
Proteomic Analysis of Retinal Tissue in an S100B Autoimmune Glaucoma Model

 ,
,  , ,
, ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Immunization
2.3. Mass Spectrometric Analysis of Retinae Samples
2.4. Data Evaluation and Functional Analysis
2.5. Tissue Preparation for Immunohistology
2.6. Hematoxylin and Eosin Staining
2.7. Immunofluorescence Staining
2.8. Histological Evaluation
2.9. Statistics of Immunohistology Evaluations
3. Results
3.1. Characteristics of Proteins in the Retina-Specific Spectral Library
3.2. Comparative Proteomic Analysis Revealed Complex, Time-Dependent Regulations after S100B Immunization
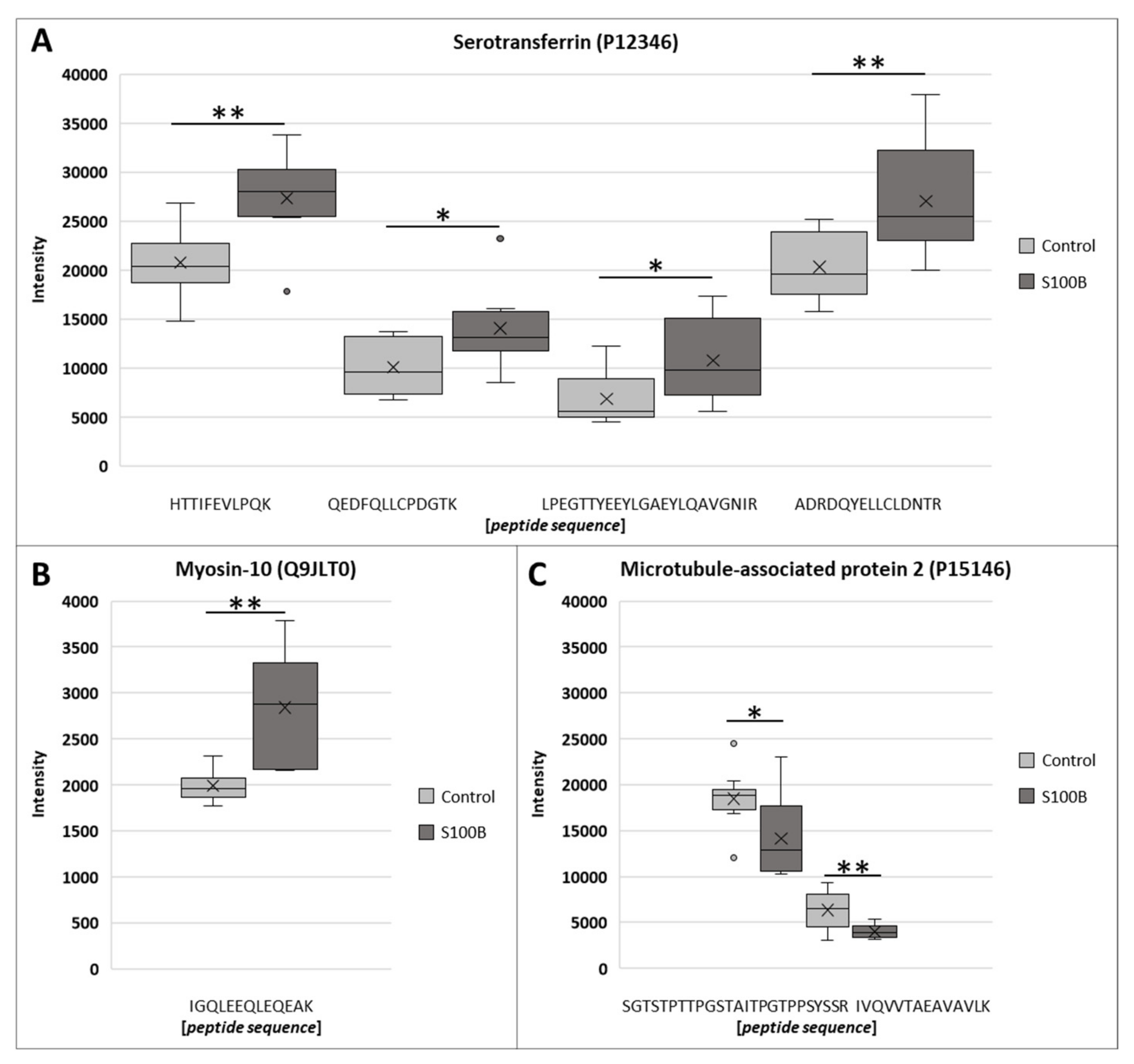
3.3. Characteristics of Differential Proteins
3.4. No Loss of Retinal Ganglion Cells
3.5. Altered Expression of Calretinin+ Amacrine Cells
3.6. Different HSP60 Expression after Immunization
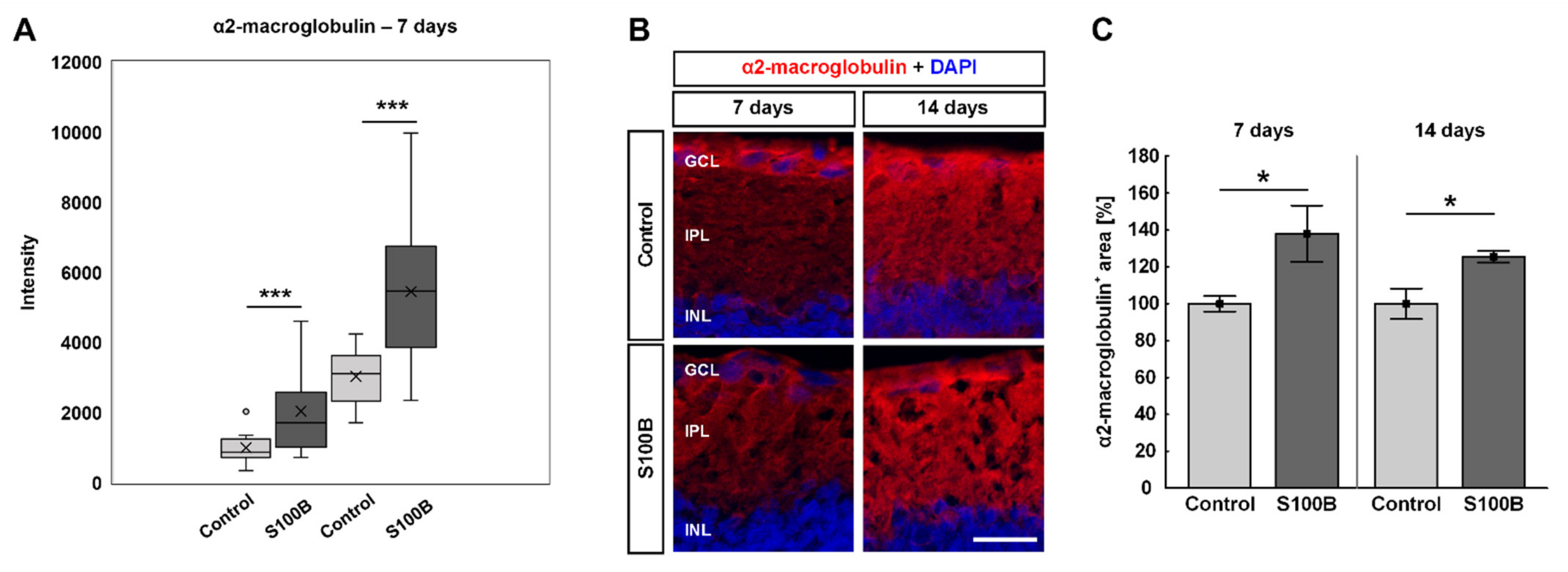
3.7. Increased Levels of α2-Macroglobulin in S100B Retinae
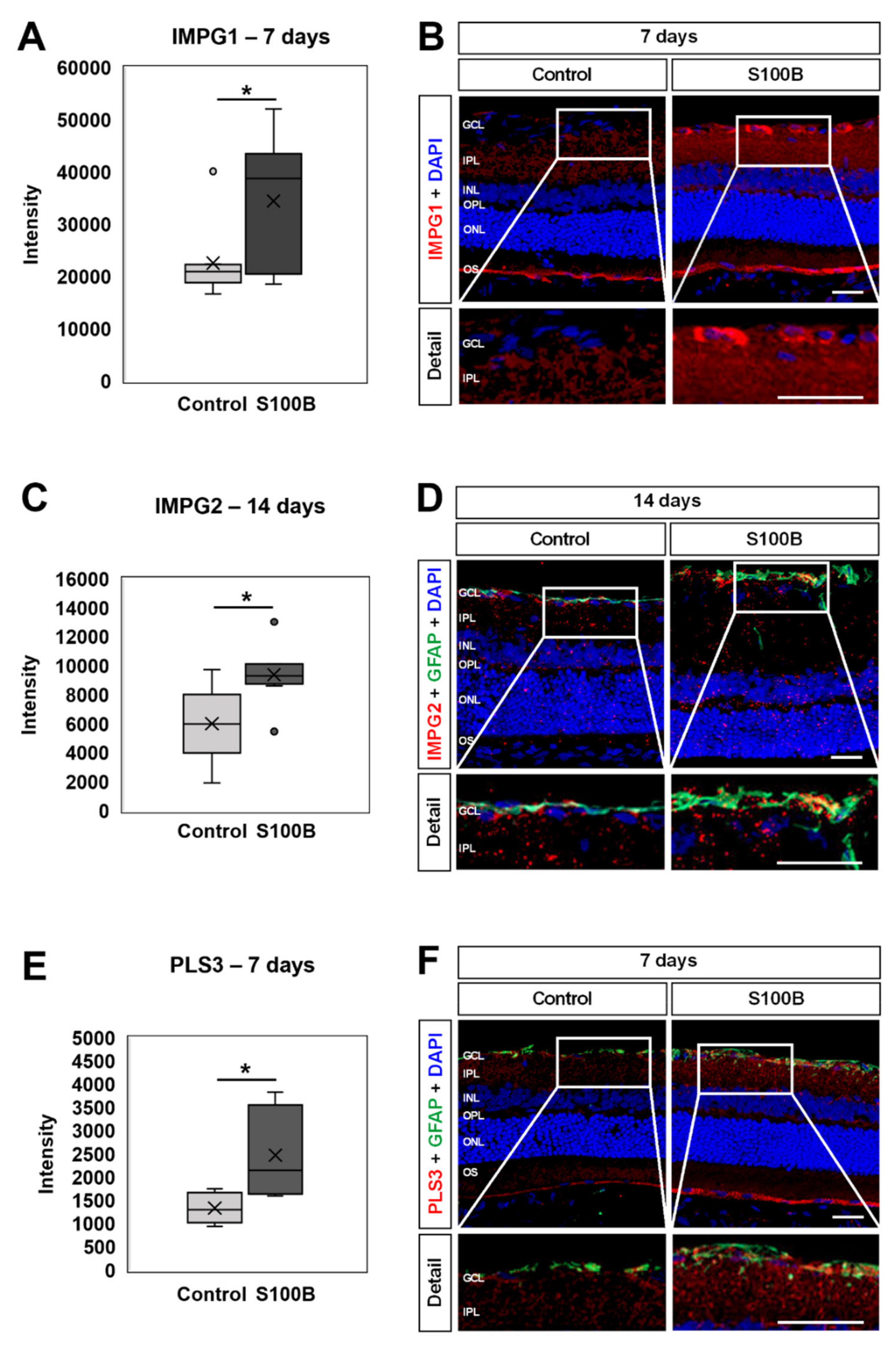
3.8. Up-Regulation of Interphotoreceptor Matrix Proteoglycans and Plastin 3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- EGS. European Glaucoma Society Terminology and Guidelines for Glaucoma, 4th Edition—Chapter 2: Classification and terminology. Br. J. Ophthalmol. 2017, 101, 73–127. [Google Scholar] [CrossRef] [Green Version]
- Hennis, A.; Wu, S.Y.; Nemesure, B.; Honkanen, R.; Leske, M.C.; Barbados Eye Studies, G. Awareness of incident open-angle glaucoma in a population study: The Barbados Eye Studies. Ophthalmology 2007, 114, 1816–1821. [Google Scholar] [CrossRef] [PubMed]
- Leite, M.T.; Sakata, L.M.; Medeiros, F.A. Managing glaucoma in developing countries. Arq. Bras. Oftalmol. 2011, 74, 83–84. [Google Scholar] [CrossRef] [Green Version]
- Rotchford, A.P.; Kirwan, J.F.; Muller, M.A.; Johnson, G.J.; Roux, P. Temba glaucoma study: A population-based cross-sectional survey in urban South Africa. Ophthalmology 2003, 110, 376–382. [Google Scholar] [CrossRef]
- Sathyamangalam, R.V.; Paul, P.G.; George, R.; Baskaran, M.; Hemamalini, A.; Madan, R.V.; Augustian, J.; Prema, R.; Lingam, V. Determinants of glaucoma awareness and knowledge in urban Chennai. Indian J. Ophthalmol. 2009, 57, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Budenz, D.L.; Barton, K.; Whiteside-de Vos, J.; Schiffman, J.; Bandi, J.; Nolan, W.; Herndon, L.; Kim, H.; Hay-Smith, G.; Tielsch, J.M.; et al. Prevalence of glaucoma in an urban West African population: The Tema Eye Survey. JAMA Ophthalmol. 2013, 131, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Mallick, J.; Devi, L.; Malik, P.K.; Mallick, J. Update on Normal Tension Glaucoma. J. Ophthalmic Vis. Res. 2016, 11, 204–208. [Google Scholar] [CrossRef]
- Sommer, A.; Tielsch, J.M.; Katz, J.; Quigley, H.A.; Gottsch, J.D.; Javitt, J.; Singh, K. Relationship between intraocular pressure and primary open angle glaucoma among white and black Americans. The Baltimore Eye Survey. Arch. Ophthalmol. 1991, 109, 1090–1095. [Google Scholar] [CrossRef]
- Vasudevan, S.K.; Gupta, V.; Crowston, J.G. Neuroprotection in glaucoma. Indian J. Ophthalmol. 2011, 59 (Suppl. S1), S102–S113. [Google Scholar] [CrossRef]
- Sucher, N.J.; Lipton, S.A.; Dreyer, E.B. Molecular basis of glutamate toxicity in retinal ganglion cells. Vis. Res. 1997, 37, 3483–3493. [Google Scholar] [CrossRef] [Green Version]
- Tezel, G.; Wax, M.B. The immune system and glaucoma. Curr. Opin. Ophthalmol. 2004, 15, 80–84. [Google Scholar] [CrossRef] [PubMed]
- Grus, F.H.; Joachim, S.C.; Wuenschig, D.; Rieck, J.; Pfeiffer, N. Autoimmunity and glaucoma. J. Glaucoma 2008, 17, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wax, M.B. The case for autoimmunity in glaucoma. Exp. Eye Res. 2011, 93, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Joachim, S.C.; Pfeiffer, N.; Grus, F.H. Autoantibodies in patients with glaucoma: A comparison of IgG serum antibodies against retinal, optic nerve, and optic nerve head antigens. Graefes Arch. Clin. Exp. Ophthalmol. 2005, 243, 817–823. [Google Scholar] [CrossRef]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Analysis of IgG antibody patterns against retinal antigens and antibodies to alpha-crystallin, GFAP, and alpha-enolase in sera of patients with “wet” age-related macular degeneration. Graefes Arch. Clin. Exp. Ophthalmol. 2007, 245, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Boehm, N.; Wolters, D.; Thiel, U.; Lossbrand, U.; Wiegel, N.; Pfeiffer, N.; Grus, F.H. New insights into autoantibody profiles from immune privileged sites in the eye: A glaucoma study. Brain Behav. Immun. 2012, 26, 96–102. [Google Scholar] [CrossRef]
- Gramlich, O.W.; Beck, S.; von Thun Und Hohenstein-Blaul, N.; Boehm, N.; Ziegler, A.; Vetter, J.M.; Pfeiffer, N.; Grus, F.H. Enhanced insight into the autoimmune component of glaucoma: IgG autoantibody accumulation and pro-inflammatory conditions in human glaucomatous retina. PLoS ONE 2013, 8, e57557. [Google Scholar] [CrossRef]
- Chang, E.E.; Goldberg, J.L. Glaucoma 2.0: Neuroprotection, neuroregeneration, neuroenhancement. Ophthalmology 2012, 119, 979–986. [Google Scholar] [CrossRef] [Green Version]
- Pascale, A.; Drago, F.; Govoni, S. Protecting the retinal neurons from glaucoma: Lowering ocular pressure is not enough. Pharmacol. Res. 2012, 66, 19–32. [Google Scholar] [CrossRef]
- Gupta, D.; Chen, P.P. Glaucoma. Am. Fam. Phys. 2016, 93, 668–674. [Google Scholar]
- Grus, F.H.; Boehm, N.; Beck, S.; Schlich, M.; Lossbrandt, U.; Pfeiffer, N. Autoantibody Profiles in Tear Fluid as a Diagnostic Tool in Glaucoma. Investig. Ophthalmol. Vis. Sci. 2010, 51, 6110. [Google Scholar]
- Michetti, F.; D’Ambrosi, N.; Toesca, A.; Puglisi, M.A.; Serrano, A.; Marchese, E.; Corvino, V.; Geloso, M.C. The S100B story: From biomarker to active factor in neural injury. J. Neurochem. 2019, 148, 168–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorci, G.; Riuzzi, F.; Arcuri, C.; Tubaro, C.; Bianchi, R.; Giambanco, I.; Donato, R. S100B protein in tissue development, repair and regeneration. World J. Biol. Chem. 2013, 4, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Heizmann, C.W.; Fritz, G.; Schafer, B.W. S100 proteins: Structure, functions and pathology. Front. Biosci. A J. Virtual Libr. 2002, 7, d1356–d1368. [Google Scholar]
- Bianchi, R.; Kastrisianaki, E.; Giambanco, I.; Donato, R. S100B protein stimulates microglia migration via RAGE-dependent up-regulation of chemokine expression and release. J. Biol. Chem. 2011, 286, 7214–7226. [Google Scholar] [CrossRef] [Green Version]
- Langeh, U.; Singh, S. Targeting S100B Protein as a Surrogate Biomarker and its Role in Various Neurological Disorders. Curr. Neuropharmacol. 2021, 19, 265–277. [Google Scholar] [CrossRef] [PubMed]
- Noristani, R.; Kuehn, S.; Stute, G.; Reinehr, S.; Stellbogen, M.; Dick, H.B.; Joachim, S.C. Retinal and Optic Nerve Damage is Associated with Early Glial Responses in an Experimental Autoimmune Glaucoma Model. J. Mol. Neurosci. 2016, 58, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Casola, C.; Reinehr, S.; Kuehn, S.; Stute, G.; Spiess, B.M.; Dick, H.B.; Joachim, S.C. Specific Inner Retinal Layer Cell Damage in an Autoimmune Glaucoma Model Is Induced by GDNF With or Without HSP27. Investig. Ophthalmol. Vis. Sci. 2016, 57, 3626–3639. [Google Scholar] [CrossRef] [Green Version]
- Casola, C.; Schiwek, J.E.; Reinehr, S.; Kuehn, S.; Grus, F.H.; Kramer, M.; Dick, H.B.; Joachim, S.C. S100 Alone Has the Same Destructive Effect on Retinal Ganglion Cells as in Combination with HSP 27 in an Autoimmune Glaucoma Model. J. Mol. Neurosci. 2015, 56, 228–236. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, S.; Guntermann, A.; Kuehn, S.; Palmhof, M.; Grotegut, P.; Serschnitzki, B.; Steinbach, S.; Dick, H.B.; Marcus, K.; Joachim, S.C.; et al. Rat retinae data for use as spectral library, for pathway remodeling as well as protein mapping. Data Brief 2021, 37, 107212. [Google Scholar] [CrossRef]
- Steinbach, S.; Serschnitzki, B.; Gerlach, M.; Marcus, K.; May, C. Spiked human. Data Brief 2019, 23, 103711. [Google Scholar] [CrossRef] [PubMed]
- Barkovits, K.; Pacharra, S.; Pfeiffer, K.; Steinbach, S.; Eisenacher, M.; Marcus, K.; Uszkoreit, J. Reproducibility, Specificity and Accuracy of Relative Quantification Using Spectral Library-based Data-independent Acquisition. Mol. Cell. Proteom. 2020, 19, 181–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jassal, B.; Matthews, L.; Viteri, G.; Gong, C.; Lorente, P.; Fabregat, A.; Sidiropoulos, K.; Cook, J.; Gillespie, M.; Haw, R.; et al. The reactome pathway knowledgebase. Nucleic Acids Res. 2020, 48, D498–D503. [Google Scholar] [CrossRef]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef]
- Benning, L.; Reinehr, S.; Grotegut, P.; Kuehn, S.; Stute, G.; Dick, H.B.; Joachim, S.C. Synapse and Receptor Alterations in Two Different S100B-Induced Glaucoma-Like Models. Int. J. Mol. Sci. 2020, 21, 6998. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, S.; Kuehn, S.; Casola, C.; Koch, D.; Stute, G.; Grotegut, P.; Dick, H.B.; Joachim, S.C. HSP27 immunization reinforces AII amacrine cell and synapse damage induced by S100 in an autoimmune glaucoma model. Cell Tissue Res. 2018, 371, 237–249. [Google Scholar] [CrossRef]
- Casson, R.J.; Chidlow, G.; Crowston, J.G.; Williams, P.A.; Wood, J.P.M. Retinal energy metabolism in health and glaucoma. Prog. Retin. Eye Res. 2021, 81, 100881. [Google Scholar] [CrossRef] [PubMed]
- Doozandeh, A.; Yazdani, S. Neuroprotection in Glaucoma. J. Ophthalmic Vis. Res. 2016, 11, 209–220. [Google Scholar] [CrossRef] [Green Version]
- Christensen, I.; Lu, B.; Yang, N.; Huang, K.; Wang, P.; Tian, N. The Susceptibility of Retinal Ganglion Cells to Glutamatergic Excitotoxicity Is Type-Specific. Front. Neurosci. 2019, 13, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinehr, S.; Reinhard, J.; Gandej, M.; Gottschalk, I.; Stute, G.; Faissner, A.; Dick, H.B.; Joachim, S.C. S100B immunization triggers NFκB and complement activation in an autoimmune glaucoma model. Sci. Rep. 2018, 8, 9821. [Google Scholar] [CrossRef] [PubMed]
- Reinehr, S.; Reinhard, J.; Wiemann, S.; Stute, G.; Kuehn, S.; Woestmann, J.; Dick, H.B.; Faissner, A.; Joachim, S.C. Early remodelling of the extracellular matrix proteins tenascin-C and phosphacan in retina and optic nerve of an experimental autoimmune glaucoma model. J. Cell. Mol. Med. 2016, 20, 2122–2137. [Google Scholar] [CrossRef] [PubMed]
- Quigley, H.A. Open-angle glaucoma. N. Engl. J. Med. 1993, 328, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.V.; Jope, R.S. The role of microtubule-associated protein 2 (MAP-2) in neuronal growth, plasticity, and degeneration. J. Neurosci. Res. 1992, 33, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Okabe, S.; Shiomura, Y.; Hirokawa, N. Immunocytochemical localization of microtubule-associated proteins 1A and 2 in the rat retina. Brain Res. 1989, 483, 335–346. [Google Scholar] [CrossRef]
- Gabriel, R.; Wilhelm, M.; Straznicky, C. Microtubule-associated protein 2 (MAP2)-immunoreactive neurons in the retina of Bufo marinus: Colocalisation with tyrosine hydroxylase and serotonin in amacrine cells. Cell Tissue Res. 1992, 269, 175–182. [Google Scholar] [CrossRef]
- Tucker, R.P.; Matus, A.I. Microtubule-associated proteins characteristic of embryonic brain are found in the adult mammalian retina. Dev. Biol. 1988, 130, 423–434. [Google Scholar] [CrossRef]
- Vaillant, A.R.; Zanassi, P.; Walsh, G.S.; Aumont, A.; Alonso, A.; Miller, F.D. Signaling mechanisms underlying reversible, activity-dependent dendrite formation. Neuron 2002, 34, 985–998. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Walker, G.; Lynn, S.A.; Keeling, E.; Munday, R.; Johnston, D.A.; Page, A.; Scott, J.A.; Goverdhan, S.; Lotery, A.J.; Ratnayaka, J.A. The Alzheimer’s-related amyloid beta peptide is internalised by R28 neuroretinal cells and disrupts the microtubule associated protein 2 (MAP-2). Exp. Eye Res. 2016, 153, 110–121. [Google Scholar] [CrossRef] [Green Version]
- Pow, D.V.; Sullivan, R.K. Nuclear kinesis, neurite sprouting and abnormal axonal projections of cone photoreceptors in the aged and AMD-afflicted human retina. Exp. Eye Res. 2007, 84, 850–857. [Google Scholar] [CrossRef] [PubMed]
- Kolb, H. Amacrine cells of the mammalian retina: Neurocircuitry and functional roles. Eye 1997, 11 Pt 6, 904–923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masland, R.H. The neuronal organization of the retina. Neuron 2012, 76, 266–280. [Google Scholar] [CrossRef] [Green Version]
- Wassle, H.; Boycott, B.B. Functional architecture of the mammalian retina. Physiol. Rev. 1991, 71, 447–480. [Google Scholar] [CrossRef]
- Akopian, A.; Kumar, S.; Ramakrishnan, H.; Viswanathan, S.; Bloomfield, S.A. Amacrine cells coupled to ganglion cells via gap junctions are highly vulnerable in glaucomatous mouse retinas. J. Comp. Neurol. 2016, 527, 159–173. [Google Scholar] [CrossRef]
- Pasteels, B.; Rogers, J.; Blachier, F.; Pochet, R. Calbindin and calretinin localization in retina from different species. Vis. Neurosci. 1990, 5, 1–16. [Google Scholar] [CrossRef]
- Gabriel, R.; Witkovsky, P. Cholinergic, but not the rod pathway-related glycinergic (All), amacrine cells contain calretinin in the rat retina. Neurosci. Lett. 1998, 247, 179–182. [Google Scholar] [CrossRef]
- Balasubramanian, R.; Gan, L. Development of Retinal Amacrine Cells and Their Dendritic Stratification. Curr. Ophthalmol. Rep. 2014, 2, 100–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boncristiano, S.; Calhoun, M.E.; Howard, V.; Bondolfi, L.; Kaeser, S.A.; Wiederhold, K.H.; Staufenbiel, M.; Jucker, M. Neocortical synaptic bouton number is maintained despite robust amyloid deposition in APP23 transgenic mice. Neurobiol. Aging 2005, 26, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Jansen, D.; Janssen, C.I.; Vanmierlo, T.; Dederen, P.J.; van Rooij, D.; Zinnhardt, B.; Nobelen, C.L.; Janssen, A.L.; Hafkemeijer, A.; Mutsaers, M.P.; et al. Cholesterol and synaptic compensatory mechanisms in Alzheimer’s disease mice brain during aging. J. Alzheimer’s Dis. JAD 2012, 31, 813–826. [Google Scholar] [CrossRef]
- Scheff, S.W.; Price, D.A. Synaptic pathology in Alzheimer’s disease: A review of ultrastructural studies. Neurobiol. Aging 2003, 24, 1029–1046. [Google Scholar] [CrossRef]
- Choii, G.; Ko, J. Gephyrin: A central GABAergic synapse organizer. Exp. Mol. Med. 2015, 47, e158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagarajan, S.K.; Fritschy, J.M. Gephyrin: A master regulator of neuronal function? Nat. Rev. Neurosci. 2014, 15, 141–156. [Google Scholar] [CrossRef] [Green Version]
- Hales, C.M.; Rees, H.; Seyfried, N.T.; Dammer, E.B.; Duong, D.M.; Gearing, M.; Montine, T.J.; Troncoso, J.C.; Thambisetty, M.; Levey, A.I.; et al. Abnormal gephyrin immunoreactivity associated with Alzheimer disease pathologic changes. J. Neuropathol. Exp. Neurol. 2013, 72, 1009–1015. [Google Scholar] [CrossRef]
- Tsai, T.; Grotegut, P.; Reinehr, S.; Joachim, S.C. Role of Heat Shock Proteins in Glaucoma. Int. J. Mol. Sci. 2019, 20, 5160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binder, R.J. Functions of heat shock proteins in pathways of the innate and adaptive immune system. J. Immunol. 2014, 193, 5765–5771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Srivastava, P. Roles of heat-shock proteins in innate and adaptive immunity. Nat. Rev. Immunol. 2002, 2, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Wax, M.B.; Tezel, G.; Saito, I.; Gupta, R.S.; Harley, J.B.; Li, Z.; Romano, C. Anti-Ro/SS-A positivity and heat shock protein antibodies in patients with normal-pressure glaucoma. Am. J. Ophthalmol. 1998, 125, 145–157. [Google Scholar] [CrossRef]
- Tezel, G.; Hernandez, R.; Wax, M.B. Immunostaining of heat shock proteins in the retina and optic nerve head of normal and glaucomatous eyes. Arch. Ophthalmol. 2000, 118, 511–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saccà, S.C.; Centofanti, M.; Izzotti, A. New Proteins as Vascular Biomarkers in Primary Open Angle Glaucomatous Aqueous Humor. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4242–4253. [Google Scholar] [CrossRef]
- Guo, C.; Wu, N.; Niu, X.; Wu, Y.; Chen, D.; Guo, W. Comparison of T Helper Cell Patterns in Primary Open-Angle Glaucoma and Normal-Pressure Glaucoma. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2018, 24, 1988–1996. [Google Scholar] [CrossRef] [PubMed]
- Grabska-Liberek, I.; Skonieczna, K.; Olesinska, M.; Terelak-Borys, B.; Kociecki, J.; Sikora, M.; Jamrozy-Witkowska, A.; Tesla, P.; Czarnocka, B. Levels of antibodies against human heat shock protein (HSP) 60 in patients with glaucoma in Poland. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2015, 21, 828–832. [Google Scholar] [CrossRef] [Green Version]
- Wax, M.B.; Tezel, G.; Yang, J.; Peng, G.; Patil, R.V.; Agarwal, N.; Sappington, R.M.; Calkins, D.J. Induced autoimmunity to heat shock proteins elicits glaucomatous loss of retinal ganglion cell neurons via activated T-cell-derived fas-ligand. J. Neurosci. 2008, 28, 12085–12096. [Google Scholar] [CrossRef] [Green Version]
- Grotegut, P.; Perumal, N.; Kuehn, S.; Smit, A.; Dick, H.B.; Grus, F.H.; Joachim, S.C. Minocycline reduces inflammatory response and cell death in a S100B retina degeneration model. J. Neuroinflamm. 2020, 17, 375. [Google Scholar] [CrossRef] [PubMed]
- Guan, S.S.; Sheu, M.L.; Yang, R.S.; Chan, D.C.; Wu, C.T.; Yang, T.H.; Chiang, C.K.; Liu, S.H. The pathological role of advanced glycation end products-downregulated heat shock protein 60 in islet beta-cell hypertrophy and dysfunction. Oncotarget 2016, 7, 23072–23087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bie, A.S.; Palmfeldt, J.; Hansen, J.; Christensen, R.; Gregersen, N.; Corydon, T.J.; Bross, P. A cell model to study different degrees of Hsp60 deficiency in HEK293 cells. Cell Stress Chaperones 2011, 16, 633–640. [Google Scholar] [CrossRef] [Green Version]
- Magnoni, R.; Palmfeldt, J.; Christensen, J.H.; Sand, M.; Maltecca, F.; Corydon, T.J.; West, M.; Casari, G.; Bross, P. Late onset motoneuron disorder caused by mitochondrial Hsp60 chaperone deficiency in mice. Neurobiol. Dis. 2013, 54, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Gonias, S.L.; Carmichael, A.; Mettenburg, J.M.; Roadcap, D.W.; Irvin, W.P.; Webb, D.J. Identical or overlapping sequences in the primary structure of human alpha(2)-macroglobulin are responsible for the binding of nerve growth factor-beta, platelet-derived growth factor-BB, and transforming growth factor-beta. J. Biol. Chem. 2000, 275, 5826–5831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LaMarre, J.; Wollenberg, G.K.; Gonias, S.L.; Hayes, M.A. Cytokine binding and clearance properties of proteinase-activated alpha 2-macroglobulins. Lab. Investig. A J. Tech. Methods Pathol. 1991, 65, 3–14. [Google Scholar]
- Barcelona, P.F.; Saragovi, H.U. A Pro-Nerve Growth Factor (proNGF) and NGF Binding Protein, alpha2-Macroglobulin, Differentially Regulates p75 and TrkA Receptors and Is Relevant to Neurodegeneration Ex Vivo and In Vivo. Mol. Cell. Biol. 2015, 35, 3396–3408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skornicka, E.L.; Shi, X.; Koo, P.H. Comparative binding of biotinylated neurotrophins to alpha(2)-macroglobulin family of proteins: Relationship between cytokine-binding and neuro-modulatory activities of the macroglobulins. J. Neurosci. Res. 2002, 67, 346–353. [Google Scholar] [CrossRef]
- Westwood, M.; Aplin, J.D.; Collinge, I.A.; Gill, A.; White, A.; Gibson, J.M. alpha 2-Macroglobulin: A new component in the insulin-like growth factor/insulin-like growth factor binding protein-1 axis. J. Biol. Chem. 2001, 276, 41668–41674. [Google Scholar] [CrossRef] [Green Version]
- Wolf, B.B.; Gonias, S.L. Neurotrophin binding to human alpha 2-macroglobulin under apparent equilibrium conditions. Biochemistry 1994, 33, 11270–11277. [Google Scholar] [CrossRef]
- French, K.; Yerbury, J.J.; Wilson, M.R. Protease activation of alpha2-macroglobulin modulates a chaperone-like action with broad specificity. Biochemistry 2008, 47, 1176–1185. [Google Scholar] [CrossRef] [PubMed]
- Whiten, D.R.; Cox, D.; Horrocks, M.H.; Taylor, C.G.; De, S.; Flagmeier, P.; Tosatto, L.; Kumita, J.R.; Ecroyd, H.; Dobson, C.M.; et al. Single-Molecule Characterization of the Interactions between Extracellular Chaperones and Toxic alpha-Synuclein Oligomers. Cell Rep. 2018, 23, 3492–3500. [Google Scholar] [CrossRef]
- Wyatt, A.R.; Zammit, N.W.; Wilson, M.R. Acute phase proteins are major clients for the chaperone action of alpha(2)-macroglobulin in human plasma. Cell Stress Chaperones 2013, 18, 161–170. [Google Scholar] [CrossRef] [Green Version]
- Yerbury, J.J.; Kumita, J.R.; Meehan, S.; Dobson, C.M.; Wilson, M.R. alpha2-Macroglobulin and haptoglobin suppress amyloid formation by interacting with prefibrillar protein species. J. Biol. Chem. 2009, 284, 4246–4254. [Google Scholar] [CrossRef] [Green Version]
- Wollenberg, G.K.; LaMarre, J.; Rosendal, S.; Gonias, S.L.; Hayes, M.A. Binding of tumor necrosis factor alpha to activated forms of human plasma alpha 2 macroglobulin. Am. J. Pathol. 1991, 138, 265–272. [Google Scholar]
- Borth, W.; Urbanski, A.; Prohaska, R.; Susanj, M.; Luger, T.A. Binding of recombinant interleukin-1 beta to the third complement component and alpha 2-macroglobulin after activation of serum by immune complexes. Blood 1990, 75, 2388–2395. [Google Scholar] [CrossRef] [Green Version]
- Matsuda, T.; Hirano, T.; Nagasawa, S.; Kishimoto, T. Identification of alpha 2-macroglobulin as a carrier protein for IL-6. J. Immunol. 1989, 142, 148–152. [Google Scholar]
- Bai, Y.; Sivori, D.; Woo, S.B.; Neet, K.E.; Lerner, S.F.; Saragovi, H.U. During glaucoma, alpha2-macroglobulin accumulates in aqueous humor and binds to nerve growth factor, neutralizing neuroprotection. Investig. Opthalmol. Vis. Sci. 2011, 52, 5260–5265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Z.; Rudzinski, M.; Meerovitch, K.; Lebrun-Julien, F.; Birman, E.; Di Polo, A.; Saragovi, H.U. Alpha2-macroglobulin is a mediator of retinal ganglion cell death in glaucoma. J. Biol. Chem. 2008, 283, 29156–29165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Shi, Z.; Zhuo, Y.; Liu, J.; Malakhov, A.; Ko, E.; Burgess, K.; Schaefer, H.; Esteban, P.F.; Tessarollo, L.; et al. In glaucoma the upregulated truncated TrkC.T1 receptor isoform in glia causes increased TNF-alpha production, leading to retinal ganglion cell death. Investig. Opthalmology Vis. Sci. 2010, 51, 6639–6651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Dergham, P.; Nedev, H.; Xu, J.; Galan, A.; Rivera, J.C.; ZhiHua, S.; Mehta, H.M.; Woo, S.B.; Sarunic, M.V.; et al. Chronic and acute models of retinal neurodegeneration TrkA activity are neuroprotective whereas p75NTR activity is neurotoxic through a paracrine mechanism. J. Biol. Chem. 2010, 285, 39392–39400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cornelissen, A.; Guo, L.; Sakamoto, A.; Virmani, R.; Finn, A.V. New insights into the role of iron in inflammation and atherosclerosis. EBioMedicine 2019, 47, 598–606. [Google Scholar] [CrossRef] [Green Version]
- Castillo, E.; Leon, J.; Mazzei, G.; Abolhassani, N.; Haruyama, N.; Saito, T.; Saido, T.; Hokama, M.; Iwaki, T.; Ohara, T.; et al. Author Correction: Comparative profiling of cortical gene expression in Alzheimer’s disease patients and mouse models demonstrates a link between amyloidosis and neuroinflammation. Sci. Rep. 2021, 11, 18377. [Google Scholar] [CrossRef]
- Funke, S.; Perumal, N.; Beck, S.; Gabel-Scheurich, S.; Schmelter, C.; Teister, J.; Gerbig, C.; Gramlich, O.W.; Pfeiffer, N.; Grus, F.H. Glaucoma related Proteomic Alterations in Human Retina Samples. Sci. Rep. 2016, 6, 29759. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Iglesias, H.; Alvarez, L.; Garcia, M.; Escribano, J.; Rodriguez-Calvo, P.P.; Fernandez-Vega, L.; Coca-Prados, M. Comparative proteomic study in serum of patients with primary open-angle glaucoma and pseudoexfoliation glaucoma. J. Proteom. 2014, 98, 65–78. [Google Scholar] [CrossRef]
- Ishikawa, M.; Sawada, Y.; Yoshitomi, T. Structure and function of the interphotoreceptor matrix surrounding retinal photoreceptor cells. Exp. Eye Res. 2015, 133, 3–18. [Google Scholar] [CrossRef] [Green Version]
- Felemban, M.; Dorgau, B.; Hunt, N.C.; Hallam, D.; Zerti, D.; Bauer, R.; Ding, Y.; Collin, J.; Steel, D.; Krasnogor, N.; et al. Extracellular matrix component expression in human pluripotent stem cell-derived retinal organoids recapitulates retinogenesis in vivo and reveals an important role for IMPG1 and CD44 in the development of photoreceptors and interphotoreceptor matrix. Acta Biomater. 2018, 74, 207–221. [Google Scholar] [CrossRef]
- Kuehn, M.H.; Hageman, G.S. Expression and characterization of the IPM 150 gene (IMPG1) product, a novel human photoreceptor cell-associated chondroitin-sulfate proteoglycan. Matrix Biol. J. Int. Soc. Matrix Biol. 1999, 18, 509–518. [Google Scholar] [CrossRef]
- Acharya, S.; Rodriguez, I.R.; Moreira, E.F.; Midura, R.J.; Misono, K.; Todres, E.; Hollyfield, J.G. SPACR, a novel interphotoreceptor matrix glycoprotein in human retina that interacts with hyaluronan. J. Biol. Chem. 1998, 273, 31599–31606. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Lee, J.W.; Nishiyama, K.; Shadrach, K.G.; Rayborn, M.E.; Hollyfield, J.G. SPACRCAN in the interphotoreceptor matrix of the mouse retina: Molecular, developmental and promoter analysis. Exp. Eye Res. 2003, 76, 1–14. [Google Scholar] [CrossRef]
- Reinhard, J.; Renner, M.; Wiemann, S.; Shakoor, D.A.; Stute, G.; Dick, H.B.; Faissner, A.; Joachim, S.C. Ischemic injury leads to extracellular matrix alterations in retina and optic nerve. Sci. Rep. 2017, 7, 43470. [Google Scholar] [CrossRef]
- Wiemann, S.; Yousf, A.; Joachim, S.C.; Peters, C.; Mueller-Buehl, A.M.; Wagner, N.; Reinhard, J. Knock-Out of Tenascin-C Ameliorates Ischemia-Induced Rod-Photoreceptor Degeneration and Retinal Dysfunction. Front. Neurosci. 2021, 15, 642176. [Google Scholar] [CrossRef] [PubMed]
- Takagi, Y.; Farrow, R.E.; Billington, N.; Nagy, A.; Batters, C.; Yang, Y.; Sellers, J.R.; Molloy, J.E. Myosin-10 produces its power-stroke in two phases and moves processively along a single actin filament under low load. Proc. Natl. Acad. Sci. USA 2014, 111, E1833–E1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.Y.; Bradley, J.M.; Keller, K.E. Phenotypic and Functional Alterations in Tunneling Nanotubes Formed by Glaucomatous Trabecular Meshwork Cells. Investig. Opthalmol. Vis. Sci. 2019, 60, 4583–4595. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.Y.; Yang, Y.F.; Keller, K.E. Myosin-X Silencing in the Trabecular Meshwork Suggests a Role for Tunneling Nanotubes in Outflow Regulation. Investig. Opthalmol. Vis. Sci. 2019, 60, 843–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, P.; Azizi, L.; Kukkurainen, S.; Gao, T.; Baikoghli, M.; Jacquier, M.-C.; Sun, Y.; Määttä, J.A.E.; Cheng, R.H.; Wehrle-Haller, B.; et al. Crystal structure of the FERM-folded talin head reveals the determinants for integrin binding. Proc. Natl. Acad. Sci. USA 2020, 117, 32402–32412. [Google Scholar] [CrossRef] [PubMed]
- Vecino, E.; Heller, J.P.; Veiga-Crespo, P.; Martin, K.R.; Fawcett, J.W. Influence of extracellular matrix components on the expression of integrins and regeneration of adult retinal ganglion cells. PLoS ONE 2015, 10, e0125250. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 7 Days after S100B Immunization | |||||
|---|---|---|---|---|---|
| Statistical Rank | Uniprot ID | Protein Name | Qvalue (FDR) ≤0.01 | p-Value ≤0.02 | Ratio ≥30% |
| 1 | Q7TNY6 | Golgi resident protein GCP60 | 1.38 × 10−3 | 0.002 | 48% |
| 2 | O35814 | Stress-induced-phosphoprotein 1 | 1.15 × 10−5 | 0.002 | 48% |
| 3 | P25809 | Creatine kinase U-type, mitochondrial | 2.07 × 10−7 | 0.004 | 35% |
| 4 | P02770 | Albumin | 1.85 × 10−10 | 0.005 | 33% |
| 5 | P56574 | Isocitrate dehydrogenase [NADP], mitochondrial | 7.62 × 10−4 | 0.006 | 53% |
| 6 | P63004 | Platelet-activating factor acetylhydrolase IB subunit alpha | 3.21 × 10−5 | 0.007 | 45% |
| 7 | P12785 | Fatty acid synthase | 2.00 × 10−6 | 0.007 | 57% |
| 8 | P49432 | Pyruvate dehydrogenase e1 component subunit beta, mitochondrial | 3.79 × 10−6 | 0.010 | 108% |
| 9 | P12346 | Serotransferrin | 2.08 × 10−8 | 0.011 | 33% |
| 10 | Q63151 | Long-chain-fatty-acid--CoA ligase 3 | 7.13 × 10−4 | 0.012 | 45% |
| 11 | P14841 | Cystatin-C | 3.09 × 10−4 | 0.012 | 61% |
| 12 | Q63598 | Plastin-3 | 1.39 × 10−3 | 0.012 | 85% |
| 13 | Q9JLT0 | Myosin-10 | 1.75 × 10−4 | 0.013 | 43 % |
| 14 | Q924S5 | Lon protease homolog, mitochondrial | 4.22 × 10−5 | 0.015 | 40% |
| 15 | O70351 | 3-hydroxyacyl-CoA dehydrogenase type-2 | 9.91 × 10−4 | 0.016 | 85% |
| 16 | Q8K1P7 | Transcription activator BRG1 | 3.13 × 10−9 | 0.018 | 55% |
| 17 | P15146 | Microtubule-associated protein 2 | 3.03 × 10−5 | 0.019 | 58% |
| 18 | Q5PPM7 | Rod outer segment membrane protein 1 | 2.64 × 10−9 | 0.021 | 51% |
| 19 | P21531 | 60S ribosomal protein L3 | 6.73 × 10−5 | 0.021 | 31% |
| 20 | Q5XI73 | Rho GDP-dissociation inhibitor 1 | 5.02 × 10−7 | 0.021 | 77% |
| 14 Days after S100B Immunization | |||||
|---|---|---|---|---|---|
| Statistical Rank | Uniprot ID | Protein Name | Qvalue (FDR) ≤0.01 | p-Value ≤0.02 | Ratio ≥30% |
| 1 | Q5XI31 | GPI transamidase component PIG-S | 3.05 × 10−3 | 0.003 | 36% |
| 2 | P11506 | Plasma membrane calcium-transporting ATPase 2 | 1.37 × 10−8 | 0.003 | 54% |
| 3 | P41542 | General vesicular transport factor p115 | 1.81 × 10−4 | 0.003 | 46% |
| 4 | P63039 | 60 kDa heat shock protein, mitochondrial | 4.80 × 10−11 | 0.004 | 45% |
| 5 | P21670 | Proteasome subunit alpha type-4 | 2.04 × 10−7 | 0.005 | 69% |
| 6 | Q9JK11 | Reticulon-4 | 3.50 × 10−9 | 0.010 | 37% |
| 7 | Q62927 | cGMP-gated cation channel alpha-1 | 5.20 × 10−6 | 0.011 | 30% |
| 8 | Q9ePH8 | Polyadenylate-binding protein 1 | 1.99 × 10−7 | 0.012 | 59% |
| 9 | Q6IMY8 | Heterogeneous nuclear ribonucleoprotein U | 4.57 × 10−9 | 0.012 | 109% |
| 10 | P14881 | Beta-crystallin A3 | 2.14 × 10−10 | 0.013 | 298% |
| 11 | D3ZLZ7 | Inosine-5’-monophosphate dehydrogenase 1 | 5.11 × 10−10 | 0.015 | 32% |
| 12 | O35476 | Medium-wave-sensitive opsin 1 | 1.18 × 10−4 | 0.016 | 152% |
| 13 | P63012 | Ras-related protein Rab-3A | 1.34 × 10−8 | 0.016 | 41% |
| 14 | P84083 | ADP-ribosylation factor 5 | 9.07 × 10−8 | 0.017 | 37% |
| 15 | P06761 | endoplasmic reticulum chaperone BiP | 8.01 × 10−9 | 0.018 | 71% |
| 16 | P02770 | Albumin | 3.75 × 10−11 | 0.018 | 45% |
| 17 | D3ZHV2 | Microtubule-actin cross-linking factor 1 | 2.31 × 10−4 | 0.018 | 37% |
| 18 | P62752 | 60S ribosomal protein L23a | 4.00 × 10−4 | 0.019 | 42% |
| 19 | Q68FX0 | Isocitrate dehydrogenase [NAD] subunit beta, mitochondrial | 4.00 × 10−7 | 0.020 | 32% |
| 20 | P50475 | Alanine--tRNA ligase, cytoplasmic | 5.49 × 10−8 | 0.020 | 78% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reinehr, S.; Guntermann, A.; Theile, J.; Benning, L.; Grotegut, P.; Kuehn, S.; Serschnitzki, B.; Dick, H.B.; Marcus, K.; Joachim, S.C.; et al. Proteomic Analysis of Retinal Tissue in an S100B Autoimmune Glaucoma Model. Biology 2022, 11, 16. https://doi.org/10.3390/biology11010016
Reinehr S, Guntermann A, Theile J, Benning L, Grotegut P, Kuehn S, Serschnitzki B, Dick HB, Marcus K, Joachim SC, et al. Proteomic Analysis of Retinal Tissue in an S100B Autoimmune Glaucoma Model. Biology. 2022; 11(1):16. https://doi.org/10.3390/biology11010016
Chicago/Turabian StyleReinehr, Sabrina, Annika Guntermann, Janine Theile, Lara Benning, Pia Grotegut, Sandra Kuehn, Bettina Serschnitzki, H. Burkhard Dick, Katrin Marcus, Stephanie C. Joachim, and et al. 2022. "Proteomic Analysis of Retinal Tissue in an S100B Autoimmune Glaucoma Model" Biology 11, no. 1: 16. https://doi.org/10.3390/biology11010016
APA StyleReinehr, S., Guntermann, A., Theile, J., Benning, L., Grotegut, P., Kuehn, S., Serschnitzki, B., Dick, H. B., Marcus, K., Joachim, S. C., & May, C. (2022). Proteomic Analysis of Retinal Tissue in an S100B Autoimmune Glaucoma Model. Biology, 11(1), 16. https://doi.org/10.3390/biology11010016