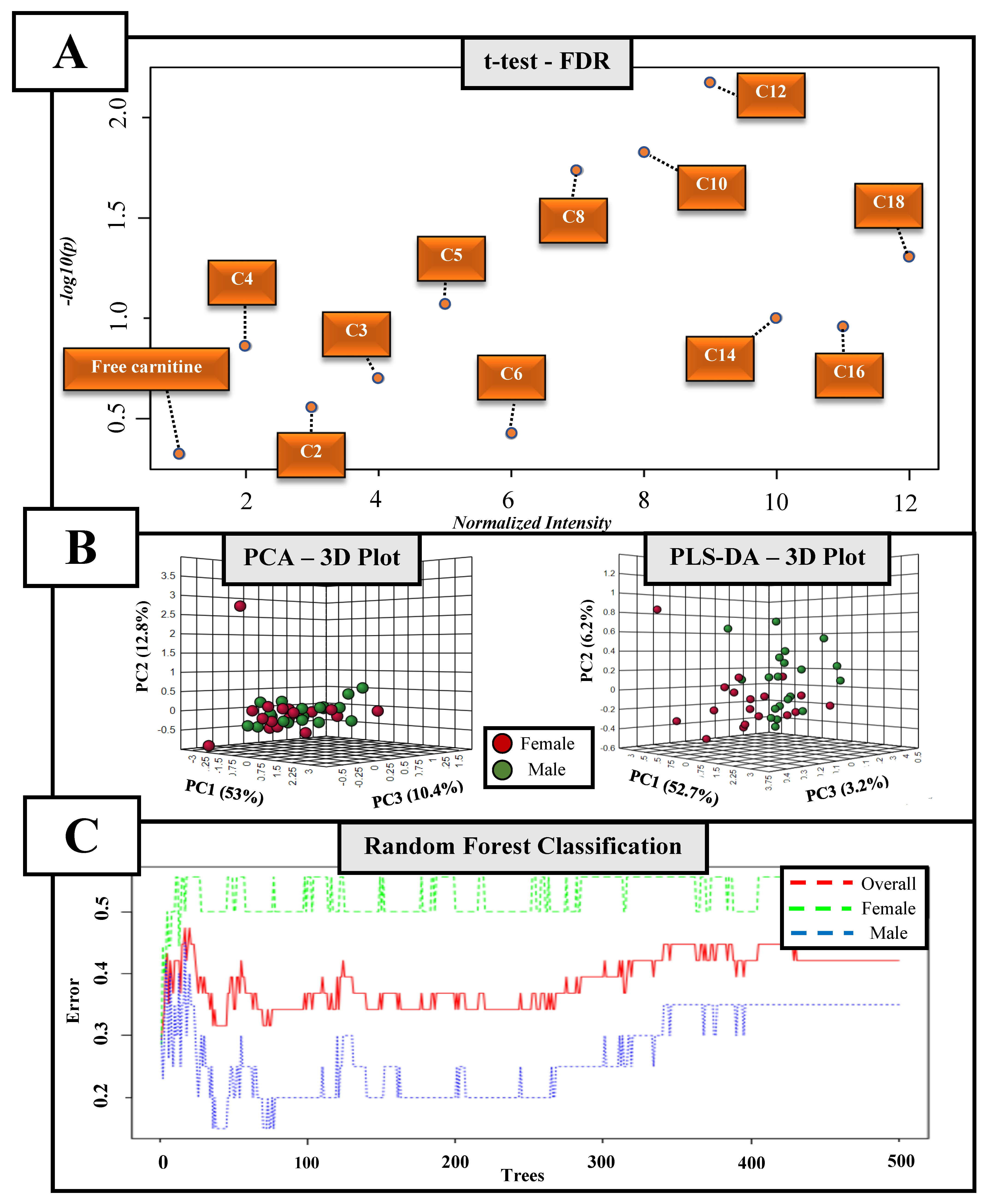
3. Results
4. Discussion
Limitations and Future Perspectives
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sawyer, S.M.; Azzopardi, P.S.; Wickremarathne, D.; Patton, G.C. The age of adolescence. Lancet Child Adolesc. Health 2018, 2, 223–228. [Google Scholar] [CrossRef]
- Christian, P.; Smith, E.R. Adolescent Undernutrition: Global Burden, Physiology, and Nutritional Risks. Ann. Nutr. Metab. 2018, 72, 316–328. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Nutrition in Adolescence: Issues and Challenges for the Health Sector: Issues in Adolescent Health and Development; WHO Press: Geneva, Switzerland, 2005.
- Lee, S.; Lim, H. Development of an Evidence-based Nutritional Intervention Protocol for Adolescent Athletes. J. Exerc. Nutr. Biochem. 2019, 23, 29–38. [Google Scholar] [CrossRef]
- Ferens, K.; Przeliorz-Pyszczek, A. Nutritional principles of adolescent swimmers. Cent. Eur. J. Sport Sci. Med. 2019, 25, 51–61. [Google Scholar] [CrossRef]
- Feijen, S.; Tate, A.; Kuppens, K.; Claes, A.; Struyf, F. Swim-Training Volume and Shoulder Pain Across the Life Span of the Competitive Swimmer: A Systematic Review. J. Athl. Train. 2020, 55, 32–41. [Google Scholar] [CrossRef] [Green Version]
- Shaw, G.; Boyd, K.T.; Burke, L.M.; Koivisto, A. Nutrition for swimming. Int. J. Sport Nutr. Exerc. Metab. 2014, 24, 360–372. [Google Scholar] [CrossRef] [Green Version]
- Shomaker, L.B.; Tanofsky-Kraff, M.; Savastano, D.M.; Kozlosky, M.; Columbo, K.M.; Wolkoff, L.E.; Zocca, J.M.; Brady, S.M.; Yanovski, S.Z.; Crocker, M.K.; et al. Puberty and observed energy intake: Boy, can they eat! Am. J. Clin. Nutr. 2010, 92, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, T.M.; Morais, J.E.; Marques, M.C.; Silva, A.J.; Marinho, D.A.; Kee, Y.H. Hydrodynamic profile of young swimmers: Changes over a competitive season. Scand J. Med. Sci. Sports 2015, 25, e184–e196. [Google Scholar] [CrossRef]
- Sammoud, S.; Nevill, A.M.; Negra, Y.; Bouguezzi, R.; Chaabene, H.; Hachana, Y. 100-m Breaststroke Swimming Performance in Youth Swimmers: The Predictive Value of Anthropometrics. Pediatr. Exerc. Sci. 2018, 30, 393–401. [Google Scholar] [CrossRef]
- Zacca, R.; Toubekis, A.; Freitas, L.; Silva, A.F.; Azevedo, R.; Vilas-Boas, J.P.; Pyne, D.B.; Castro, F.A.S.; Fernandes, R.J. Effects of detraining in age-group swimmers performance, energetics and kinematics. J. Sports Sci. 2019, 37, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Kraemer, M.B.; Garbuio, A.L.P.; Kaneko, L.O.; Gobatto, C.A.; Manchado-Gobatto, F.B.; Dos Reis, I.G.M.; Messias, L.H.D. Associations among sleep, hematologic profile, and aerobic and anerobic capacity of young swimmers: A complex network approach. Front. Physiol. 2022, 13, 948422. [Google Scholar] [CrossRef]
- Tipton, K.D.; Ferrando, A.A.; Williams, B.D.; Wolfe, R.R. Muscle protein metabolism in female swimmers after a combination of resistance and endurance exercise. J. Appl. Physiol. 1996, 81, 2034–2038. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Kato, H.; Suzuki, H.; Mizugaki, A.; Ezaki, T.; Ogita, F. Within-Day Amino Acid Intakes and Nitrogen Balance in Male Collegiate Swimmers during the General Preparation Phase. Nutrients 2018, 10, 1809. [Google Scholar] [CrossRef] [Green Version]
- Krause, S.; Langrock, M.; Weiss, M. Influence of seasonal variations in training loads on selected amino acids and parameters of the psychoimmunological network in a swimming team. Int. J. Sports Med. 2002, 23, 380–387. [Google Scholar] [CrossRef]
- Hsueh, C.F.; Wu, H.J.; Tsai, T.S.; Wu, C.L.; Chang, C.K. The Effect of Branched-Chain Amino Acids, Citrulline, and Arginine on High-Intensity Interval Performance in Young Swimmers. Nutrients 2018, 10, 1979. [Google Scholar] [CrossRef] [Green Version]
- Gnoni, A.; Longo, S.; Gnoni, G.V.; Giudetti, A.M. Carnitine in Human Muscle Bioenergetics: Can Carnitine Supplementation Improve Physical Exercise? Molecules 2020, 25, 182. [Google Scholar] [CrossRef] [Green Version]
- Kreider, R.B.; Miriel, V.; Bertun, E. Amino acid supplementation and exercise performance. Analysis of the proposed ergogenic value. Sports Med. 1993, 16, 190–209. [Google Scholar] [CrossRef]
- Flanagan, J.L.; Simmons, P.A.; Vehige, J.; Willcox, M.D.; Garrett, Q. Role of carnitine in disease. Nutr. Metab. 2010, 7, 30. [Google Scholar] [CrossRef] [Green Version]
- Brass, E.P. Supplemental carnitine and exercise. Am. J. Clin. Nutr. 2000, 72, 618S–623S. [Google Scholar] [CrossRef] [Green Version]
- Blomstrand, E. Amino acids and central fatigue. Amino Acids 2001, 20, 25–34. [Google Scholar] [CrossRef]
- Lemon, P.W.; Proctor, D.N. Protein intake and athletic performance. Sports Med. 1991, 12, 313–325. [Google Scholar] [CrossRef] [PubMed]
- Pasiakos, S.M.; Lieberman, H.R.; McLellan, T.M. Effects of protein supplements on muscle damage, soreness and recovery of muscle function and physical performance: A systematic review. Sports Med. 2014, 44, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Williams, M. Dietary supplements and sports performance: Amino acids. J. Int. Soc. Sports Nutr. 2005, 2, 63–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G. Dietary protein intake and human health. Food Funct. 2016, 7, 1251–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nunes, E.A.; Colenso-Semple, L.; McKellar, S.R.; Yau, T.; Ali, M.U.; Fitzpatrick-Lewis, D.; Sherifali, D.; Gaudichon, C.; Tome, D.; Atherton, P.J.; et al. Systematic review and meta-analysis of protein intake to support muscle mass and function in healthy adults. J. Cachexia Sarcopenia Muscle 2022, 13, 795–810. [Google Scholar] [CrossRef]
- Bassini, A.; Cameron, L.C. Sportomics: Building a new concept in metabolic studies and exercise science. Biochem. Biophys. Res. Commun. 2014, 445, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Bongiovanni, T.; Pintus, R.; Dessi, A.; Noto, A.; Sardo, S.; Finco, G.; Corsello, G.; Fanos, V. Sportomics: Metabolomics applied to sports. The new revolution? Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 11011–11019. [Google Scholar] [CrossRef]
- Bragazzi, N.L.; Khoramipour, K.; Chaouachi, A.; Chamari, K. Toward Sportomics: Shifting From Sport Genomics to Sport Postgenomics and Metabolomics Specialties. Promises, Challenges, and Future Perspectives. Int. J. Sports Physiol. Perform. 2020, 15, 1201–1202. [Google Scholar] [CrossRef]
- Sellami, M.; Elrayess, M.A.; Puce, L.; Bragazzi, N.L. Molecular Big Data in Sports Sciences: State-of-Art and Future Prospects of OMICS-Based Sports Sciences. Front. Mol. Biosci. 2021, 8, 815410. [Google Scholar] [CrossRef] [PubMed]
- Kuehnbaum, N.L.; Gillen, J.B.; Kormendi, A.; Lam, K.P.; DiBattista, A.; Gibala, M.J.; Britz-McKibbin, P. Multiplexed separations for biomarker discovery in metabolomics: Elucidating adaptive responses to exercise training. Electrophoresis 2015, 36, 2226–2236. [Google Scholar] [CrossRef]
- Sakaguchi, C.A.; Nieman, D.C.; Signini, E.F.; Abreu, R.M.; Catai, A.M. Metabolomics-Based Studies Assessing Exercise-Induced Alterations of the Human Metabolome: A Systematic Review. Metabolites 2019, 9, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schranner, D.; Kastenmuller, G.; Schonfelder, M.; Romisch-Margl, W.; Wackerhage, H. Metabolite Concentration Changes in Humans After a Bout of Exercise: A Systematic Review of Exercise Metabolomics Studies. Sports Med. Open. 2020, 6, 11. [Google Scholar] [CrossRef] [Green Version]
- Goncalves, L.C.O.; Magalhaes-Neto, A.M.; Bassini, A.; Prado, E.S.; Muniz-Santos, R.; Verli, M.V.A.; Jurisica, L.; Lopes, J.S.S.; Jurisica, I.; Andrade, C.M.B.; et al. Sportomics suggests that albuminuria is a sensitive biomarker of hydration in cross combat. Sci. Rep. 2022, 12, 8150. [Google Scholar] [CrossRef]
- Pintus, R.; Bongiovanni, T.; Corbu, S.; Francavilla, V.C.; Dess, I.A.; Noto, A.; Corsello, G.; Finco, G.; Fanos, V.; Cesare Marincola, F. Sportomics in professional soccer players: Metabolomics results during preseason. J. Sports Med. Phys. Fitness 2021, 61, 324–330. [Google Scholar] [CrossRef]
- Coelho, W.S.; Viveiros de Castro, L.; Deane, E.; Magno-Franca, A.; Bassini, A.; Cameron, L.C. Investigating the Cellular and Metabolic Responses of World-Class Canoeists Training: A Sportomics Approach. Nutrients 2016, 8, 719. [Google Scholar] [CrossRef] [PubMed]
- Berthoin, S.; Baquet, G.; Dupont, G.; Blondel, N.; Mucci, P. Critical velocity and anaerobic distance capacity in prepubertal children. Can. J. Appl. Physiol. 2003, 28, 561–575. [Google Scholar] [CrossRef] [PubMed]
- Neiva, H.P.; Fernandes, R.J.; Vilas-Boas, J.P. Anaerobic critical velocity in four swimming techniques. Int. J. Sports Med. 2011, 32, 195–198. [Google Scholar] [CrossRef]
- Toubekis, A.G.; Tokmakidis, S.P. Metabolic responses at various intensities relative to critical swimming velocity. J. Strength Cond. Res. 2013, 27, 1731–1741. [Google Scholar] [CrossRef] [PubMed]
- Toubekis, A.G.; Tsami, A.P.; Tokmakidis, S.P. Critical velocity and lactate threshold in young swimmers. Int. J. Sports Med. 2006, 27, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Sarafian, M.H.; Gaudin, M.; Lewis, M.R.; Martin, F.P.; Holmes, E.; Nicholson, J.K.; Dumas, M.E. Objective set of criteria for optimization of sample preparation procedures for ultra-high throughput untargeted blood plasma lipid profiling by ultra performance liquid chromatography-mass spectrometry. Anal. Chem. 2014, 86, 5766–5774. [Google Scholar] [CrossRef]
- Cicalini, I.; Valentinuzzi, S.; Pieragostino, D.; Consalvo, A.; Zucchelli, M.; Donzelli, S.; Ambrogi, D.; Brown, H.A.; Calton, L.J.; Stuppia, L.; et al. Analytical Evaluation of the Ideal Strategy for High-Throughput Flow Injection Analysis by Tandem Mass Spectrometry in Routine Newborn Screening. Metabolites 2021, 11, 473. [Google Scholar] [CrossRef]
- Wakayoshi, K.; Ikuta, K.; Yoshida, T.; Udo, M.; Moritani, T.; Mutoh, Y.; Miyashita, M. Determination and validity of critical velocity as an index of swimming performance in the competitive swimmer. Eur. J. Appl. Physiol. Occup. Physiol. 1992, 64, 153–157. [Google Scholar] [CrossRef] [PubMed]
- Wakayoshi, K.; Yoshida, T.; Kasai, T.; Moritani, T.; Mutoh, Y.; Miyashita, M. Validity of critical velocity as swimming fatigue threshold in the competitive swimmer. Ann. Physiol. Anthropol. 1992, 11, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Wakayoshi, K.; Yoshida, T.; Udo, M.; Kasai, T.; Moritani, T.; Mutoh, Y.; Miyashita, M. A simple method for determining critical speed as swimming fatigue threshold in competitive swimming. Int. J. Sports Med. 1992, 13, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Vanhatalo, A. The ‘Critical Power’ Concept: Applications to Sports Performance with a Focus on Intermittent High-Intensity Exercise. Sports Med. 2017, 47, 65–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.M.; Vanhatalo, A.; Burnley, M.; Morton, R.H.; Poole, D.C. Critical power: Implications for determination of V O2max and exercise tolerance. Med. Sci. Sports Exerc. 2010, 42, 1876–1890. [Google Scholar] [CrossRef] [PubMed]
- Poole, D.C.; Burnley, M.; Vanhatalo, A.; Rossiter, H.B.; Jones, A.M. Critical Power: An Important Fatigue Threshold in Exercise Physiology. Med. Sci. Sports Exerc. 2016, 48, 2320–2334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanhatalo, A.; Jones, A.M.; Burnley, M. Application of critical power in sport. Int. J. Sports Physiol. Perform. 2011, 6, 128–136. [Google Scholar] [CrossRef] [Green Version]
- MetaboAnalyst. Available online: https://www.metaboanalyst.ca/ (accessed on 20 June 2022).
- Gobatto, C.A.; Torres, R.S.; Moura, F.A.; Cunha, S.A.; Giometti, C.B.; Araujo, G.G.; Sousa, F.A.B.; Manchado-Gobatto, F.B. Corresponding Assessment Scenarios in Laboratory and on-Court Tests: Centrality Measurements by Complex Networks Analysis in Young Basketball Players. Sci. Rep. 2020, 10, 8620. [Google Scholar] [CrossRef]
- Hagberg, A.A.; Schult, D.A.; Swart, P.J. Exploring network structure, dynamics, and function using NetworkX. In Proceedings of the 7th Python in Science Conference, Gäel Varoquaux. Pasadena, CA, USA, 19–24 August 2008; Vaught, T., Millman, J., Eds.; 2008; pp. 11–15. [Google Scholar]
- Newman, M.E.J. Networks: An Introduction; Oxford University Press: Oxford, UK, 2010. [Google Scholar]
- Meeusen, R.; Decroix, L. Nutritional Supplements and the Brain. Int. J. Sport Nutr. Exerc. Metab. 2018, 28, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Jongkees, B.J.; Hommel, B.; Kuhn, S.; Colzato, L.S. Effect of tyrosine supplementation on clinical and healthy populations under stress or cognitive demands—A review. J. Psychiatr. Res. 2015, 70, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Espana, R.A.; Schmeichel, B.E.; Berridge, C.W. Norepinephrine at the nexus of arousal, motivation and relapse. Brain Res. 2016, 1641, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harley, C.W. Norepinephrine and dopamine as learning signals. Neural Plast. 2004, 11, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, T.F.; Mora, F.; Myers, R.D. Dopamine and thermoregulation: An evaluation with special reference to dopaminergic pathways. Neurosci. Biobehav. Rev. 1985, 9, 589–598. [Google Scholar] [CrossRef] [PubMed]
- Maki-Marttunen, V.; Andreassen, O.A.; Espeseth, T. The role of norepinephrine in the pathophysiology of schizophrenia. Neurosci. Biobehav. Rev 2020, 118, 298–314. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.D. Thermoregulation and norepinephrine. Science 1969, 165, 1030–1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, X.; Hasegawa, H. Central dopaminergic neurotransmission plays an important role in thermoregulation and performance during endurance exercise. Eur. J. Sport Sci. 2016, 16, 818–828. [Google Scholar] [CrossRef]
- Coull, N.A.; Watkins, S.L.; Aldous, J.W.; Warren, L.K.; Chrismas, B.C.; Dascombe, B.; Mauger, A.R.; Abt, G.; Taylor, L. Effect of tyrosine ingestion on cognitive and physical performance utilising an intermittent soccer performance test (iSPT) in a warm environment. Eur. J. Appl. Physiol. 2015, 115, 373–386. [Google Scholar] [CrossRef] [PubMed]
- Meeusen, R.; Watson, P. Amino acids and the brain: Do they play a role in “central fatigue”? Int. J. Sport Nutr. Exerc. Metab. 2007, 17, S37–S46. [Google Scholar] [CrossRef]
- Meeusen, R.; Watson, P.; Dvorak, J. The brain and fatigue: New opportunities for nutritional interventions? J. Sports Sci. 2006, 24, 773–782. [Google Scholar] [CrossRef]
- Struder, H.K.; Hollmann, W.; Platen, P.; Donike, M.; Gotzmann, A.; Weber, K. Influence of paroxetine, branched-chain amino acids and tyrosine on neuroendocrine system responses and fatigue in humans. Horm. Metab. Res. 1998, 30, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Tumilty, L.; Davison, G.; Beckmann, M.; Thatcher, R. Oral tyrosine supplementation improves exercise capacity in the heat. Eur J. Appl. Physiol. 2011, 111, 2941–2950. [Google Scholar] [CrossRef] [PubMed]
- Tumilty, L.; Davison, G.; Beckmann, M.; Thatcher, R. Failure of oral tyrosine supplementation to improve exercise performance in the heat. Med. Sci. Sports Exerc. 2014, 46, 1417–1425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watson, P.; Enever, S.; Page, A.; Stockwell, J.; Maughan, R.J. Tyrosine supplementation does not influence the capacity to perform prolonged exercise in a warm environment. Int. J. Sport Nutr. Exerc. Metab. 2012, 22, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Sirakov, M.; Plateroti, M. Thyroid Hormone Signaling and Function: News from Classical and Emerging Models. Cells 2022, 11, 453. [Google Scholar] [CrossRef]
- Citterio, C.E.; Targovnik, H.M.; Arvan, P. The role of thyroglobulin in thyroid hormonogenesis. Nat. Rev. Endocrinol. 2019, 15, 323–338. [Google Scholar] [CrossRef] [PubMed]
- Sutton, E.E.; Coill, M.R.; Deuster, P.A. Ingestion of tyrosine: Effects on endurance, muscle strength, and anaerobic performance. Int. J. Sport Nutr. Exerc. Metab. 2005, 15, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Couto, M.; Barbosa, C.; Silva, D.; Rudnitskaya, A.; Delgado, L.; Moreira, A.; Rocha, S.M. Oxidative stress in asthmatic and non-asthmatic adolescent swimmers-A breathomics approach. Pediatr. Allergy Immunol. 2017, 28, 452–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, M.; Wu, C.; Jing, C.; Shen, X.; He, M.; Wang, L.; Guo, Q.; Yan, Y.; Yan, X.; Yang, R. Blood Metabolomics Analysis Identifies Differential Serum Metabolites in Elite and Sub-elite Swimmers. Front. Physiol. 2022, 13, 858869. [Google Scholar] [CrossRef] [PubMed]
- Coqueiro, A.Y.; Rogero, M.M.; Tirapegui, J. Glutamine as an Anti-Fatigue Amino Acid in Sports Nutrition. Nutrients 2019, 11, 863. [Google Scholar] [CrossRef] [PubMed]
- d’Unienville, N.M.A.; Blake, H.T.; Coates, A.M.; Hill, A.M.; Nelson, M.J.; Buckley, J.D. Effect of food sources of nitrate, polyphenols, L-arginine and L-citrulline on endurance exercise performance: A systematic review and meta-analysis of randomised controlled trials. J. Int. Soc. Sports Nutr. 2021, 18, 76. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M. Dosing and efficacy of glutamine supplementation in human exercise and sport training. J. Nutr. 2008, 138, 2045S–2049S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlic, H.; Lohninger, A. Supplementation of L-carnitine in athletes: Does it make sense? Nutrition 2004, 20, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Walsh, N.P.; Blannin, A.K.; Robson, P.J.; Gleeson, M. Glutamine, exercise and immune function. Links and possible mechanisms. Sports Med. 1998, 26, 177–191. [Google Scholar] [CrossRef] [PubMed]
- Esen, O.; Eser, M.C.; Abdioglu, M.; Benesova, D.; Gabrys, T.; Karayigit, R. Eight Days of L-Citrulline or L-Arginine Supplementation Did Not Improve 200-m and 100-m Swimming Time Trials. Int. J. Environ. Res. Public Health 2022, 19, 4462. [Google Scholar] [CrossRef] [PubMed]
- Kargotich, S.; Rowbottom, D.G.; Keast, D.; Goodman, C.; Dawson, B.; Morton, A.R. Plasma glutamine changes after high-intensity exercise in elite male swimmers. Res. Sports Med. 2005, 13, 7–21. [Google Scholar] [CrossRef]
- Trappe, S.W.; Costill, D.L.; Goodpaster, B.; Vukovich, M.D.; Fink, W.J. The effects of L-carnitine supplementation on performance during interval swimming. Int. J. Sports Med. 1994, 15, 181–185. [Google Scholar] [CrossRef]
- Hirabara, S.M.; Gorjao, R.; Levada-Pires, A.C.; Masi, L.N.; Hatanaka, E.; Cury-Boaventura, M.F.; da Silva, E.B.; Santos-Oliveira, L.C.D.; Sousa Diniz, V.L.; Serdan, T.A.D.; et al. Host cell glutamine metabolism as a potential antiviral target. Clin. Sci. 2021, 135, 305–325. [Google Scholar] [CrossRef] [PubMed]
- Cruzat, V.; Macedo Rogero, M.; Noel Keane, K.; Curi, R.; Newsholme, P. Glutamine: Metabolism and Immune Function, Supplementation and Clinical Translation. Nutrients 2018, 10, 1564. [Google Scholar] [CrossRef] [Green Version]
- Adeva-Andany, M.M.; Carneiro-Freire, N.; Seco-Filgueira, M.; Fernandez-Fernandez, C.; Mourino-Bayolo, D. Mitochondrial beta-oxidation of saturated fatty acids in humans. Mitochondrion 2019, 46, 73–90. [Google Scholar] [CrossRef]
- Bremer, J. Carnitine--metabolism and functions. Physiol. Rev. 1983, 63, 1420–1480. [Google Scholar] [CrossRef] [PubMed]
- Morris, S.M., Jr. Arginine: Beyond protein. Am. J. Clin. Nutr. 2006, 83, 508S–512S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stechmiller, J.K.; Childress, B.; Cowan, L. Arginine supplementation and wound healing. Nutr. Clin. Pract. 2005, 20, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Holecek, M. Histidine in Health and Disease: Metabolism, Physiological Importance, and Use as a Supplement. Nutrients 2020, 12, 848. [Google Scholar] [CrossRef] [Green Version]
- Segura, R.; Ventura, J.L. Effect of L-tryptophan supplementation on exercise performance. Int. J. Sports Med. 1988, 9, 301–305. [Google Scholar] [CrossRef]
- Stensrud, T.; Ingjer, F.; Holm, H.; Stromme, S.B. L-tryptophan supplementation does not improve running performance. Int. J. Sports Med. 1992, 13, 481–485. [Google Scholar] [CrossRef]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef]
- Brosnan, M.E.; Brosnan, J.T. Histidine Metabolism and Function. J. Nutr. 2020, 150, 2570S–2575S. [Google Scholar] [CrossRef]
- Stankovic, I.M.; Niu, S.; Hall, M.B.; Zaric, S.D. Role of aromatic amino acids in amyloid self-assembly. Int. J. Biol. Macromol. 2020, 156, 949–959. [Google Scholar] [CrossRef]
- Moller, S.E.; Moller, B.M.; Olesen, M.; Fjalland, B. Effects of oral contraceptives on plasma neutral amino acids and cholesterol during a menstrual cycle. Eur. J. Clin. Pharmacol. 1996, 50, 179–184. [Google Scholar] [CrossRef]
- He, W.; Dam, T.V.; Thogersen, R.; Hansen, M.; Bertram, H.C. Fluctuations in Metabolites and Bone Markers Across the Menstrual Cycle in Eumenorrheic Women and Oral Contraceptive Users. J. Clin. Endocrinol. Metab. 2022, 107, 1577–1588. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, F.M.M.; Sanches, P.H.G.; Silva, Á.A.R.; Reis, I.G.M.d.; Carvalho, P.d.O.; Porcari, A.M.; Messias, L.H.D. Plasma Amino Acids and Acylcarnitines Are Associated with the Female but Not Male Adolescent Swimmer’s Performance: An Integration between Mass Spectrometry and Complex Network Approaches. Biology 2022, 11, 1734. https://doi.org/10.3390/biology11121734
Mendes FMM, Sanches PHG, Silva ÁAR, Reis IGMd, Carvalho PdO, Porcari AM, Messias LHD. Plasma Amino Acids and Acylcarnitines Are Associated with the Female but Not Male Adolescent Swimmer’s Performance: An Integration between Mass Spectrometry and Complex Network Approaches. Biology. 2022; 11(12):1734. https://doi.org/10.3390/biology11121734
Chicago/Turabian StyleMendes, Flávio Marcio Macedo, Pedro Henrique Godoy Sanches, Álex Ap. Rosini Silva, Ivan Gustavo Masselli dos Reis, Patrícia de Oliveira Carvalho, Andréia M. Porcari, and Leonardo Henrique Dalcheco Messias. 2022. "Plasma Amino Acids and Acylcarnitines Are Associated with the Female but Not Male Adolescent Swimmer’s Performance: An Integration between Mass Spectrometry and Complex Network Approaches" Biology 11, no. 12: 1734. https://doi.org/10.3390/biology11121734
APA StyleMendes, F. M. M., Sanches, P. H. G., Silva, Á. A. R., Reis, I. G. M. d., Carvalho, P. d. O., Porcari, A. M., & Messias, L. H. D. (2022). Plasma Amino Acids and Acylcarnitines Are Associated with the Female but Not Male Adolescent Swimmer’s Performance: An Integration between Mass Spectrometry and Complex Network Approaches. Biology, 11(12), 1734. https://doi.org/10.3390/biology11121734