
First Report of Epimeletic and Acoustic Behavior in Mediterranean Common Bottlenose Dolphins (Tursiops truncatus) Carrying Dead Calves



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
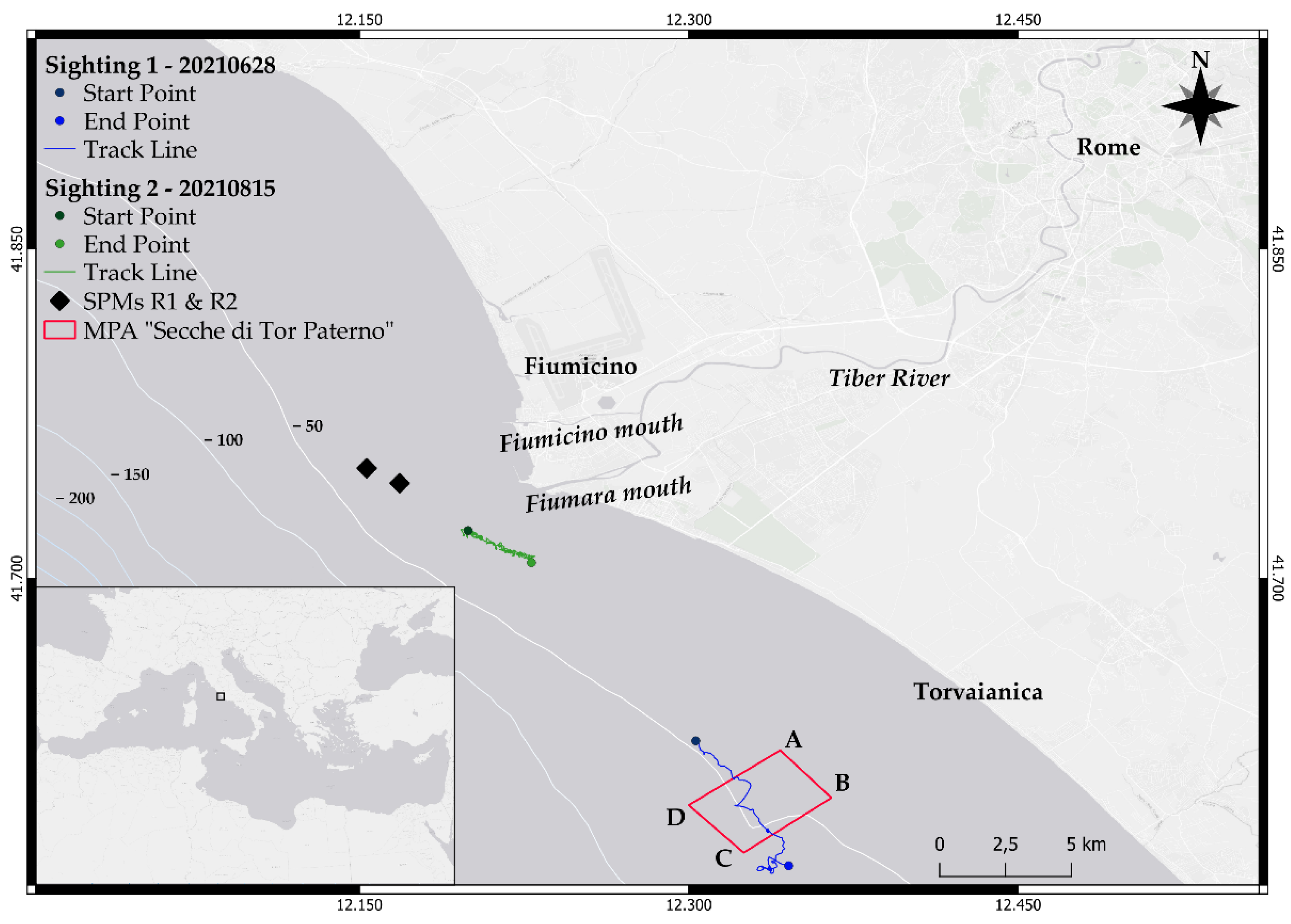
2.1. Study Area
2.2. Data Collection
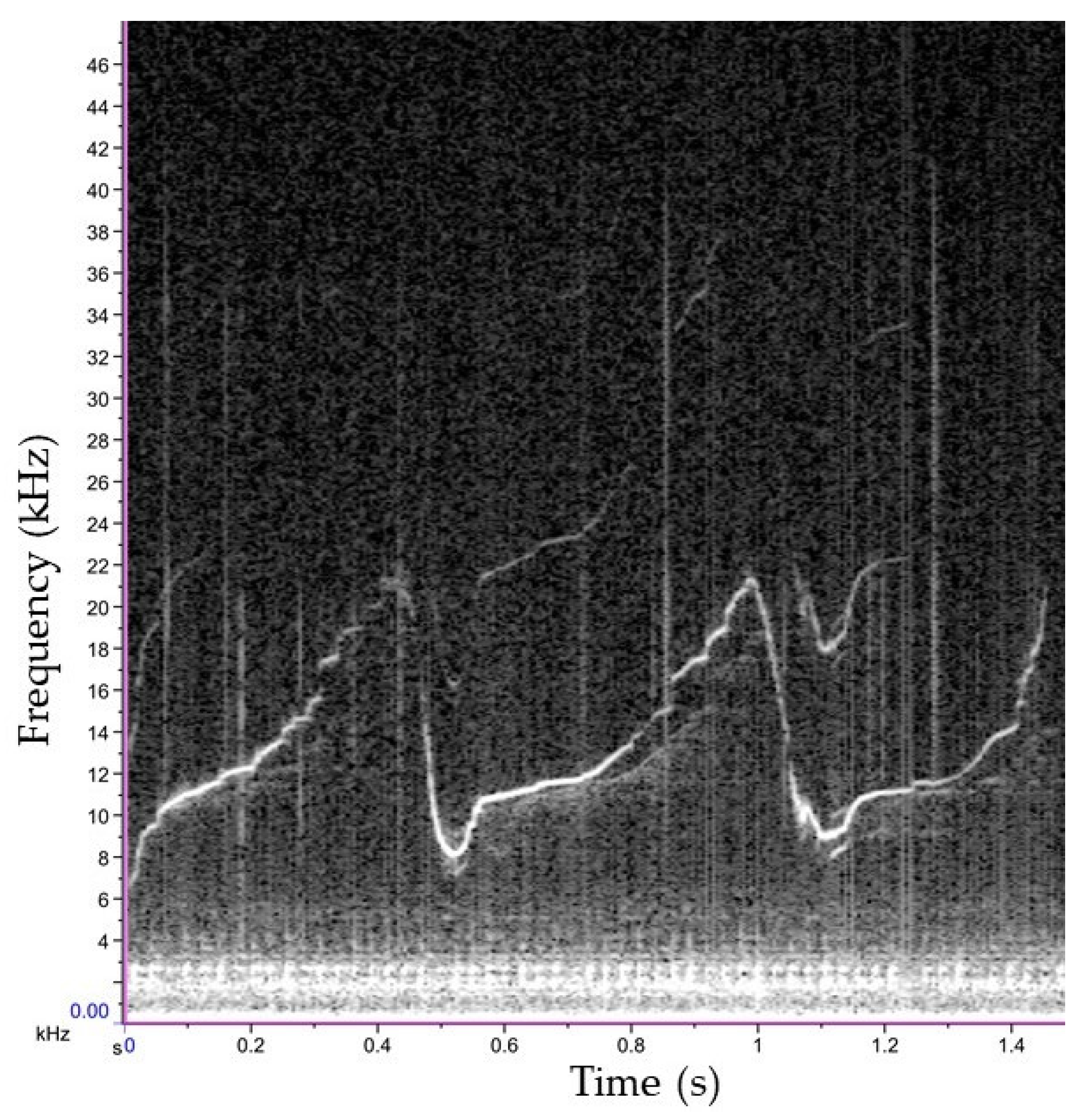
2.3. Data Analysis
3. Results
3.1. Sighting 1
3.2. Sighting 2
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Biro, D.; Humle, T.; Koops, K.; Sousa, C.; Hayashi, M.; Matsuzawa, T. Chimpanzee mothers at Bossou, Guinea carry the mummified remains of their dead infants. Curr. Biol. 2010, 20, R351–R352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bercovitch, F.B. Giraffe cow reaction to the death of her newborn calf. Afr. J. Ecol. 2013, 51, 376–379. [Google Scholar] [CrossRef]
- Douglas-Hamilton, I.; Bhalla, S.; Wittemyer, G.; Vollrath, F. Behavioural reactions of elephants towards a dying and deceased matriarch. Appl. Anim. Behav. Sci. 2006, 100, 87–102. [Google Scholar] [CrossRef]
- Reggente, M.A.L.V.; Papale, E.; McGinty, N.; Eddy, L.; de Lucia, G.A.; Bertulli, C.G. Social relationships and death-related behaviour in aquatic mammals: A systematic review. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20170260. [Google Scholar] [CrossRef] [Green Version]
- Bearzi, G.; Eddy, L.; Piwetz, S.; Reggente, M.A.L.; Cozzi, B. Cetacean behavior toward the dead and dying. In Encyclopedia of Animal Cognition and Behavior; Vonk, J., Shackelford, T., Eds.; Springer: Cham, Switzerland, 2017; pp. 1–8. ISBN 978-3-319-47829-6. [Google Scholar]
- Caldwell, M.C.; Caldwell, D.K. Epimeletic (care-giving) behavior in cetacea. In Whales, Porpoises and Dolphins; Norris, K.S., Ed.; University of California Press: Berkley, CA, USA, 1966; pp. 755–790. ISBN 978-0-520-32137-3. [Google Scholar]
- Reggente, M.A.L.; Alves, F.; Nicolau, C.; Freitas, L.; Cagnazzi, D.; Baird, R.W.; Galli, P. Nurturant behavior toward dead conspecifics in free-ranging mammals: New records for odontocetes and a general review. J. Mammal. 2016, 97, 1428–1434. [Google Scholar] [CrossRef] [Green Version]
- Ritter, F. Behavioral responses of rough-toothed dolphins to a dead newborn calf. Mar. Mammal Sci. 2007, 23, 429–433. [Google Scholar] [CrossRef]
- Cheng, Z.; Pine, M.K.; Huang, S.-L.; Wang, D.; Wu, H.; Wang, K. A case of epimeletic behavior and associated acoustic records of indo-pacific humpback dolphins (Sousa chinensis). J. Mammal. 2018, 99, 1112–1119. [Google Scholar] [CrossRef]
- Fertl, D.; Schiro, A. Carrying of dead calves by free-ranging texas bottlenose dolphins (Tursiops truncatus). Aquat. Mamm. 1994, 20, 53–56. [Google Scholar]
- Bearzi, G.; Kerem, D.; Furey, N.; Pitman, R.; Rendell, L.; Reeves, R. Whale and dolphin behavioural responses to dead conspecifics. Zoology 2018, 128, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Gridley, T.; Nastasi, A.; Kriesell, H.; Elwen, S. The acoustic repertoire of wild common bottlenose dolphins (Tursiops truncatus) in Walvis Bay, Namibia. Bioacoustics 2015, 24, 153–174. [Google Scholar] [CrossRef] [Green Version]
- Perez-Ortega, B.; Daw, R.; Paradee, B.; Gimbrere, E.; May-Collado, L.J. Dolphin-watching boats affect whistle frequency modulation in bottlenose dolphins. Front. Mar. Sci. 2021, 8, 102. [Google Scholar] [CrossRef]
- La Manna, G.; Manghi, M.; Pavan, G.; Lo Mascolo, F.; Sarà, G. Behavioural strategy of common bottlenose dolphins (Tursiops truncatus) in response to different kinds of boats in the waters of Lampedusa Island (Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 2013, 23, 745–757. [Google Scholar] [CrossRef]
- May-Collado, L.J.; Wartzok, D. A comparison of bottlenose dolphin whistles in the Atlantic Ocean: Factors promoting whistle variation. J. Mammal. 2008, 89, 1229–1240. [Google Scholar] [CrossRef] [Green Version]
- Esch, H.C.; Sayigh, L.S.; Blum, J.E.; Wells, R.S. Whistles as potential indicators of stress in bottlenose dolphins (Tursiops truncatus). J. Mammal. 2009, 90, 638–650. [Google Scholar] [CrossRef] [Green Version]
- Kuczaj, S.A.; Frick, E.E.; Jones, B.L.; Lea, J.S.E.; Beecham, D.; Schnöller, F. Underwater observations of dolphin reactions to a distressed conspecific. Learn. Behav. 2015, 43, 289–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrtree, R.M.; Sayigh, L.S.; Williford, A.; Bocconcelli, A.; Curran, M.C.; Cox, T.M. First observed wild birth and acoustic record of a possible infanticide attempt on a common bottlenose dolphin (Tursiops truncatus). Mar. Mammal Sci. 2016, 32, 376–385. [Google Scholar] [CrossRef]
- Pace, D.; Mussi, B.; Miragliuolo, A.; Vivaldi, C.; Ardizzone, G. First record of a hagfish anchored to a living bottlenose dolphin in the Mediterranean Sea. J. Mammal. 2016, 97, 960–965. [Google Scholar] [CrossRef] [Green Version]
- Pace, D.S.; Giacomini, G.; Campana, I.; Paraboschi, M.; Pellegrino, G.; Silvestri, M.; Alessi, J.; Angeletti, D.; Cafaro, V.; Pavan, G.; et al. An integrated approach for cetacean knowledge and conservation in the Central Mediterranean Sea using research and social media data sources. Aquat. Conserv. Mar. Freshw. Ecosyst. 2019, 29, 1302–1323. [Google Scholar] [CrossRef]
- Pace, D.; Marco, C.; Giacomini, G.; Ferri, S.; Silvestri, M.; Papale, E.; Casoli, E.; Ventura, D.; Mingione, M.; Alaimo Di Loro, P.; et al. Capitoline dolphins: Residency patterns and abundance estimate of Tursiops truncatus at the Tiber River Estuary (Mediterranean Sea). Biology 2021, 10, 275. [Google Scholar] [CrossRef]
- Triossi, F.; Willi, D.; Pace, D.S. Occurrence of bottlenose dolphins Tursiops truncatus in natural gas fields of the Northwestern Adriatic Sea. Mar. Ecol. 2013, 34, 373–379. [Google Scholar] [CrossRef]
- Martino, S.; Pace, D.S.; Moro, S.; Casoli, E.; Ventura, D.; Frachea, A.; Silvestri, M.; Arcangeli, A.; Giacomini, G.; Ardizzone, G.; et al. Integration of presence-only data from several sources: A case study on dolphins’ spatial distribution. Ecography 2021, 44, 1533–1543. [Google Scholar] [CrossRef]
- Hall, T.W.; Higson, A.W.; Pierce, B.J.; Price, K.H.; Skousen, C.J. Haphazard sampling: Selection biases and the estimation consequences of these biases. Curr. Issues Audit. 2013, 7, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Mann, J. Behavioral sampling methods for cetaceans: A review and critique. Mar. Mammal Sci. 1999, 15, 102–122. [Google Scholar] [CrossRef]
- Lehner, P.N. Sampling methods in behavior research. Poult. Sci. 1992, 71, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Mariani, M.; Miragliuolo, A.; Mussi, B.; Russo, G.F.; Ardizzone, G.; Pace, D.S. Analysis of the natural markings of risso’s dolphins (Grampus griseus) in the Central Mediterranean Sea. J. Mammal. 2016, 97, 1512–1524. [Google Scholar] [CrossRef] [Green Version]
- Center for Conservation Bioacoustics. Raven Pro: Interactive Sound Analysis Software (Version 2.0). Ithaca, Cornell Laboratory of Ornithology, USA. 2019. Available online: https://ravensoundsoftware.com/ (accessed on 20 January 2022).
- Kriesell, H.J.; Elwen, S.H.; Nastasi, A.; Gridley, T. Identification and characteristics of signature whistles in wild bottlenose dolphins (Tursiops truncatus) from Namibia. PLoS ONE 2014, 9, e106317. [Google Scholar] [CrossRef]
- Luís, A.R.; Couchinho, M.N.; dos Santos, M.E. A quantitative analysis of pulsed signals emitted by wild bottlenose dolphins. PLoS ONE 2016, 11, e0157781. [Google Scholar] [CrossRef]
- Papale, E.; Alonge, G.; Caruso, F.; Grammauta, R.; Mazzola, S.; Mussi, B.; Pace, D.S.; Buscaino, G. The higher, the closer, the better? influence of sampling frequency and distance on the acoustic properties of short-beaked common dolphins burst pulses in the Mediterranean Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Luís, A.R.; Alves, I.S.; Sobreira, F.V.; Couchinho, M.N.; dos Santos, M.E. Brays and bits: Information theory applied to acoustic communication sequences of bottlenose dolphins. Bioacoustics 2019, 28, 286–296. [Google Scholar] [CrossRef]
- Janik, V.M.; King, S.L.; Sayigh, L.S.; Wells, R.S. Identifying signature whistles from recordings of groups of unrestrained bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 2013, 29, 109–122. [Google Scholar] [CrossRef]
- Stolen, M.K.; Barlow, J. A model life table for bottlenose dolphins (Tursiops truncatus) from the Indian River Lagoon System, Florida, USA. Mar. Mammal Sci. 2003, 19, 630–649. [Google Scholar] [CrossRef] [Green Version]
- Connor, R.C.; Wells, R.S.; Mann, J.; Read, A.J. The bottlenose dolphin: Social relationships in a fission-fusion society. In Cetacean Societies: Field Studies of Dolphins and Whales; Mann, J., Connor, R.C., Tyack, P.L., Whitehead, H., Eds.; The University of Chicago Press: Chicago, IL, USA, 2000; pp. 91–126. [Google Scholar]
- Hill, H.M.; Greer, T.; Solangi, M.; Kuczaj II, S.A. All mothers are not the same: Maternal styles in bottlenose dolphins (Tursiops truncatus). Int. J. Comp. Psychol. 2007, 20, 35–54. [Google Scholar]
- Alves, F.; Nicolau, C.; Dinis, A.; Ribeiro, C.; Freitas, L. Supportive behavior of free-ranging atlantic spotted dolphins (Stenella frontalis) toward dead neonates, with data on perinatal mortality. Acta Ethologica 2015, 18, 301–304. [Google Scholar] [CrossRef]
- King, B.J. When animals mourn. Sci. Am. 2013, 309, 62–67. [Google Scholar] [CrossRef]
- Fettermann, T.; Fiori, L.; Bader, M.; Doshi, A.; Breen, D.; Stockin, K.A.; Bollard, B. Behaviour reactions of bottlenose dolphins (Tursiops truncatus) to multirotor Unmanned Aerial Vehicles (UAVs). Sci. Rep. 2019, 9, 8558. [Google Scholar] [CrossRef] [Green Version]
- López, B.D.; López, A.; Methion, S.; Covelo, P. Infanticide attacks and associated epimeletic behaviour in free-ranging common bottlenose dolphins (Tursiops truncatus). J. Mar. Biol. Assoc. UK 2018, 98, 1159–1167. [Google Scholar] [CrossRef] [Green Version]
- Würsig, B.; Whitehead, H. Aerial behavior. In Encyclopedia of Marine Mammals, 2nd ed.; Perrin, W.F., Würsig, B., Thewissen, J.G.M., Eds.; Academic Press: London, UK, 2009; pp. 5–11. ISBN 978-0-12-373553-9. [Google Scholar]
- Quintana-Rizzo, E.; Wells, R. Behavior of an adult female bottlenose dolphin (Tursiops truncatus) toward an unrelated dead calf. Aquat. Mamm. 2016, 2016, 198–202. [Google Scholar] [CrossRef]
- Diaz-Aguirre, F.; Parra, G.J.; Passadore, C.; Möller, L. Kinship and reproductive condition correlate with affiliation patterns in female southern australian bottlenose dolphins. Sci. Rep. 2020, 10, 1891. [Google Scholar] [CrossRef] [Green Version]
- Fury, C.; Harrison, P. Abundance, site fidelity and range patterns of indo-pacific bottlenose dolphins (Tursiops aduncus) in two australian subtropical estuaries. Mar. Freshw. Res. 2008, 59, 1015–1027. [Google Scholar] [CrossRef]
- Rossman, S.; McCabe, E.B.; Barros, N.B.; Gandhi, H.; Ostrom, P.H.; Stricker, C.A.; Wells, R.S. Foraging habits in a generalist predator: Sex and age influence habitat selection and resource use among bottlenose dolphins (Tursiops truncatus). Mar. Mammal Sci. 2015, 31, 155–168. [Google Scholar] [CrossRef]
- Krasnova, V.; Chernetsky, A.; Zheludkova, A.; Bel’kovich, V. Parental behavior of the beluga whale (Delphinapterus leucas) in natural environment. Biol. Bull. 2014, 41, 349–356. [Google Scholar] [CrossRef]
- Cockcroft, V.G.; Sauer, W. Observed and inferred epimeletic (nurturant) behaviour in bottlenose dolphins. Aquat. Mamm. 1990, 16, 31–32. [Google Scholar]
- Lodi, L. Epimeletic behavior of free-ranging rough-toothed dolphins, Steno bredanensis, from Brazil. Mar. Mammal Sci. 2006, 8, 284–287. [Google Scholar] [CrossRef]
- Jones, G.J.; Sayigh, L.S. Geographic variation in rates of vocal production of free-ranging bottlenose dolphins. Mar. Mammal Sci. 2002, 18, 374–393. [Google Scholar] [CrossRef]
- La Manna, G.; Rako-Gospić, N.; Sarà, G.; Gatti, F.; Bonizzoni, S.; Ceccherelli, G. Whistle variation in mediterranean common bottlenose dolphin: The role of geographical, anthropogenic, social, and behavioral factors. Ecol. Evol. 2020, 10, 1971–1987. [Google Scholar] [CrossRef] [PubMed]
- May-Collado, L.J. Changes in whistle structure of two dolphin species during interspecific associations. Ethology 2010, 116, 1065–1074. [Google Scholar] [CrossRef]
- Papale, E.; Azzolin, M.; Cascão, I.; Gannier, A.; Lammers, M.O.; Martin, V.M.; Oswald, J.; Perez-Gil, M.; Prieto, R.; Silva, M.A.; et al. Acoustic divergence between bottlenose dolphin whistles from the Central–Eastern North Atlantic and Mediterranean Sea. Acta Ethologica 2014, 17, 155–165. [Google Scholar] [CrossRef]
- Herzing, D. Vocalizations and associated underwater behavior of free-ranging atlantic spotted dolphins, Stenella frontalis and bottlenose dolphins, Tursiops truncatus. Aquat. Mamm. 1996, 22, 61–79. [Google Scholar]
- Smolker, R.A.; Mann, J.; Smuts, B.B. Use of signature whistles during separations and reunions by wild bottlenose dolphin mothers and infants. Behav. Ecol. Sociobiol. 1993, 33, 393–402. [Google Scholar] [CrossRef]
- Herzing, D. Synchronous and rhythmic vocalizations and correlated underwater behavior of free-ranging atlantic spotted dolphins (Stenella frontalis) and bottlenose dolphins (Tursiops truncatus) in the Bahamas. Anim. Behav. Cogn. 2015, 2, 14–29. [Google Scholar] [CrossRef]
- Dos Santos, M.E.; Caporin, G.; Moreira, H.O.; Ferreira, A.J.; Coelho, J.L.B. Acoustic behavior in a local population of bottlenose dolphins. In Sensory Abilities of Cetaceans: Laboratory and Field Evidence; Thomas, J.A., Kastelein, R.A., Eds.; NATO ASI Series; Springer: Boston, MA, USA, 1990; pp. 585–598. ISBN 978-1-4899-0858-2. [Google Scholar]
- Snijders, L.; Blumstein, D.T.; Stanley, C.R.; Franks, D.W. Animal social network theory can help wildlife conservation. Trends Ecol. Evol. 2017, 32, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Pace, D.S.; Arcangeli, A.; Mussi, B.; Vivaldi, C.; Ledon, C.; Lagorio, S.; Giacomini, G.; Pavan, G.; Ardizzone, G. Habitat suitability modeling in different sperm whale social groups. J. Wildl. Manag. 2018, 82, 1062–1073. [Google Scholar] [CrossRef]
- Bolaños-Jiménez, J.; Morteo, E.; Fruet, P.F.; Delfìn-Alfonso, C.A.; Secchi, E.R.; Bello-Pineda, J. Seasonal population parameters reveal sex-related dynamics of common bottlenose dolphins in open waters of the southwestern Gulf of Mexico. Mar. Mammal Sci. 2021, 1–20. [Google Scholar] [CrossRef]
- Pace, D.S.; Miragliuolo, A.; Mariani, M.; Vivaldi, C.; Mussi, B. Sociality of sperm whale off Ischia Island (Tyrrhenian Sea, Italy). Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 71–82. [Google Scholar] [CrossRef]
- Pace, D.S.; Mussi, B.; Gordon, J.C.; Wurtz, M. Foreword. Aquat. Conserv. Mar. Freshw. Ecosyst. 2014, 24, 1–3. [Google Scholar] [CrossRef]
- Frau, S.; Ronchetti, F.; Perretti, F.; Addis, A.; Ceccherelli, G.; La Manna, G. The influence of fish farm activity on the social structure of the common bottlenose dolphin in Sardinia (Italy). PeerJ 2021, 9, e10960. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vocalization | Spectrogram Parameters (Raven Pro 1.6) |
|---|---|
| Whistles | Hamming window, size 1024, DFT 1024, Overlap 50%, Hop size 512 |
| Click trains/Burst Pulses | Hamming window, size 512, DFT 512, Overlap 50%, Hop size 256 |
| Bray-call elements | Hann window, size 2048, DFT 2048, Overlap 50%, Hop size 1024 |
| Whistles | Definition |
| Minimum Frequency (Hz) | Frequency at the lower limit of the whistle |
| Maximum Frequency (Hz) | Frequency at the upper limit of the whistle |
| Frequency Range (Hz) | Maximum frequency-minimum frequency |
| Start Frequency (Hz) | Frequency at the beginning of the whistle |
| End Frequency (Hz) | Frequency at the end of the whistle |
| Duration (s) | Total duration calculated as: ending time-beginning time |
| Number of Inflection Points | Mathematic definition in sine function of a change from positive to negative or negative to positive slope |
| Harmonics (presence/absence) | Presence of multiples of the fundamental frequency |
| Step (presence/absence) | Abrupt discontinuity in frequency |
| Interruptions (presence/absence) | Abrupt discontinuity in time |
| Click trains/Burst Pulses | Definition |
| Number of pulses | Number of pulses composing the sound |
| Duration (s) | Time from the first click to the last click |
| Repetition Rate (pulse/second) | Number of pulses per second |
| Inter-click-interval (ms) | Inverse of the repetition rate |
| Whistle Category | Maximum Frequency (Hz) | Minimum Frequency (Hz) | Frequency Range (Hz) | Start Frequency (Hz) | End Frequency (Hz) | Duration (s) | Inflection Points (N) |
|---|---|---|---|---|---|---|---|
| OWs | 20507 ± 1794 | 7213 ± 1213 | 13294 ± 2472 | 9151 ± 2566 | 17802 ± 3759 | 0.98 ± 0.46 | 2.1 ± 1.7 |
| SW_1 | 21969 ± 1075 | 6851 ± 989 | 15118 ± 1450 | 7579 ± 1667 | 18928 ± 1792 | 1.38 ± 0.06 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pedrazzi, G.; Giacomini, G.; Pace, D.S. First Report of Epimeletic and Acoustic Behavior in Mediterranean Common Bottlenose Dolphins (Tursiops truncatus) Carrying Dead Calves. Biology 2022, 11, 337. https://doi.org/10.3390/biology11020337
Pedrazzi G, Giacomini G, Pace DS. First Report of Epimeletic and Acoustic Behavior in Mediterranean Common Bottlenose Dolphins (Tursiops truncatus) Carrying Dead Calves. Biology. 2022; 11(2):337. https://doi.org/10.3390/biology11020337
Chicago/Turabian StylePedrazzi, Giulia, Giancarlo Giacomini, and Daniela Silvia Pace. 2022. "First Report of Epimeletic and Acoustic Behavior in Mediterranean Common Bottlenose Dolphins (Tursiops truncatus) Carrying Dead Calves" Biology 11, no. 2: 337. https://doi.org/10.3390/biology11020337
APA StylePedrazzi, G., Giacomini, G., & Pace, D. S. (2022). First Report of Epimeletic and Acoustic Behavior in Mediterranean Common Bottlenose Dolphins (Tursiops truncatus) Carrying Dead Calves. Biology, 11(2), 337. https://doi.org/10.3390/biology11020337