
Advanced Diffusion MRI of the Visual System in Glaucoma: From Experimental Animal Models to Humans


Abstract
:Simple Summary
Abstract
1. Introduction
Magnetic Resonance Imaging and Principles of Diffusion
2. Materials and Methods
2.1. Literature Search
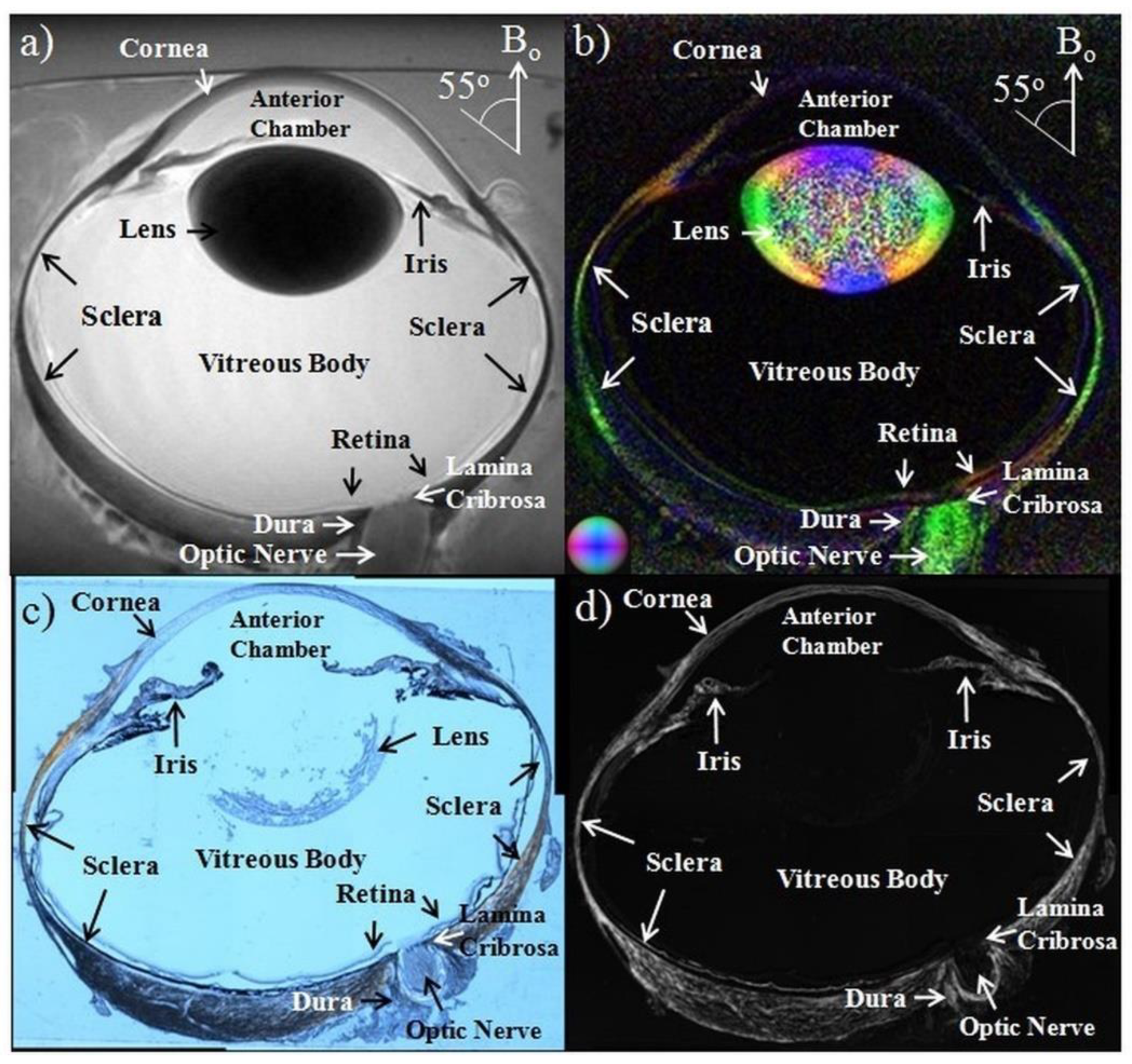
2.2. Anatomical MRI in Glaucona
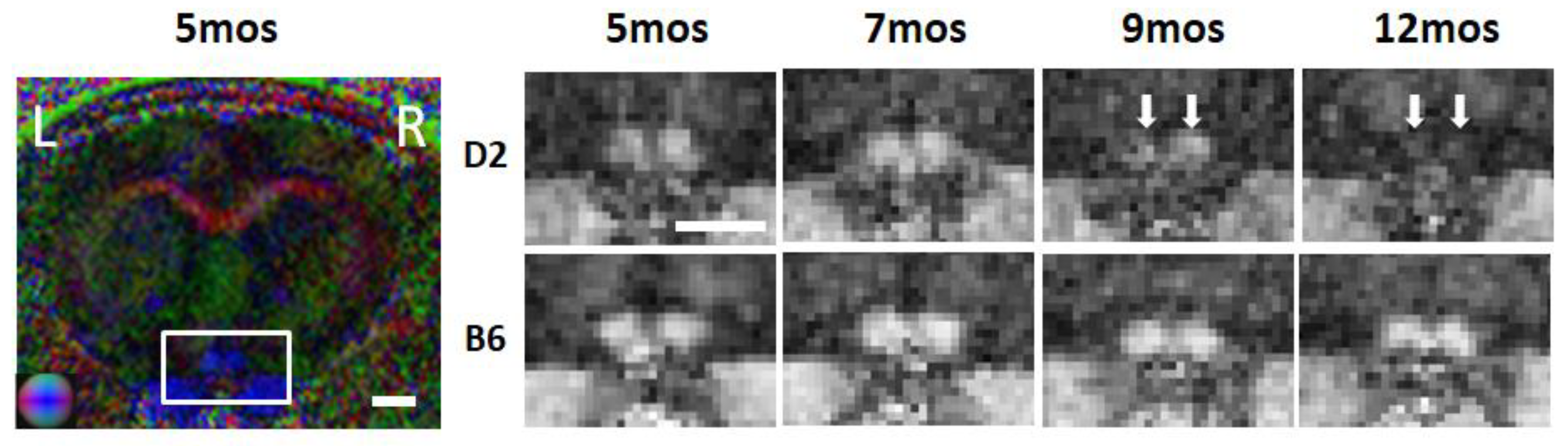
2.3. Diffusion-Weighted Imaging (DWI) in Glaucoma
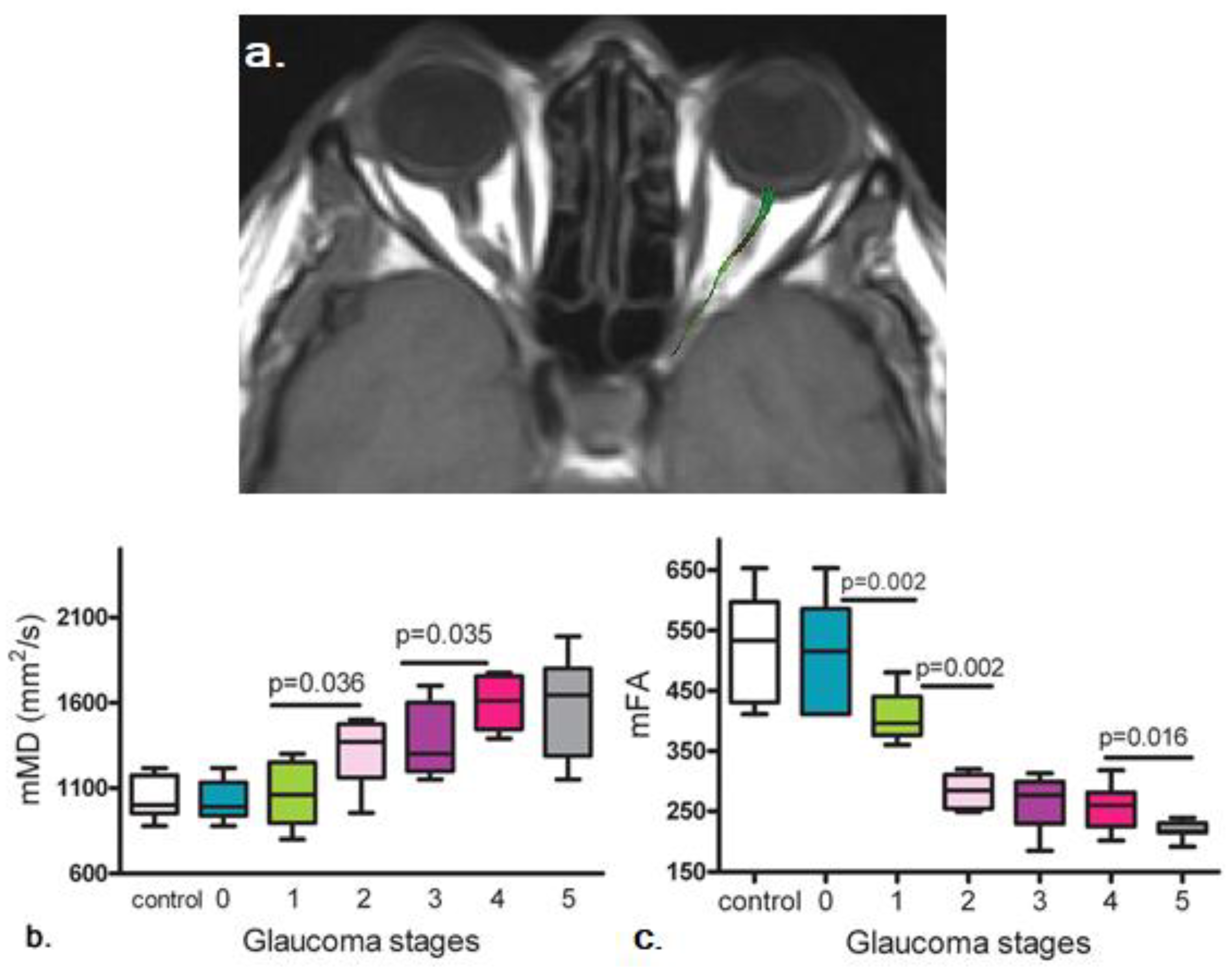
2.4. Diffusion Tensor Imaging (DTI) in Glaucoma
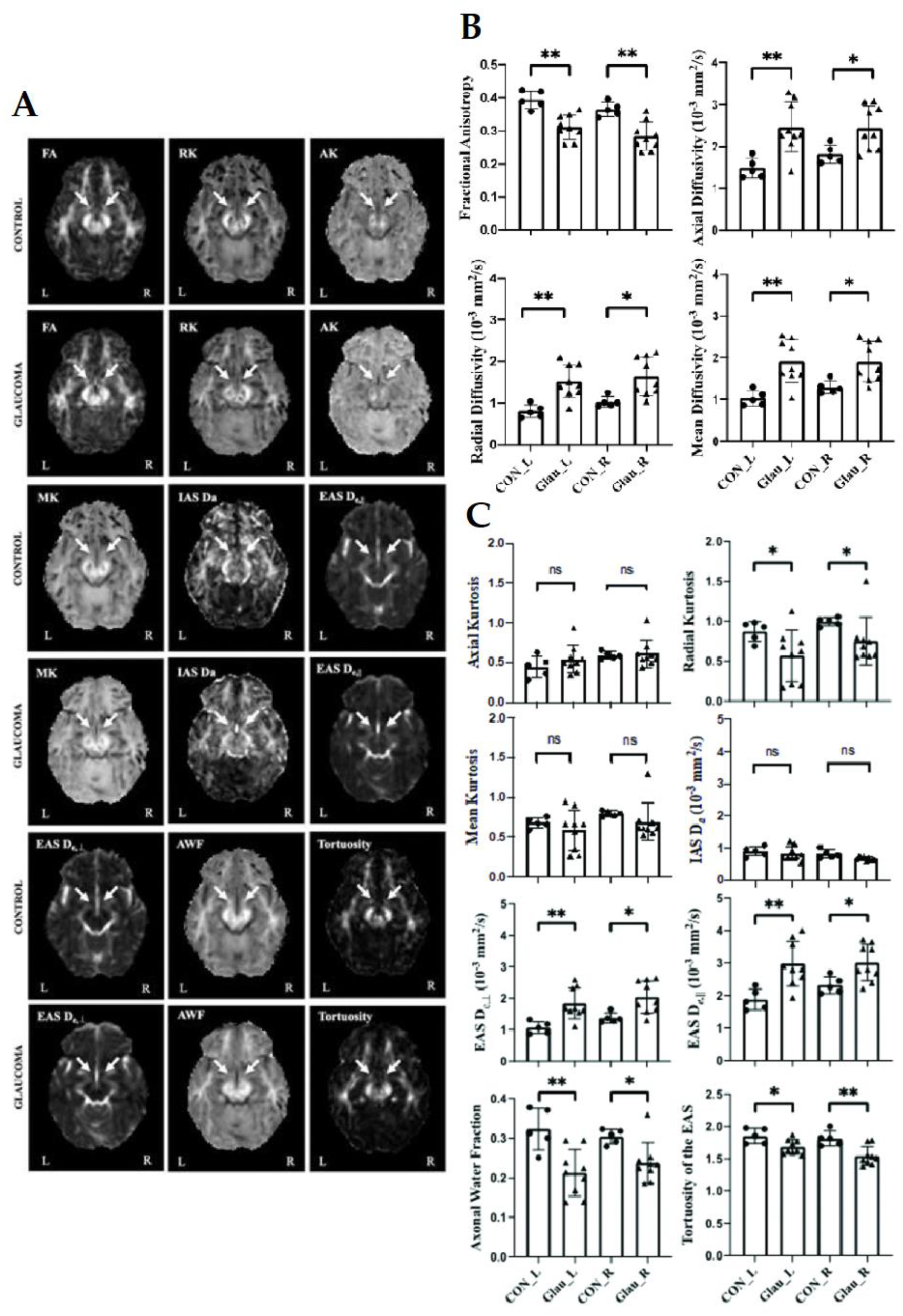
2.5. Diffusion Kurtosis Imaging (DKI) in Glaucoma
2.6. White Matter Tract Integrity (WMTI) Model in Glaucoma
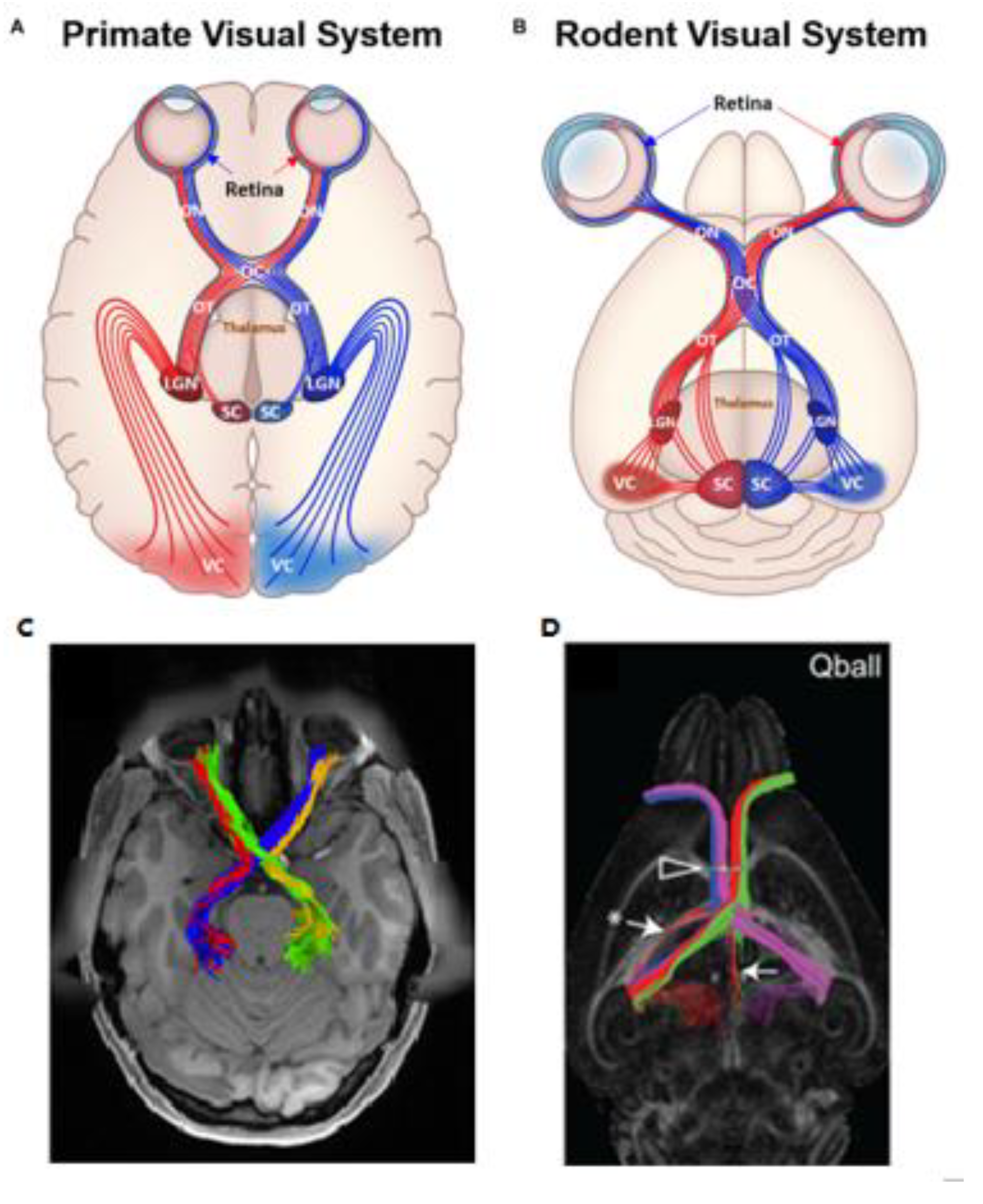
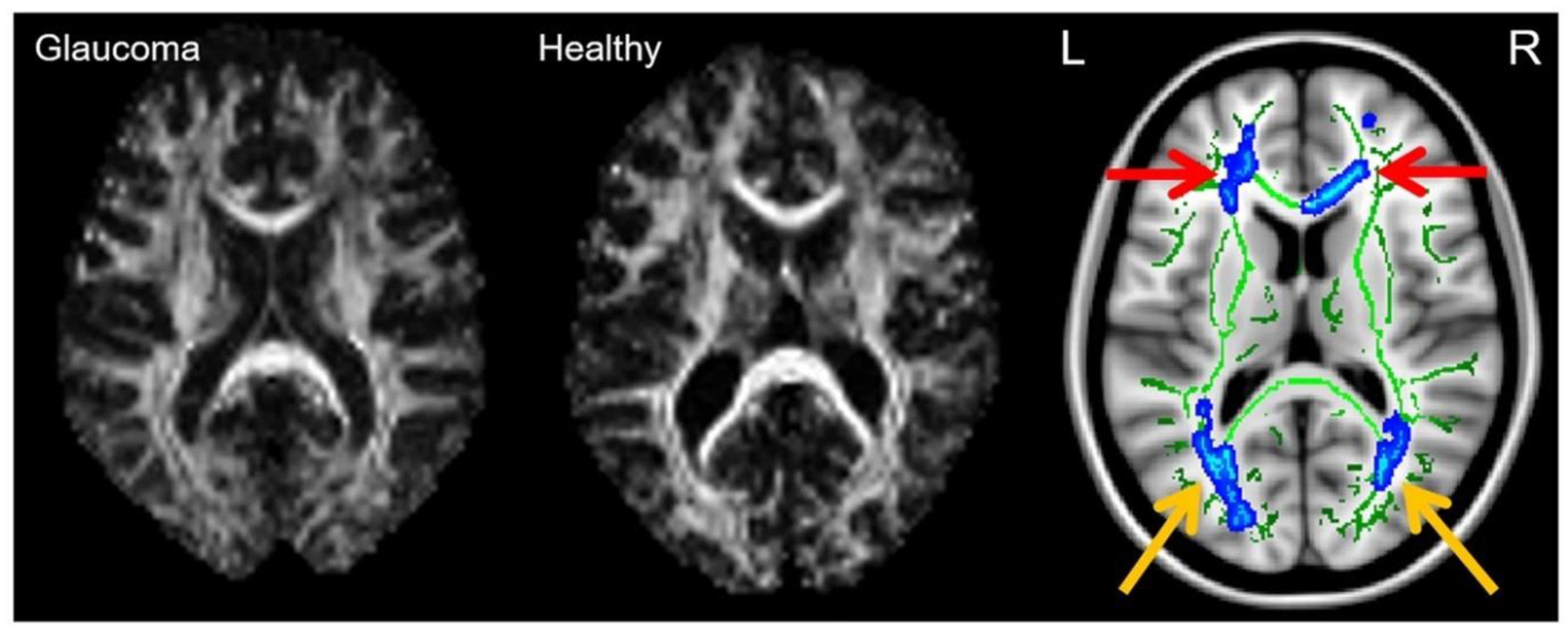
2.7. Diffusion MRI Tractography in Glaucoma
3. Challenges and Limitations
3.1. Limitations of the Literature Review Methods
3.2. Limited Specificity from DWI and DTI Results
3.3. Imaging Quality
3.4. Reproducibility
4. Future Prospective and Opportunities
4.1. Exploring Comorbid Conditions
4.2. Development of Clinically-Feasible Imaging Sequences
4.3. Glaucoma Detection through a Combination of Imaging Techniques
4.4. Evaluation of Neurotherapeutic Options for Glaucoma
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tham, Y.-C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.-Y. Global Prevalence of Glaucoma and Projections of Glaucoma Burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Berdahl, J.P.; Allingham, R.R. Intracranial pressure and glaucoma. Curr. Opin. Ophthalmol. 2010, 21, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Kapetanakis, V.V.; Chan, M.P.Y.; Foster, P.; Cook, D.; Owen, C.; Rudnicka, A. Global variations and time trends in the prevalence of primary open angle glaucoma (POAG): A systematic review and meta-analysis. Br. J. Ophthalmol. 2015, 100, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A. Glaucoma. Lancet 2011, 377, 1367–1377. [Google Scholar] [CrossRef]
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The Pathophysiology and Treatment of Glaucoma: A review. JAMA—J. Am. Med. Assoc. 2014, 311, 1901–1911. [Google Scholar] [CrossRef] [Green Version]
- Doucette, L.P.; Rasnitsyn, A.; Seifi, M.; Walter, M.A. The interactions of genes, age, and environment in glaucoma pathogenesis. Surv. Ophthalmol. 2015, 60, 310–326. [Google Scholar] [CrossRef]
- Wostyn, P. Glaucoma as a dangerous interplay between ocular fluid and cerebrospinal fluid. Med. Hypotheses 2019, 127, 97–99. [Google Scholar] [CrossRef]
- Bonomi, L.; Marchini, G.; Marraffa, M.; Bernardi, P.; De Franco, I.; Perfetti, S.; Varotto, A.; Tenna, V. Prevalence of glaucoma and intraocular pressure distribution in a defined population: The Egna-Neumarkt study. Ophthalmology 1998, 105, 209–215. [Google Scholar] [CrossRef]
- Klein, B.E.K.; Klein, R.; Sponsel, W.E.; Franke, T.; Cantor, L.B.; Martone, J.; Menage, M.J. Prevalence of Glaucoma: The Beaver Dam Eye Study. Ophthalmology 1992, 99, 1499–1504. [Google Scholar] [CrossRef]
- Van der Merwe, Y.; Murphy, M.C.; Sims, J.R.; Faiq, M.A.; Yang, X.-L.; Ho, L.C.; Conner, I.P.; Yu, Y.; Leung, C.K.; Wollstein, G.; et al. Citicoline Modulates Glaucomatous Neurodegeneration Through Intraocular Pressure-Independent Control. Neurotherapeutics 2021, 18, 1339–1359. [Google Scholar] [CrossRef]
- Rudnicka, A.R.; Mt-Isa, S.; Owen, C.; Cook, D.; Ashby, D. Variations in Primary Open-Angle Glaucoma Prevalence by Age, Gender, and Race: A Bayesian Meta-Analysis. Investig. Opthalmol. Vis. Sci. 2006, 47, 4254–4261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.-L.; Van Der Merwe, Y.; Sims, J.; Parra, C.; Ho, L.C.; Schuman, J.S.; Wollstein, G.; Lathrop, K.L.; Chan, K.C. Age-related Changes in Eye, Brain and Visuomotor Behavior in the DBA/2J Mouse Model of Chronic Glaucoma. Sci. Rep. 2018, 8, 4643. [Google Scholar] [CrossRef] [PubMed]
- Rezaie, T.; Child, A.; Hitchings, R.; Brice, G.; Miller, L.; Coca-Prados, M.; Héon, E.; Krupin, T.; Ritch, R.; Kreutzer, D.; et al. Adult-Onset Primary Open-Angle Glaucoma Caused by Mutations in Optineurin. Science 2002, 295, 1077–1079. [Google Scholar] [CrossRef] [PubMed]
- Wolfs, R.C.W.; Klaver, C.C.W.; Ramrattan, R.S.; van Duijn, C.M.; Hofman, A.; de Jong, P.T.V.M. Genetic Risk of Primary Open-angle Glaucoma. Arch. Ophthalmol. 1998, 116, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, M.; Kanamori, A.; Negi, A. Diabetes mellitus as a Risk Factor for Glaucomatous Optic Neuropathy. Ophthalmologica 2005, 219, 1–10. [Google Scholar] [CrossRef]
- Mitchell, P.; Hourihan, F.; Sandbach, J.; Wang, J.J. The relationship between glaucoma and myopia: The blue mountains eye study. Ophthalmology 1999, 106, 2010–2015. [Google Scholar] [CrossRef]
- Quigley, H.A.; Broman, A.T. The number of people with glaucoma worldwide in 2010 and 2020. Br. J. Ophthalmol. 2006, 90, 262–267. [Google Scholar] [CrossRef] [Green Version]
- Kamińska, A.; Romano, G.; Rejdak, R.; Zweifel, S.; Fiedorowicz, M.; Rejdak, M.; Bajka, A.; Amato, R.; Bucolo, C.; Avitabile, T.; et al. Influence of trace elements on neurodegenerative diseases of the eye—The glaucoma model. Int. J. Mol. Sci. 2021, 22, 4323. [Google Scholar] [CrossRef]
- Chow, L.S.; Paley, M.N. Recent advances on optic nerve magnetic resonance imaging and post-processing. Magn. Reson. Imaging 2021, 79, 76–84. [Google Scholar] [CrossRef]
- Posarelli, C.; Ortenzio, P.; Ferreras, A.; Toro, M.D.; Passani, A.; Loiudice, P.; Oddone, F.; Casini, G.; Figus, M. Twenty-Four-Hour Contact Lens Sensor Monitoring of Aqueous Humor Dynamics in Surgically or Medically Treated Glaucoma Patients. J. Ophthalmol. 2019, 2019, 9890831. [Google Scholar] [CrossRef] [Green Version]
- Schacknow, P.N.; Samples, J.R. The Glaucoma Book: A Practical, Evidence-Based Approach to Patient Care; Springer: New York, NY, USA, 2010. [Google Scholar]
- Jonas, J.B.; Müller-Bergh, J.A.; Schlötzer-Schrehardt, U.M.; Naumann, G.O. Histomorphometry of the human optic nerve. Investig. Ophthalmol. Vis. Sci. 1990, 31, 736–744. [Google Scholar]
- Shin, J.W.; Sung, K.R.; Park, S.-W. Patterns of Progressive Ganglion Cell–Inner Plexiform Layer Thinning in Glaucoma Detected by OCT. Ophthalmology 2018, 125, 1515–1525. [Google Scholar] [CrossRef] [PubMed]
- Leung, C.K.-S.; Chan, W.-M.; Hui, Y.-L.; Yung, W.H.; Woo, J.; Tsang, M.-K.; Tse, K.-K. Analysis of Retinal Nerve Fiber Layer and Optic Nerve Head in Glaucoma with Different Reference Plane Offsets, Using Optical Coherence Tomography. Investig. Opthalmol. Vis. Sci. 2005, 46, 891–899. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouri-Mahdavi, K.; Weiss, R.E. Detection of Glaucoma Deterioration in the Macular Region with Optical Coherence Tomography: Challenges and Solutions. Am. J. Ophthalmol. 2021, 222, 277–284. [Google Scholar] [CrossRef]
- Jindal, V. Glaucoma: An Extension of Various Chronic Neurodegenerative Disorders. Mol. Neurobiol. 2013, 48, 186–189. [Google Scholar] [CrossRef]
- Chen, Z.; Lin, F.; Wang, J.; Li, Z.; Dai, H.; Mu, K.; Ge, J.; Zhang, H. Diffusion tensor magnetic resonance imaging reveals visual pathway damage that correlates with clinical severity in glaucoma. Clin. Exp. Ophthalmol. 2013, 41, 43–49. [Google Scholar] [CrossRef]
- Lawlor, M.; Danesh-Meyer, H.; Levin, L.A.; Davagnanam, I.; De Vita, E.; Plant, G.T. Glaucoma and the brain: Trans-synaptic degeneration, structural change, and implications for neuroprotection. Surv. Ophthalmol. 2018, 63, 296–306. [Google Scholar] [CrossRef]
- Sponsel, W.E.; Groth, S.L.; Satsangi, N.; Maddess, T.; Reilly, M. Refined Data Analysis Provides Clinical Evidence for Central Nervous System Control of Chronic Glaucomatous Neurodegeneration. Transl. Vis. Sci. Technol. 2014, 3, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Skaat, A.; Muylaert, S.; Mogil, R.S.; Furlanetto, R.L.; Netto, C.F.; Banik, R.; Liebmann, J.M.; Ritch, R.; Park, S.C. Relationship Between Optic Nerve Head Drusen Volume and Structural and Functional Optic Nerve Damage. J. Glaucoma 2017, 26, 1095–1100. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Li, J.; Xu, L.; Zhang, L.; Wang, Z.C.; Yang, H.; Chen, C.X.; Wu, X.S.; Jonas, J.B. Anterior visual pathway assessment by magnetic resonance imaging in normal-pressure glaucoma. Acta Ophthalmol. 2012, 90, e295–e302. [Google Scholar] [CrossRef]
- Brown, H.D.H.; Woodall, R.L.; Kitching, R.E.; Baseler, H.A.; Morland, A.B. Using magnetic resonance imaging to assess visual deficits: A review. Ophthalmic Physiol. Opt. 2016, 36, 240–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
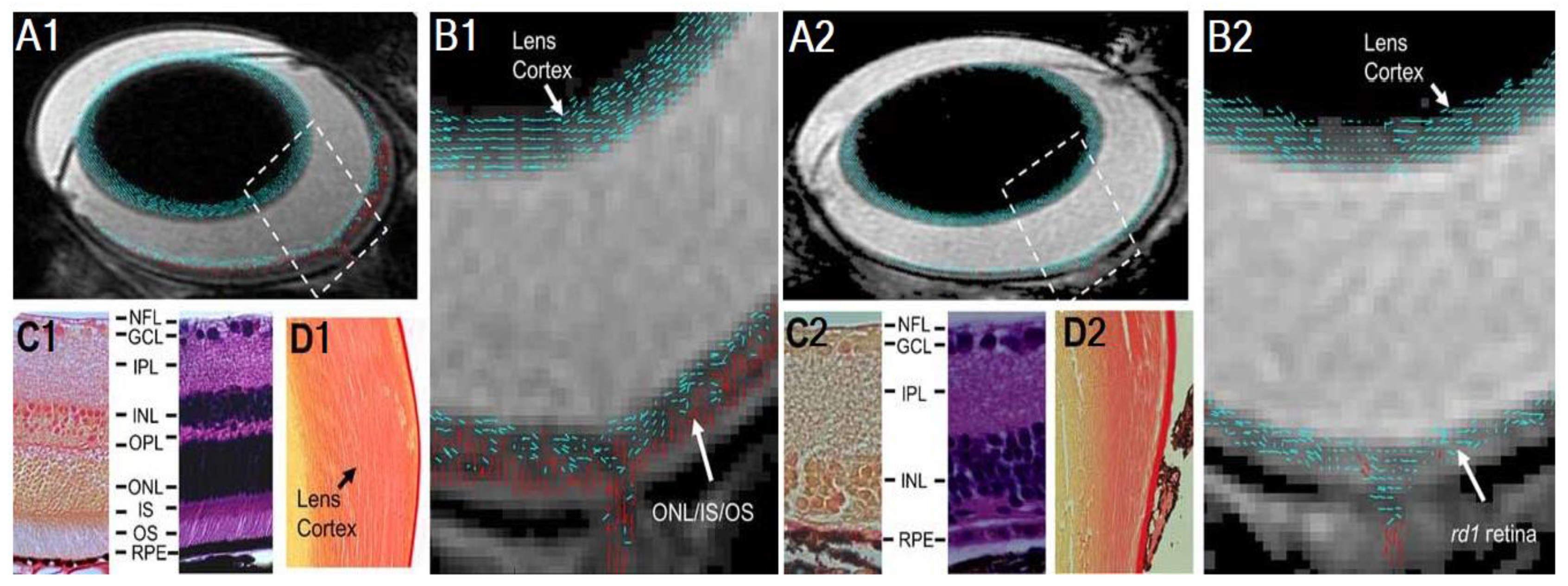
- Cheng, H.; Nair, G.; Walker, T.A.; Kim, M.K.; Pardue, M.T.; Thulé, P.M.; Olson, D.E.; Duong, T.Q. Structural and functional MRI reveals multiple retinal layers. Proc. Natl. Acad. Sci. USA 2006, 103, 17525–17530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; De La Garza, B.; Shih, Y.-Y.I.; Muir, E.R.; Duong, T.Q. Layer-specific blood-flow MRI of retinitis pigmentosa in RCS rats. Exp. Eye Res. 2012, 101, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smirnakis, S.M.; Brewer, A.A.; Schmid, M.; Tolias, A.S.; Schüz, A.; Augath, M.; Inhoffen, W.; Wandell, B.A.; Logothetis, N.K. Lack of long-term cortical reorganization after macaque retinal lesions. Nature 2005, 435, 300–307. [Google Scholar] [CrossRef]
- Deng, W.; Faiq, M.A.; Liu, C.; Adi, V.; Chan, K.C.W. Applications of Manganese-Enhanced Magnetic Resonance Imaging in Ophthalmology and Visual Neuroscience. Front. Neural Circuits 2019, 13, 35. [Google Scholar] [CrossRef]
- He, J.; Zhang, F.; Xie, G.; Yao, S.; Feng, Y.; Bastos, D.C.A.; Rathi, Y.; Makris, N.; Kikinis, R.; Golby, A.J.; et al. Comparison of multiple tractography methods for reconstruction of the retinogeniculate visual pathway using diffusion MRI. Hum. Brain Mapp. 2021, 42, 3887–3904. [Google Scholar] [CrossRef]
- Moldrich, R.X.; Pannek, K.; Hoch, R.; Rubenstein, J.L.; Kurniawan, N.D.; Richards, L.J. Comparative mouse brain tractography of diffusion magnetic resonance imaging. NeuroImage 2010, 51, 1027–1036. [Google Scholar] [CrossRef] [Green Version]
- Dai, A.I.; Saygili, O. Primary congenital glaucoma in infants with abnormal brain MRI findings. Ann. Saudi Med. 2007, 27, 264–267. [Google Scholar] [CrossRef]
- Williams, A.L.; Lackey, J.; Wizov, S.S.; Chia, T.M.T.; Gatla, S.; Moster, M.L.; Sergott, R.C.; Spaeth, G.L.; Lai, S. Evidence for Widespread Structural Brain Changes in Glaucoma: A Preliminary Voxel-Based MRI Study. Investig. Opthalmol. Vis. Sci. 2013, 54, 5880–5887. [Google Scholar] [CrossRef] [Green Version]
- Lestak, J.; Tintera, J.; Svata, Z.; Ettler, L.; Rozsival, P. Glaucoma and CNS. Comparison of fMRI results in high tension and normal tension glaucoma. Biomed. Pap. 2014, 158, 144–153. [Google Scholar] [CrossRef]
- Lee, J.Y.; Jeong, H.J.; Lee, J.H.; Kim, Y.J.; Kim, E.Y.; Kim, Y.Y.; Ryu, T.; Cho, Z.-H.; Kim, Y.-B. An Investigation of Lateral Geniculate Nucleus Volume in Patients With Primary Open-Angle Glaucoma Using 7 Tesla Magnetic Resonance Imaging. Investig. Opthalmol. Vis. Sci. 2014, 55, 3468–3476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, M.; Omodaka, K.; Tatewaki, Y.; Himori, N.; Matsudaira, I.; Nishiguchi, K.M.; Murata, T.; Taki, Y.; Nakazawa, T. Quantitative MRI evaluation of glaucomatous changes in the visual pathway. PLoS ONE 2018, 13, e0197027. [Google Scholar] [CrossRef] [PubMed]
- Demer, J.L.; Clark, R.A.; Suh, S.Y.; Giaconi, J.A.; Nouri-Mahdavi, K.; Law, S.K.; Bonelli, L.; Coleman, A.L.; Caprioli, J. Magnetic Resonance Imaging of Optic Nerve Traction During Adduction in Primary Open-Angle Glaucoma With Normal Intraocular Pressure. Investig. Opthalmol. Vis. Sci. 2017, 58, 4114–4125. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, T.; Sabel, B.A.; Chen, Z.; Wen, H.; Li, J.; Xie, X.; Yang, D.; Chen, W.; Wang, N.; et al. Structural brain alterations in primary open angle glaucoma: A 3T MRI study. Sci. Rep. 2016, 6, 18969. [Google Scholar] [CrossRef]
- Jelescu, I.O.; Veraart, J.; Adisetiyo, V.; Milla, S.S.; Novikov, D.; Fieremans, E. One diffusion acquisition and different white matter models: How does microstructure change in human early development based on WMTI and NODDI? NeuroImage 2015, 107, 242–256. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Sun, S.-W.; Naismith, R.T.; Snyder, A.Z.; Cross, A.; Song, S.-K. Assessing optic nerve pathology with diffusion MRI: From mouse to human. NMR Biomed. 2008, 21, 928–940. [Google Scholar] [CrossRef] [Green Version]
- Chilla, G.S.; Tan, C.H.; Xu, C.; Poh, C.L. Diffusion weighted magnetic resonance imaging and its recent trend—A survey. Quant. Imaging Med. Surg. 2015, 5, 407–422. [Google Scholar] [CrossRef]
- Posse, S.; Cuenod, A.C.; Le Bihan, D. Human brain: Proton diffusion MR spectroscopy. Radiology 1993, 188, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.-Y.; Poon, Y.-Y.; Chan, J.Y.H.; Chan, S.H.H. Baroreflex functionality in the eye of diffusion tensor imaging. J. Physiol. 2018, 597, 41–55. [Google Scholar] [CrossRef]
- Kasi, A.; Faiq, M.A.; Chan, K.C. In vivo imaging of structural, metabolic and functional brain changes in glaucoma. Neural Regen. Res. 2019, 14, 446–449. [Google Scholar] [CrossRef]
- Fiedorowicz, M.; Wełniak-Kamińska, M.; Świątkiewicz, M.; Orzeł, J.; Chorągiewicz, T.; Toro, M.D.; Rejdak, R.; Bogorodzki, P.; Grieb, P. Changes of Ocular Dimensions as a Marker of Disease Progression in a Murine Model of Pigmentary Glaucoma. Front. Pharmacol. 2020, 11, 1377. [Google Scholar] [CrossRef] [PubMed]
- Kashiwagi, K.; Okubo, T.; Tsukahara, S. Association of Magnetic Resonance Imaging of Anterior Optic Pathway with Glaucomatous Visual Field Damage and Optic Disc Cupping. J. Glaucoma 2004, 13, 189–195. [Google Scholar] [CrossRef] [PubMed]
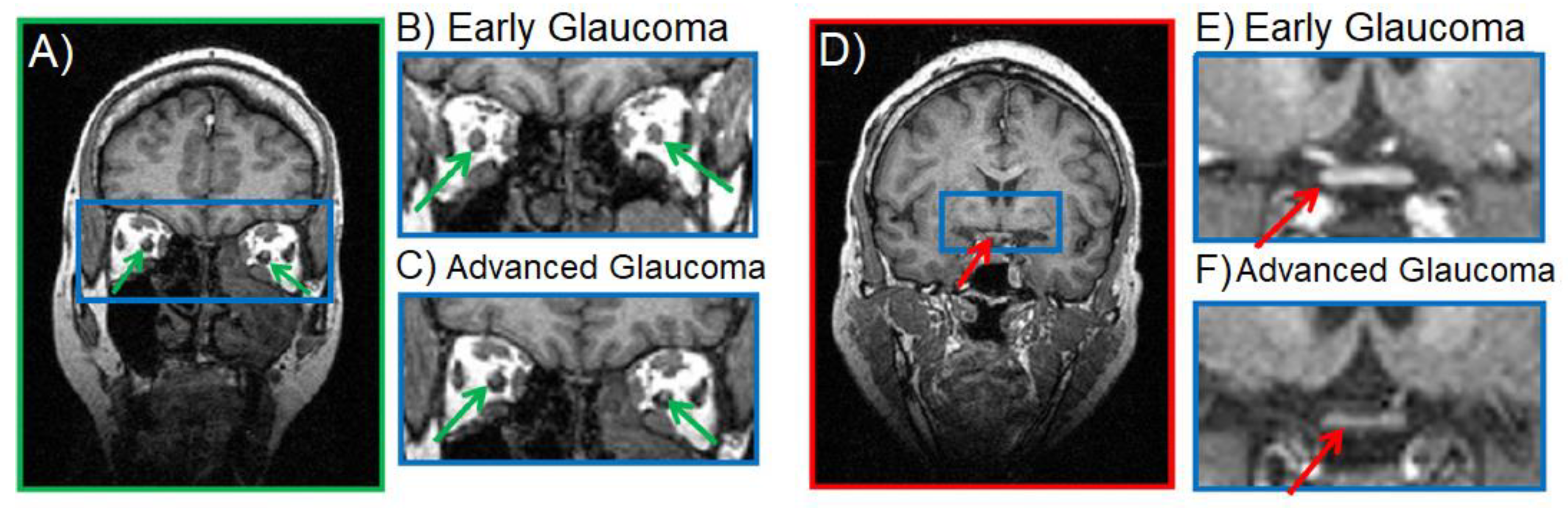
- Dai, H.; Mu, K.; Qi, J.; Wang, C.; Zhu, W.; Xia, L.; Chen, Z.; Zhang, H.; Ai, F.; Morelli, J. Assessment of Lateral Geniculate Nucleus Atrophy with 3T MR Imaging and Correlation with Clinical Stage of Glaucoma. Am. J. Neuroradiol. 2011, 32, 1347–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, N.; Greenberg, G.; De Tilly, L.N.; Gray, B.; Polemidiotis, M.; Yucel, Y.H. Atrophy of the lateral geniculate nucleus in human glaucoma detected by magnetic resonance imaging. Br. J. Ophthalmol. 2008, 93, 56–60. [Google Scholar] [CrossRef]
- Gupta, N.; Ang, L.-C.; De Tilly, L.N.; Bidaisee, L.; Yücel, Y.H. Human glaucoma and neural degeneration in intracranial optic nerve, lateral geniculate nucleus, and visual cortex. Br. J. Ophthalmol. 2006, 90, 674–678. [Google Scholar] [CrossRef]
- Yu, L.; Xie, B.; Yin, X.; Liang, M.; Evans, A.C.; Wang, J.; Dai, C. Reduced Cortical Thickness in Primary Open-Angle Glaucoma and Its Relationship to the Retinal Nerve Fiber Layer Thickness. PLoS ONE 2013, 8, e73208. [Google Scholar] [CrossRef] [Green Version]
- Yücel, Y.H.; Zhang, Q.; Gupta, N.; Kaufman, P.L.; Weinreb, R.N. Loss of Neurons in Magnocellular and Parvocellular Layers of the Lateral Geniculate Nucleus in Glaucoma. Arch. Ophthalmol. 2000, 118, 378–384. [Google Scholar] [CrossRef]
- Manogaran, P.; Samardzija, M.; Schad, A.N.; Wicki, C.A.; Walker-Egger, C.; Rudin, M.; Grimm, C.; Schippling, S. Retinal pathology in experimental optic neuritis is characterized by retrograde degeneration and gliosis. Acta Neuropathol. Commun. 2019, 7, 116. [Google Scholar] [CrossRef] [Green Version]
- Naismith, R.T.; Xu, J.; Tutlam, N.T.; Snyder, A.; Benzinger, T.; Shimony, J.; Shepherd, J.; Trinkaus, K.; Cross, A.H.; Song, S.-K. Disability in optic neuritis correlates with diffusion tensor-derived directional diffusivities. Neurology 2009, 72, 589–594. [Google Scholar] [CrossRef]
- Mori, S.; Zhang, J. Principles of Diffusion Tensor Imaging and Its Applications to Basic Neuroscience Research. Neuron 2006, 51, 527–539. [Google Scholar] [CrossRef] [Green Version]
- Fieremans, E.; Jensen, J.; Helpern, J.A. White matter characterization with diffusional kurtosis imaging. NeuroImage 2011, 58, 177–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meral, I.; Bilgili, Y. Diffusion Changes in the Vitreous Humor of the Eye during Aging. Am. J. Neuroradiol. 2011, 32, 1563–1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Wang, Q.; Chen, S.; Wickline, S.A.; Song, S.-K. In vivo diffusion tensor MRI of the mouse retina: A noninvasive visualization of tissue organization. NMR Biomed. 2010, 24, 447–451. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Sanz, M.; Galindo-Romero, C.; Valiente-Soriano, F.J.; Nadal-Nicolas, F.M.; Ortin-Martinez, A.; Rovere, G.; Salinas-Navarro, M.; Lucas-Ruiz, F.; Sanchez-Migallon, M.C.; Sobrado-Calvo, P.; et al. Shared and Differential Retinal Responses against Optic Nerve Injury and Ocular Hypertension. Front. Neurosci. 2017, 11, 235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagmann, P.; Jonasson, L.; Maeder, P.; Thiran, J.-P.; Wedeen, V.J.; Meuli, R. Understanding Diffusion MR Imaging Techniques: From Scalar Diffusion-weighted Imaging to Diffusion Tensor Imaging and Beyond. RadioGraphics 2006, 26, S205–S223. [Google Scholar] [CrossRef] [Green Version]
- Veraart, J.; Sijbers, J.; Sunaert, S.; Leemans, A.; Jeurissen, B. Weighted linear least squares estimation of diffusion MRI parameters: Strengths, limitations, and pitfalls. NeuroImage 2013, 81, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Soares, J.M.; Marques, P.; Alves, V.; Sousa, N. A hitchhiker’s guide to diffusion tensor imaging. Front. Neurosci. 2013, 7, 31. [Google Scholar] [CrossRef] [Green Version]
- Basser, P.J.; Pierpaoli, C. Microstructural and physiological features of tissues elucidated by quantitative-diffusion-tensor MRI. J. Magn. Reson. 2011, 213, 560–570. [Google Scholar] [CrossRef]
- Alexander, A.L.; Lee, J.E.; Lazar, M.; Field, A.S. Diffusion tensor imaging of the brain. Neurotherapeutics 2007, 4, 316–329. [Google Scholar] [CrossRef] [Green Version]
- Zikou, A.; Kitsos, G.; Tzarouchi, L.; Astrakas, L.; Alexiou, G.; Argyropoulou, M. Voxel-Based Morphometry and Diffusion Tensor Imaging of the Optic Pathway in Primary Open-Angle Glaucoma: A Preliminary Study. Am. J. Neuroradiol. 2011, 33, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Tellouck, L.; Durieux, M.; Coupé, P.; Cougnard-Grégoire, A.; Tellouck, J.; Tourdias, T.; Munsch, F.; Garrigues, A.; Helmer, C.; Malet, F.; et al. Optic radiations microstructural changes in glaucoma and association with severity: A study using 3tesla-magnetic resonance diffusion tensor imaging. Investig. Opthalmol. Vis. Sci. 2016, 57, 6539–6547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ho, L.C.; Wang, B.; Conner, I.P.; Van Der Merwe, Y.; Bilonick, R.A.; Kim, S.-G.; Wu, E.X.; Sigal, I.A.; Wollstein, G.; Schuman, J.S.; et al. In Vivo Evaluation of White Matter Integrity and Anterograde Transport in Visual System After Excitotoxic Retinal Injury with Multimodal MRI and OCT. Investig. Opthalmol. Vis. Sci. 2015, 56, 3788–3800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelhorn, T.; Michelson, G.; Waerntges, S.; Hempel, S.; El-Rafei, A.; Struffert, T.; Doerfler, A. A New Approach to Assess Intracranial White Matter Abnormalities in Glaucoma Patients: Changes of Fractional Anisotropy Detected by 3T Diffusion Tensor Imaging. Acad. Radiol. 2012, 19, 485–488. [Google Scholar] [CrossRef] [PubMed]
- Garaci, F.G.; Bolacchi, F.; Cerulli, A.; Melis, M.; Spanò, A.; Cedrone, C.; Floris, R.; Simonetti, G.; Nucci, C. Optic Nerve and Optic Radiation Neurodegeneration in Patients with Glaucoma: In Vivo Analysis with 3-T Diffusion-Tensor MR Imaging. Radiology 2009, 252, 496–501. [Google Scholar] [CrossRef] [Green Version]
- Colbert, M.K.; Ho, L.C.; van der Merwe, Y.; Yang, X.; McLellan, G.J.; Hurley, S.A.; Field, A.S.; Yun, H.; Du, Y.; Conner, I.P.; et al. Diffusion Tensor Imaging of Visual Pathway Abnormalities in Five Glaucoma Animal Models. Investig. Opthalmol. Vis. Sci. 2021, 62, 21. [Google Scholar] [CrossRef]
- Giorgio, A.; Zhang, J.; Costantino, F.; De Stefano, N.; Frezzotti, P. Diffuse brain damage in normal tension glaucoma. Hum. Brain Mapp. 2018, 39, 532–541. [Google Scholar] [CrossRef] [Green Version]
- Frezzotti, P.; Giorgio, A.; Motolese, I.; De Leucio, A.; Iester, M.; Motolese, E.; Federico, A.; De Stefano, N. Structural and Functional Brain Changes beyond Visual System in Patients with Advanced Glaucoma. PLoS ONE 2014, 9, e105931. [Google Scholar] [CrossRef] [Green Version]
- Qu, X.; Wang, Q.; Chen, W.; Li, T.; Guo, J.; Wang, H.; Zhang, X.; Wang, Y.; Wang, N.; Xian, J. Combined machine learning and diffusion tensor imaging reveals altered anatomic fiber connectivity of the brain in primary open-angle glaucoma. Brain Res. 2019, 1718, 83–90. [Google Scholar] [CrossRef]
- Sun, Z.; Parra, C.; Bang, J.W.; Fieremans, E.; Wollstein, G.; Schuman, J.S.; Chan, K.C. Diffusion Kurtosis Imaging Reveals Optic Tract Damage That Correlates with Clinical Severity in Glaucoma. In Proceedings of the 2020 42nd Annual International Conference of the IEEE Engineering in Medicine & Biology Society (EMBC), Montreal, QC, Canada, 20–24 July 2020; Volume 2020, pp. 1746–1749. [Google Scholar]
- Wang, R.; Tang, Z.; Sun, X.; Wu, L.; Wang, J.; Zhong, Y.; Xiao, Z. White matter abnormalities and correlation with severity in normal tension glaucoma: A whole brain atlas-based diffusion tensor study. Investig. Opthalmol. Vis. Sci. 2018, 59, 1313–1322. [Google Scholar] [CrossRef]
- Miller, N.; Liu, Y.; Krivochenitser, R.; Rokers, B. Linking neural and clinical measures of glaucoma with diffusion magnetic resonance imaging (dMRI). PLoS ONE 2019, 14, e0217011. [Google Scholar] [CrossRef] [Green Version]
- Chang, S.T.; Xu, J.; Trinkaus, K.; Pekmezci, M.; Arthur, S.N.; Song, S.-K.; Barnett, E.M. Optic nerve diffusion tensor imaging parameters and their correlation with optic disc topography and disease severity in adult glaucoma patients and controls. J. Glaucoma 2014, 23, 513–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mac Nair, C.E.; Nickells, R.W. Neuroinflammation in Glaucoma and Optic Nerve Damage. Prog. Mol. Biol. Transl. Sci. 2015, 134, 343–363. [Google Scholar] [PubMed]
- Lu, P.; Shi, L.; Du, H.; Xie, B.; Li, C.; Li, S.; Liu, T.; Feng, H.; Wang, J. Reduced white matter integrity in primary open-angle glaucoma: A DTI study using tract-based spatial statistics. J. Neuroradiol. 2013, 40, 89–93. [Google Scholar] [CrossRef]
- Gerlach, D.A.; Marshall-Goebel, K.; Hasan, K.M.; Kramer, L.A.; Alperin, N.; Rittweger, J. MRI-derived diffusion parameters in the human optic nerve and its surrounding sheath during head-down tilt. NPJ Microgravity 2017, 3, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, M.A.; Knott, M.; Heidemann, R.; Michelson, G.; Kober, T.; Dörfler, A.; Engelhorn, T. Investigation of lateral geniculate nucleus volume and diffusion tensor imaging in patients with normal tension glaucoma using 7 tesla magnetic resonance imaging. PLoS ONE 2018, 13, e0198830. [Google Scholar] [CrossRef]
- Michelson, G.; Engelhorn, T.; Wärntges, S.; El Rafei, A.; Hornegger, J.; Doerfler, A. DTI parameters of axonal integrity and demyelination of the optic radiation correlate with glaucoma indices. Graefe’s Arch. Clin. Exp. Ophthalmol. 2013, 251, 243–253. [Google Scholar] [CrossRef] [PubMed]
- Sidek, S.; Ramli, N.; Rahmat, K.; Abdulrahman, F.; Tan, L. Glaucoma severity affects diffusion tensor imaging (DTI) parameters of the optic nerve and optic radiation. Eur. J. Radiol. 2014, 83, 1437–1441. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.; Yin, D.; Hu, C.; Morelli, J.N.; Hu, S.; Yan, X.; Xu, D. Whole-brain voxel-based analysis of diffusion tensor MRI parameters in patients with primary open angle glaucoma and correlation with clinical glaucoma stage. Neuroradiology 2012, 55, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Sun, J.; Zhang, X.; Feng, Y.; Pan, A.; Gao, M.; Zhao, H. Microstructural visual pathway abnormalities in patients with primary glaucoma: 3 T diffusion kurtosis imaging study. Clin. Radiol. 2018, 73, 591.e9–591.e15. [Google Scholar] [CrossRef]
- Song, X.-Y.; Puyang, Z.; Chen, A.-H.; Zhao, J.; Li, X.-J.; Chen, Y.-Y.; Tang, W.-J.; Zhang, Y.-Y. Diffusion Tensor Imaging Detects Microstructural Differences of Visual Pathway in Patients with Primary Open-Angle Glaucoma and Ocular Hypertension. Front. Hum. Neurosci. 2018, 12, 426. [Google Scholar] [CrossRef] [Green Version]
- Stowell, C.; Burgoyne, C.; Tamm, E.R.; Ethier, C.R.; Dowling, J.E.; Downs, C.; Ellisman, M.H.; Fisher, S.; Fortune, B.; Fruttiger, M.; et al. Biomechanical aspects of axonal damage in glaucoma: A brief review. Exp. Eye Res. 2017, 157, 13–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, M.C.; Conner, I.P.; Teng, C.; Lawrence, J.D.; Safiullah, Z.; Wang, B.; Bilonick, R.A.; Kim, S.-G.; Wollstein, G.; Schuman, J.S.; et al. Retinal Structures and Visual Cortex Activity are Impaired Prior to Clinical Vision Loss in Glaucoma. Sci. Rep. 2016, 6, 31464. [Google Scholar] [CrossRef] [PubMed]
- Steven, A.J.; Zhuo, J.; Melhem, E.R. Diffusion Kurtosis Imaging: An Emerging Technique for Evaluating the Microstructural Environment of the Brain. Am. J. Roentgenol. 2014, 202, W26–W33. [Google Scholar] [CrossRef]
- Jensen, J.H.; Helpern, J.A.; Ramani, A.; Lu, H.; Kaczynski, K. Diffusional kurtosis imaging: The quantification of non-gaussian water diffusion by means of magnetic resonance imaging. Magn. Reson. Med. 2005, 53, 1432–1440. [Google Scholar] [CrossRef] [PubMed]
- Aksoy, D.Ö.; Akkan, J.C.U.; Alkan, A.; Aralaşmak, A.; Temur, H.O.; Yurtsever, I. Magnetic Resonance Spectroscopy Features of the Visual Pathways in Patients with Glaucoma. Clin. Neuroradiol. 2019, 29, 615–621. [Google Scholar] [CrossRef]
- Nucci, C.; Garaci, F.; Altobelli, S.; Di Ciò, F.; Martucci, A.; Aiello, F.; Lanzafame, S.; Di Giuliano, F.; Picchi, E.; Minosse, S.; et al. Diffusional Kurtosis Imaging of White Matter Degeneration in Glaucoma. J. Clin. Med. 2020, 9, 3122. [Google Scholar] [CrossRef]
- Maurano, S.T.P.; Da Silva, D.J.; Ávila, M.P.; Magacho, L. Cognitive evaluation of patients with glaucoma and its comparison with individuals with Alzheimer’s disease. Int. Ophthalmol. 2017, 38, 1839–1844. [Google Scholar] [CrossRef]
- Li, T.; Qu, X.; Chen, W.; Wang, Q.; Wang, H.; Wang, Y.; Huang, C.; Zhang, X.; Wang, N.; Xian, J. Altered information flow and microstructure abnormalities of visual cortex in normal-tension glaucoma: Evidence from resting-state fMRI and DKI. Brain Res. 2020, 1741, 146874. [Google Scholar] [CrossRef]
- De Santis, S.; Jones, D.K.; Roebroeck, A. Including diffusion time dependence in the extra-axonal space improves in vivo estimates of axonal diameter and density in human white matter. NeuroImage 2016, 130, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Benitez, A.; Jensen, J.H.; Falangola, M.F.; Nietert, P.J.; Helpern, J.A. Modeling white matter tract integrity in aging with diffusional kurtosis imaging. Neurobiol. Aging 2018, 70, 265–275. [Google Scholar] [CrossRef]
- Chung, S.; Fieremans, E.; Wang, X.; Kucukboyaci, N.E.; Morton, C.J.; Babb, J.; Amorapanth, P.; Foo, F.-Y.; Novikov, D.; Flanagan, S.; et al. White Matter Tract Integrity: An Indicator of Axonal Pathology after Mild Traumatic Brain Injury. J. Neurotrauma 2018, 35, 1015–1020. [Google Scholar] [CrossRef] [PubMed]
- Fieremans, E.; Benitez, A.; Jensen, J.; Falangola, M.; Tabesh, A.; Deardorff, R.; Spampinato, M.; Babb, J.; Novikov, D.; Ferris, S.; et al. Novel White Matter Tract Integrity Metrics Sensitive to Alzheimer Disease Progression. Am. J. Neuroradiol. 2013, 34, 2105–2112. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.K. Diffusion MRI Theory, Methods, and Applications; Oxford University Press: New York, NY, USA, 2010. [Google Scholar]
- You, Y.; Joseph, C.; Wang, C.; Gupta, V.; Liu, S.; Yiannikas, C.; Chua, B.E.; Chitranshi, N.; Shen, T.; Dheer, Y.; et al. Demyelination precedes axonal loss in the transneuronal spread of human neurodegenerative disease. Brain 2019, 142, 426–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanekamp, S.; Ćurčić-Blake, B.; Caron, B.; McPherson, B.; Timmer, A.; Prins, D.; Boucard, C.C.; Yoshida, M.; Ida, M.; Hunt, D.; et al. White matter alterations in glaucoma and monocular blindness differ outside the visual system. Sci. Rep. 2021, 11, 6866. [Google Scholar] [CrossRef] [PubMed]
- McDowell, C.M.; Kizhatil, K.; Elliott, M.H.; Overby, D.R.; van Batenburg-Sherwood, J.; Millar, J.C.; Kuehn, M.H.; Zode, G.; Acott, T.S.; Anderson, M.G.; et al. Consensus Recommendation for Mouse Models of Ocular Hypertension to Study Aqueous Humor Outflow and Its Mechanisms. Investig. Opthalmol. Vis. Sci. 2022, 63, 12. [Google Scholar] [CrossRef]
- Jones, D.K.; Knösche, T.R.; Turner, R. White matter integrity, fiber count, and other fallacies: The do’s and don’ts of diffusion MRI. NeuroImage 2013, 73, 239–254. [Google Scholar] [CrossRef]
- Ding, A.Y.; Chan, K.C.; Wu, E.X. Effect of cerebrovascular changes on brain DTI quantitation: A hypercapnia study. Magn. Reson. Imaging 2012, 30, 993–1001. [Google Scholar] [CrossRef]
- Jenkinson, M.; Bannister, P.; Brady, M.; Smith, S. Improved Optimization for the Robust and Accurate Linear Registration and Motion Correction of Brain Images. NeuroImage 2002, 17, 825–841. [Google Scholar] [CrossRef]
- Yamada, M.; Momoshima, S.; Masutani, Y.; Fujiyoshi, K.; Abe, O.; Nakamura, M.; Aoki, S.; Tamaoki, N.; Okano, H. Diffusion-tensor neuronal fiber tractography and manganese-enhanced mr imaging of primate visual pathway in the common marmoset: Preliminary results. Radiology 2008, 249, 855–864. [Google Scholar] [CrossRef]
- Pottabatula, B.; Smith, G.; Nagaraja, N.; Albayram, M.S. Demonstration of acute retinal ischemia on diffusion weighted magnetic resonance imaging. Clin. Imaging 2020, 59, 126–128. [Google Scholar] [CrossRef]
- Tuch, D.S. Q-ball imaging. Magn. Reson. Med. 2004, 52, 1358–1372. [Google Scholar] [CrossRef]
- Fujiyoshi, K.; Hikishima, K.; Nakahara, J.; Tsuji, O.; Hata, J.; Konomi, T.; Nagai, T.; Shibata, S.; Kaneko, S.; Iwanami, A.; et al. Application ofq-Space Diffusion MRI for the Visualization of White Matter. J. Neurosci. 2016, 36, 2796–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Miranda, J.C.; Pathak, S.; Engh, J.; Jarbo, K.; Verstynen, T.; Yeh, F.-C.; Wang, Y.; Mintz, A.; Boada, F.; Schneider, W.; et al. High-definition fiber tractography of the human brain. Neurosurgery 2012, 71, 430–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnett, A. Theory of Q-ball imaging redux: Implications for fiber tracking. Magn. Reson. Med. 2009, 62, 910–923. [Google Scholar] [CrossRef]
- Hui, E.S.; Cheung, M.M.; Chan, K.C.; Wu, E.X. B-value dependence of DTI quantitation and sensitivity in detecting neural tissue changes. NeuroImage 2010, 49, 2366–2374. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, V.; Bang, J.W.; Parra, C.; Colbert, M.K.; O’Connell, C.; Arshad, A.; Faiq, M.A.; Conner, I.P.; Redfern, M.S.; Wollstein, G.; et al. Widespread brain reorganization perturbs visuomotor coordination in early glaucoma. Sci. Rep. 2019, 9, 14168. [Google Scholar] [CrossRef]
- Lee, J.K.; Koppelmans, V.; Riascos, R.; Hasan, K.M.; Pasternak, O.; Mulavara, A.P.; Bloomberg, J.J.; Seidler, R. Spaceflight-Associated Brain White Matter Microstructural Changes and Intracranial Fluid Redistribution. JAMA Neurol. 2019, 76, 412–419. [Google Scholar] [CrossRef]
- Wang, J.-J.; Lin, W.-Y.; Lu, C.-S.; Weng, Y.-H.; Ng, S.-H.; Wang, C.-H.; Liu, H.-L.; Hsieh, R.-H.; Wan, Y.-L.; Wai, Y.-Y. Parkinson Disease: Diagnostic Utility of Diffusion Kurtosis Imaging. Radiology 2011, 261, 210–217. [Google Scholar] [CrossRef]
- Surova, Y.; Lampinen, B.; Nilsson, M.; Lätt, J.; Hall, S.; Widner, H.; van Westen, D.; Hansson, O.; Swedish BioFINDER study; van Westen, D.; et al. Alterations of Diffusion Kurtosis and Neurite Density Measures in Deep Grey Matter and White Matter in Parkinson’s Disease. PLoS ONE 2016, 11, e0157755. [Google Scholar] [CrossRef] [Green Version]
- Drouyer, E.; Dkhissi-Benyahya, O.; Chiquet, C.; WoldeMussie, E.; Ruiz, G.; Wheeler, L.A.; Denis, P.; Cooper, H.M. Glaucoma Alters the Circadian Timing System. PLoS ONE 2008, 3, e3931. [Google Scholar] [CrossRef] [Green Version]
- Bang, J.W.; Parra, C.; Wollstein, G.; Schuman, J.S.; Chan, K. Sleep-regulating systems are impaired in glaucoma. Investig. Ophthalmol. Vis. Sci. 2021, 62, 2347. [Google Scholar]
- Koller, K.; Rafal, R.D.; Mullins, P.G. Circadian circuits in humans: White matter microstructure predicts daytime sleepiness. Cortex 2019, 122, 97–107. [Google Scholar] [CrossRef] [PubMed]
- Narayana, P.A.; Yu, X.; Hasan, K.M.; Wilde, E.A.; Levin, H.S.; Hunter, J.V.; Miller, E.R.; Patel, V.K.S.; Robertson, C.S.; McCarthy, J.J. Multi-modal MRI of mild traumatic brain injury. NeuroImage Clin. 2015, 7, 87–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, L.; Nguyen, J.V.; Lehar, M.; Menon, A.; Rha, E.; Arena, J.; Ryu, J.; Marsh-Armstrong, N.; Marmarou, C.R.; Koliatsos, V.E. Repetitive mild traumatic brain injury with impact acceleration in the mouse: Multifocal axonopathy, neuroinflammation, and neurodegeneration in the visual system. Exp. Neurol. 2016, 275 Pt 3, 436–449. [Google Scholar] [CrossRef]
- Paul, K.; Huelnhagen, T.; Oberacker, E.; Wenz, D.; Kuehne, A.; Waiczies, H.; Schmitter, S.; Stachs, O.; Niendorf, T. Multiband diffusion-weighted MRI of the eye and orbit free of geometric distortions using a RARE-EPI hybrid. NMR Biomed. 2018, 31, e3872. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, M.; Lätt, J.; van Westen, D.; Brockstedt, S.; Lasič, S.; Ståhlberg, F.; Topgaard, D. Noninvasive mapping of water diffusional exchange in the human brain using filter-exchange imaging. Magn. Reson. Med. 2013, 69, 1572–1580. [Google Scholar] [CrossRef]
- Nilsson, M.; Lasič, S.; Drobnjak, I.; Topgaard, D.; Westin, C.-F. Resolution limit of cylinder diameter estimation by diffusion MRI: The impact of gradient waveform and orientation dispersion. NMR Biomed. 2017, 30, e3711. [Google Scholar] [CrossRef]
- Sims, J.R.; Chen, A.M.; Sun, Z.; Deng, W.; Colwell, N.A.; Colbert, M.K.; Zhu, J.; Sainulabdeen, A.; Faiq, M.A.; Bang, J.W. Role of Structural, Metabolic, and Functional MRI in Monitoring Visual System Impairment and Recovery. J. Magn. Reson. Imaging 2021, 54, 1706–1729. [Google Scholar] [CrossRef]
- Ho, L.C.; Sigal, I.A.; Jan, N.-J.; Yang, X.; Van Der Merwe, Y.; Yu, Y.; Chau, Y.; Leung, C.; Conner, I.P.; Jin, T.; et al. Non-invasive MRI Assessments of Tissue Microstructures and Macromolecules in the Eye upon Biomechanical or Biochemical Modulation. Sci. Rep. 2016, 6, 32080. [Google Scholar] [CrossRef]
- Zhang, P.; Wen, W.; Sun, X.; He, S. Selective reduction of fMRI responses to transient achromatic stimuli in the magnocellular layers of the LGN and the superficial layer of the SC of early glaucoma patients. Hum. Brain Mapp. 2016, 37, 558–569. [Google Scholar] [CrossRef]
- Sidek, S.; Ramli, N.; Rahmat, K.; Ramli, N.M.; Abdulrahman, F.; Kuo, T.L. In vivo proton magnetic resonance spectroscopy (1H-MRS) evaluation of the metabolite concentration of optic radiation in primary open angle glaucoma. Eur. Radiol. 2016, 26, 4404–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doganay, S.; Cankaya, C.; Alkan, A. Evaluation of corpus geniculatum laterale and vitreous fluid by magnetic resonance spectroscopy in patients with glaucoma; a preliminary study. Eye 2012, 26, 1044–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, K.C.; So, K.-F.; Wu, E.X. Proton magnetic resonance spectroscopy revealed choline reduction in the visual cortex in an experimental model of chronic glaucoma. Exp. Eye Res. 2009, 88, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Taoka, T.; Masutani, Y.; Kawai, H.; Nakane, T.; Matsuoka, K.; Yasuno, F.; Kishimoto, T.; Naganawa, S. Evaluation of glymphatic system activity with the diffusion MR technique: Diffusion tensor image analysis along the perivascular space (DTI-ALPS) in Alzheimer’s disease cases. Jpn. J. Radiol. 2017, 35, 172–178. [Google Scholar] [CrossRef]
- Deng, W.; Liu, C.; Parra, C.; Sims, J.R.; Faiq, M.A.; Sainulabdeen, A.; Song, H.; Chan, K.C. Quantitative imaging of the clearance systems in the eye and the brain. Quant. Imaging Med. Surg. 2020, 10, 1–14. [Google Scholar] [CrossRef]
- Ho, L.C.; Conner, I.P.; Do, C.-W.; Kim, S.-G.; Wu, E.X.; Wollstein, G.; Schuman, J.S.; Chan, K.C.W. In Vivo Assessment of Aqueous Humor Dynamics Upon Chronic Ocular Hypertension and Hypotensive Drug Treatment Using Gadolinium-Enhanced MRI. Investig. Opthalmol. Vis. Sci. 2014, 55, 3747–3757. [Google Scholar] [CrossRef] [Green Version]
- Chan, K.C.; Fu, Q.-L.; Hui, E.S.; So, K.-F.; Wu, E.X. Evaluation of the retina and optic nerve in a rat model of chronic glaucoma using in vivo manganese-enhanced magnetic resonance imaging. NeuroImage 2008, 40, 1166–1174. [Google Scholar] [CrossRef]
- Chan, K.C.; Fu, Q.-L.; Guo, H.; So, K.-F.; Wu, E.X. GD-DTPA enhanced MRI of ocular transport in a rat model of chronic glaucoma. Exp. Eye Res. 2008, 87, 334–341. [Google Scholar] [CrossRef]
- Wheeler, L.A.; Gil, D.W.; WoldeMussie, E. Role of Alpha-2 Adrenergic Receptors in Neuroprotection and Glaucoma. Surv. Ophthalmol. 2001, 45 (Suppl. 3), S290–S294. [Google Scholar] [CrossRef]
- Takahashi, N.; Matsunaga, N.; Natsume, T.; Kitazawa, C.; Itani, Y.; Hama, A.; Hayashi, I.; Shimazawa, M.; Hara, H.; Takamatsu, H. A longitudinal comparison in cynomolgus macaques of the effect of brimonidine on optic nerve neuropathy using diffusion tensor imaging magnetic resonance imaging and spectral domain optical coherence tomography. Heliyon 2021, 7, e06701. [Google Scholar] [CrossRef]
- Wiendl, H.; Elger, C.; Förstl, H.; Hartung, H.-P.; Oertel, W.; Reichmann, H.; Schwab, S. Gaps Between Aims and Achievements in Therapeutic Modification of Neuronal Damage (“Neuroprotection”). Neurotherapeutics 2015, 12, 449–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
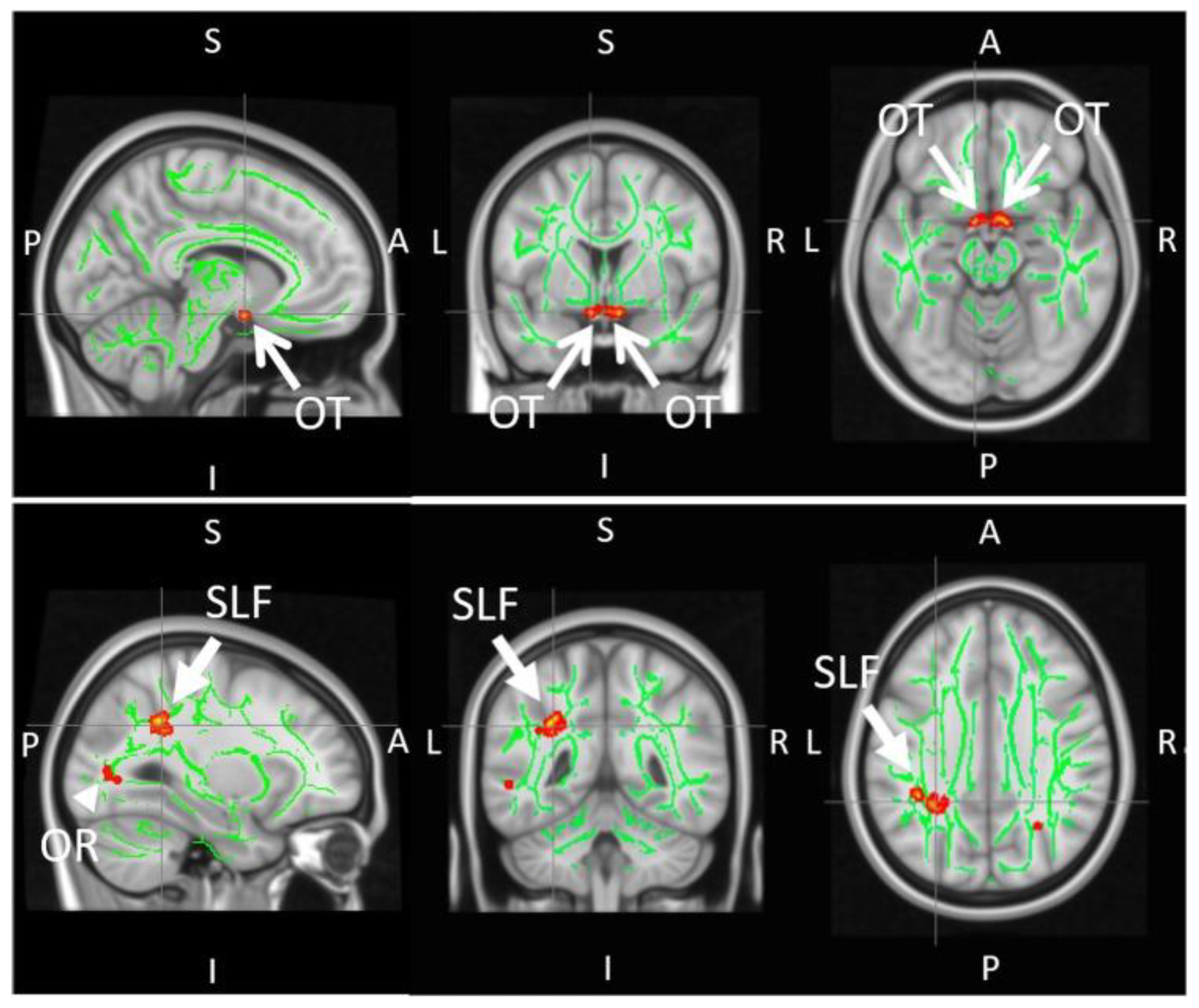{kind=link}
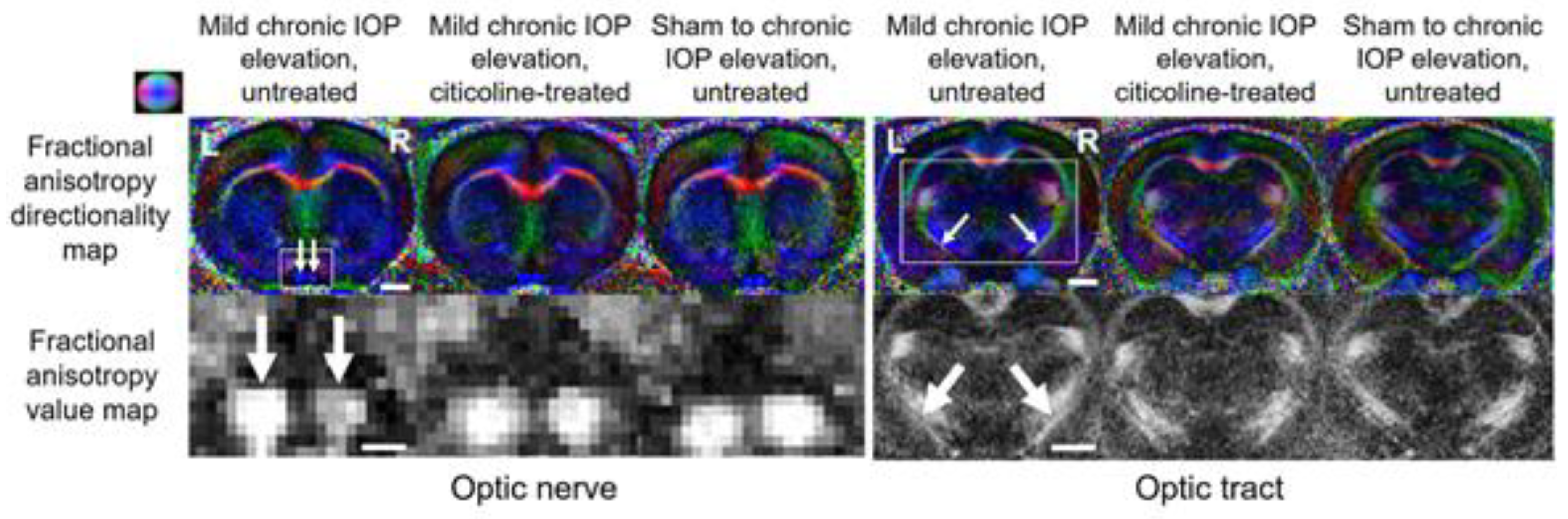{kind=link}
| DTI Parameter | Equation | |
|---|---|---|
| Mean Diffusivity (MD) | (λ1 + λ2 + λ3)/3 | (2) |
| Fractional Anisotropy (FA) | √(3[(λ1 − MD)2 + (λ2 − MD)2 + (λ3 − MD)2])/(2(λ12 + λ22 + λ32)] | (3) |
| Axial Diffusivity (AD) | λ1 | (4) |
| Radial Diffusivity (RD) | (λ2 + λ3)/2 | (5) |
| Glaucoma Type | DTI Parametric Change | Field Strength (T) | Reference | |||
|---|---|---|---|---|---|---|
| FA | AD | RD | MD | |||
| Normal-Tension Glaucoma | ↓ | ↑ | ↑ | ↑ | 1.5 | [31] |
| Normal-Tension Glaucoma | ↓ | ↑ | NA | NA | 1.5 | [77] |
| Normal-Tension Glaucoma | ↓ | NA | NA | NA | 7 | [87] |
| Normal-Tension Glaucoma | ↓ | ↓ | ↑ | ↑ | 3 | [81] |
| Normal-Tension Glaucoma | ↓ | NS | ↑ | NA | 3 | [88] |
| Primary Open-Angle Glaucoma | ↓ | NA | NA | ↑ | 3 | [89] |
| Primary Open-Angle Glaucoma | ↓ | NS | ↑ | NS | 3 | [90] |
| Primary Open-Angle Glaucoma | ↓ | NS | ↑ | NA | 3 | [88] |
| Primary Open-Angle Glaucoma | ↓ | ↑ | ↑ | ↑ | 3 | [83] |
| Primary Open-Angle Glaucoma | ↓ | NA | NA | NA | 3 | [85] |
| Primary Open-Angle Glaucoma | ↓ | NA | NA | ↑ | 3 | [91] |
| Primary Open-Angle Glaucoma | ↓ | NA | NA | ↑ | 3 | [92] |
| Optic Nerve | Optic Tract | Optic Radiation (LGN to V1) | LGN | Field Strength (T) | Glaucoma Type | Reference | |
|---|---|---|---|---|---|---|---|
| Fractional Anisotropy | ↓ | ↓ | ↓ | NA | 7 | NTG | [87] |
| Fractional Anisotropy | ↓ | NA | ↓ | NA | 3 | POAG | [72] |
| Fractional Anisotropy | NA | NS | ↓ | NS | 3 | POAG | [92] |
| Radial Diffusivity | ↑ | NA | ↑ | NA | 3 | POAG | [72] |
| Mean Diffusivity | NA | ↑ * | NS | ↑ | 3 | POAG | [92] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendoza, M.; Shotbolt, M.; Faiq, M.A.; Parra, C.; Chan, K.C. Advanced Diffusion MRI of the Visual System in Glaucoma: From Experimental Animal Models to Humans. Biology 2022, 11, 454. https://doi.org/10.3390/biology11030454
Mendoza M, Shotbolt M, Faiq MA, Parra C, Chan KC. Advanced Diffusion MRI of the Visual System in Glaucoma: From Experimental Animal Models to Humans. Biology. 2022; 11(3):454. https://doi.org/10.3390/biology11030454
Chicago/Turabian StyleMendoza, Monica, Max Shotbolt, Muneeb A. Faiq, Carlos Parra, and Kevin C. Chan. 2022. "Advanced Diffusion MRI of the Visual System in Glaucoma: From Experimental Animal Models to Humans" Biology 11, no. 3: 454. https://doi.org/10.3390/biology11030454
APA StyleMendoza, M., Shotbolt, M., Faiq, M. A., Parra, C., & Chan, K. C. (2022). Advanced Diffusion MRI of the Visual System in Glaucoma: From Experimental Animal Models to Humans. Biology, 11(3), 454. https://doi.org/10.3390/biology11030454