
Effects of Chlorophyll a and b in Reducing Genotoxicity of 2-Amino-3,8-dimethylimidazo[4,5-F]quinoxaline (MeIQx)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Drosophila Strains
2.3. Experimental Procedure
2.3.1. MeIQx and Chlorophyll a and Chlorophyll b Co-Treatment Applications
2.3.2. Chlorophyll a and Chlorophyll b Pre-Treatment Applications
2.4. Preparation and Microscopic Analysis of the Wings
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Robbana-Barnat, S.; Rabache, M.; Rialland, E.; Fradin, J. Heterocyclic amines: Occurrence and prevention in cooked food. Environ. Health Perspect. 1996, 104, 280–288. [Google Scholar] [CrossRef] [PubMed]
- Jägerstad, M.; Skog, K. Genotoxicity of heat-processed foods. Mutat. Res. 2005, 574, 156–172. [Google Scholar] [CrossRef] [PubMed]
- Skog, K.I.; Johansson, M.A.; Jägerstad, M.I. Carcinogenic heterocyclic amines in model systems and cooked foods: A review on formation, occurrence and intake. Food Chem. Toxicol. 1998, 36, 879–896. [Google Scholar] [CrossRef]
- Akram, M.; Riaz, M.; Wadood, A.; Wadood, C.; Hazrat, A.; Mukhtiar, M.; Zakki, S.A.; Daniyal, M.; Shariati, M.A.F.; Khan, S.; et al. Medicinal plants with anti-mutagenic potential. Biotechnol. Biotechnol. Equip. 2020, 34, 309–318. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.; Lin, Q.; Liang, Y. Plant-derived antioxidants protect the nervous system from aging by inhibiting oxidative stress. Front. Aging Neurosci. 2020, 12, 209. [Google Scholar] [CrossRef]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An overview on the role of dietary phenolics for the treatment of cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef] [Green Version]
- Block, G. Vitamin C and cancer prevention: The epidemiological evidence. Am. J. Clin. Nutr. 1991, 53, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Byers, T.; Perry, G. Dietary carotenes, vitamin C and vitamin E as protective antioxidant in human cancer. Ann. Rev. Nutr. 1992, 12, 139–159. [Google Scholar] [CrossRef]
- De Flora, S.; Izzotti, A.D.; D’agostini, F.; Balansky, R.M.; Noonan, D.; Albini, A. Multiple point of intervention in the prevention of cancer and other mutation-related diseases. Mutat. Res. 2001, 480, 9–22. [Google Scholar] [CrossRef]
- Fabiani, R.; Bartelomeo, A.; Rosignoli, P.; Morozzi, G. Antioxidants prevent the lymphocyte DNA damage induced by PMA-stimulated monocytes. Nutr. Cancer 2001, 39, 284–291. [Google Scholar] [CrossRef]
- Promraksa, B.; Phetcharaburani, J.; Namwat, N.; Techasen, A.; Boonsiri, P.; Loilome, W. Evaluation of anticancer potential of Thai medicinal herb extracts against cholangiocarcinoma cell lines. PLoS ONE 2019, 14, e0216721. [Google Scholar] [CrossRef] [PubMed]
- Bernards, A.; Hariharan, I.K. Of flies and men- studying human disease in Drosophila. Curr. Opin. Genet Dev. 2001, 11, 274–278. [Google Scholar] [CrossRef]
- Boulet, M.; Miller, M.; Vandel, L.; Waltzer, L. From Drosophila blood cells to human leukemia. Adv. Exp. Med. Biol. 2018, 1076, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Link, N.; Bellen, H.J. Using Drosophila to drive the diagnosis and understand the mechanisms of rare human diseases. Development 2020, 147, dev191411. [Google Scholar] [CrossRef] [PubMed]
- Lindsley, D.L.; Zimm, G.G. The Genome of Drosophila Melanogaster; Academic Press: San Diago, CA, USA, 1992; p. 1133. [Google Scholar]
- Graf, U.; Wurgler, F.E.; Katz, A.J.; Frei, H.; Juan, H.; Hall, C.B.; Kale, P.G. Somatic mutation and recombination test in Drosophila melanogaster. Environ. Mutagenes. 1984, 6, 153–188. [Google Scholar] [CrossRef]
- Demir, E.; Kocaoglu, S.; Kaya, B. Antigenotoxic properties of chlorophyllin and chlorophylls in the Drosophila wing spot test. Fresen. Environ. Bull. 2010, 19, 3131–3138. [Google Scholar]
- Demir, E.; Kaya, B.; Kocaoglu Cenkci, S. Antigenotoxic activities of ascorbic acid, chlorophyll a and chlorophyll b in acrolein and malondialdehyde-ınduced genotoxicity in Drosophila melanogaster. Ekoloji 2013, 22, 36–42. [Google Scholar] [CrossRef]
- Marcos, R.; Carmona, E.R. The wing-spot and the comet tests as useful assays detecting genotoxicity in Drosophila. Methods Mol. Biol. 2013, 1044, 417–427. [Google Scholar]
- Frei, H.; Wurgler, F.E. Statistical methods to decide whether mutagenic test data from Drosophila assays indicate a positive, negative or inconclusive result. Mutat. Res. 1988, 203, 297–308. [Google Scholar] [CrossRef]
- Kastenbaum, M.A.; Bowman, K.O. Tables for determining the statistical significance of mutation frequencies. Mutat. Res. 1970, 9, 527–549. [Google Scholar]
- Abraham, S.K. Antigenotoxicity of coffee in the Drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Graf, U.; Van Schaik, N. Improved high bioactivation cross for the wing somatic mutation and recombination test in Drosophila melanogaster. Mutat. Res. 1992, 271, 59–67. [Google Scholar] [CrossRef]
- De Morrais, C.R.; Carvalho, S.M.; Carvalho Naves, M.P.; Araujo, G.; De Rezende, A.A.A.; Bonetti, A.M.; Spano, M.A. Mutagenic, recombinogenic and carcinogenic potential of thiamethoxam insecticide and formulated product in somatic cells of Drosophila Melanogaster. Chemosphere 2017, 187, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Schut, H.A.J.; Snyderwıne, E.G. DNA adducts of heterocyclic amine food mutagens: Implications for mutagenesis and carcinogenesis. Carcinogenesis 1999, 20, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Sınha, R. An epidemiologic approach to studying heterocyclic amines. Mutat. Res. 2002, 506–507, 197–204. [Google Scholar] [CrossRef]
- Gooderham, N.J.; Murray, S.; Lynch, A.M.; Yadollahi-Farsani, M.; Zhao, K.; Boobis, A.R.; Davies, D.S. Food-derived heterocyclic amine mutagens: Variable metabolism and significance to humans. Drug Metab. Dispos. 2001, 29, 529–534. [Google Scholar] [PubMed]
- Boobıs, A.R.; Lynch, A.M.; Murray, S.; De La Torre, R.; Solans, A.; Farr, M.; Segura, J.; Gooderham, N.J.; Davıes, D.S. CYP1A2-catalyzed conversion of dietary heterocyclic amines to their Proximate carcinogens is their major route of metabolism in humans. Cancer Res. 1994, 54, 89–94. [Google Scholar]
- Shin, A.; Shrubsole, M.J.; Ness, R.M.; Wu, H.; Sinha, R.; Smalley, W.E.; Shyr, Y.; Zheng, W. Meat and meat-mutagen intake, doneness preference and the risk of colorectal polyps: The Tennessee colorectal polyp study. Int. J. Cancer 2007, 121, 136–142. [Google Scholar] [CrossRef]
- National Research Council Committee on Chemical Environmental Mutagens. Quantitative Relationship between Mutagenic and Carcinogenic Potencies: A Feasibility Study; National Academy Press: Washington, DC, USA, 1983; p. 37. [Google Scholar]
- Durling, L.J.K.; Abramsson-Zetterberg, L. A comparison of genotoxicity between three common heterocyclic amines and acrylamide. Mutat. Res. 2005, 580, 103–110. [Google Scholar] [CrossRef]
- Heddle, J.A.; Knize, M.G.; Dawod, D.; Zhang, X.B. A Test of the Mutagenicity of Cooked Meats in vivo. Mutagenesis 2001, 16, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Itoh, T.; Kuwahara, T.; Suzuki, T.; Hayashi, M.; Ohnishi, Y. Regional mutagenicity of heterocyclic amines in the intestine: Mutation analysis of the cII gene in lambda/lacZ transgenic mice. Mutat. Res. 2003, 539, 99–108. [Google Scholar] [CrossRef]
- Knasmuller, S.; Schwab, C.E.; Land, S.J.; Wang, C.Y.; Sanyal, R.; Kundi, M.; Parzefall, W.; Darroudi, F. Genotoxic effects of heterocyclic aromatic amines in human derived hepatoma (HepG2) cells. Mutagenesis 1999, 14, 533–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knize, M.G.; Salmon, C.P.; Felton, J.S. Mutagenic activity and heterocyclic amine carcinogens in commercial pet foods. Mutat. Res. 2003, 539, 195–201. [Google Scholar] [CrossRef]
- Lynch, A.M.; Gooderham, N.J.; Davies, D.; Boobis, A.R. Genetic analysis of pHIP intestinal mutations in Muta™Mouse. Mutagenesis 1998, 13, 601–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfau, W.; Martin, F.L.; Cole, K.J.; Venitt, S.; Phillips, D.H.; Grover, P.L.; Marquardt, H. Heterocyclic aromatic amines induce DNA strand breaks and cell transformation. Carcinogenesis 1999, 20, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Akanuma, M.; Shimoi, K.; Kinae, N.; Ohta, T. Food-derived heterocyclic amines potentiate the mutagenicity of a drinking water mutagen 3-chloro-4-(dichloromethyl)-5-hydroxy-2(5H)-furanone (MX). Mutat. Res. 1997, 377, 225–229. [Google Scholar] [CrossRef]
- Puri, B.K.; Ijeh, C.; Monro, J.A. Removal of DNA adducts. Med. Hypotheses 2019, 127, 11–14. [Google Scholar] [CrossRef]
- Jägerstad, M.; Skog, K.; Arvıdsson, P.; Solyakov, A. Chemistry, formation and occurrence of genotoxic heterocyclic amines identified in model systems and cooked foods. Z. Lebensm.-Forsch. A 1998, 207, 419–427. [Google Scholar] [CrossRef]
- Jackson, L.S.; Hargraves, W.A. effects of time and temperature on the formation of MeIQ, and DiMeIQ, in a model system containing threonine, glucose, and creatine. J. Agric. Food Chem. 1995, 43, 1678–1684. [Google Scholar] [CrossRef]
- Skog, K.; Knize, M.; Felton, J.S.; Jägerstad, M. Formation of new heterocyclic amine mutagens by heating creatinine, alanine, threonine and glucose. Mutat. Res. 1992, 268, 191–197. [Google Scholar] [CrossRef]
- Persson, E.; Sjöholm, I.; Skog, K. Effect of high water-holding capacity on the formation of heterocyclic amines in fried beefburgers. J. Agric. Food Chem. 2003, 51, 4472–4477. [Google Scholar] [CrossRef] [PubMed]
- Puangsombat, K.; Smith, J.S. Inhibition of heterocyclic amine formation in beef patties by ethanolic extracts of rosemary. J. Food Sci. 2010, 75, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Balogh, Z.; Gray, J.I.; Gomaa, E.A.; Booren, A.M. formation and inhibition of heterocyclic aromatic amines in fried ground beef patties. Food Chem. Toxicol. 2000, 38, 395–401. [Google Scholar] [CrossRef]
- Cheng, K.W.; Wu, Q.; Zheng, Z.P.; Peng, X.; Simon, J.E.; Chen, F.; Wang, M. Inhibitory effect of fruit extracts on the formation of heterocyclic amines. J. Agric. Food Chem. 2007, 55, 10359–61035. [Google Scholar] [CrossRef] [PubMed]
- Monti, S.M.; Ritieni, A.; Sacchi, R.; Skog, K.; Borgen, E.; Fogliano, V. Characterization of phenolic compounds in virgin olive oil and their effect on the formation of carcinogenic/mutagenic heterocyclic amines in a model system. J. Agric. Food Chem. 2001, 49, 3969–3975. [Google Scholar] [CrossRef]
- Murkovic, M.; Steinberger, D.; Pfannhauser, W. Antioxidant spices reduce the formation of heterocyclic amines in fried meat. Z. Lebensm. Forsch. A 1998, 207, 477–480. [Google Scholar] [CrossRef]
- Salmon, C.P.; Knize, M.G.; Felton, J.S. Effects of marinating on heterocyclic amine carcinogen formation in grilled chicken. Food Chem. Toxicol. 1997, 35, 433–441. [Google Scholar] [CrossRef]
- Vitaglione, P.; Monti, S.M.; Ambrosino, P.; Skog, K.; Fogliano, V. Carotenoids from tomatoes inhibit heterocyclic amine formation. Eur. Food Res. Technol. 2002, 215, 108–113. [Google Scholar] [CrossRef]
- Taira, K.; Miyashita, Y.; Okamoto, K.; Arimoto, S.; Takahashi, E.; Negishi, T. Novel antimutagenic factors derived from the edible mushroom Agrocybe cylindracea. Mutat. Res. 2005, 586, 115–123. [Google Scholar] [CrossRef]
- Takahashi, E.; Marczylo, T.H.; Watanabe, T.; Nagai, S.; Hayatsu, H.; Negishi, T. Preventive effects of anthraquinone food pigments on the DNA damage induced by carcinogens in Drosophila. Mutat. Res. 2001, 480–481, 139–145. [Google Scholar] [CrossRef]
- Nakahara, K.; Roy, M.K.; Ono, H.; Maeda, I.; Ohnishi-Kameyama, M.; Yoshida, M.; Trakoontivakorn, G. Antimutagenic constituents against heterocyclic amines. J. Agric. Food Chem. 2003, 51, 6456–6460. [Google Scholar] [CrossRef] [PubMed]
- Egner, P.A.; Wang, J.B.; Zhu, Y.R.; Zhang, B.C.; Wu, Y.; Zhang, Q.N.; Qian, G.S.; Kuang, S.Y.; Gange, S.J.; Jacobson, L.P.; et al. Chlorophyllin intervention reduces aflatoxin–DNA adducts in individuals at high risk for liver cancer. Med. Sci. 2001, 98, 14601–14606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Vogel, J.; Jonker-Termont, D.S.; Katan, M.B.; Van Der Meer, R. Natural chlorophyll but not chlorophyllin prevents heme-induced cytotoxic and hyperproliferative effects in rat colon. J. Nutr. 2005, 135, 1995–2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arimoto, S.; Fukuoka, S.; Itome, C.; Nakano, H.; Rai, H.; Hayatsu, H. Binding of polycyclic planar mutagens to chlorophyllin resulting in inhibition of the mutagenic activity. Mutat. Res. 1993, 287, 293–305. [Google Scholar] [CrossRef]
- Dashwood, R.; Liew, C. Chlorophyllin-enhanced excretion of urinary and fecal mutagens in rats given 2-amino-3- methylimidazo [4,5-f]quinoline. Environ. Mol. Mutagenes. 1992, 20, 199–205. [Google Scholar] [CrossRef]
- Simonich, M.T.; Egner, P.A.; Roebuck, B.D.; Orner, G.A.; Jubert, C.; Pereira, C.; Groopman, J.D.; Kensler, T.W.; Dashwood, R.H.; Williams, D.E.; et al. Natural chlorophyll inhibits aflatoxin B1-induced multi-organ carcinogenesis in the rat. Carcinogenesis 2007, 28, 1294–1302. [Google Scholar] [CrossRef] [Green Version]
- Negishi, T.; Arimoto, S.; Nishizaki, C.; Hayatsu, H. Inhibitory effect of chlorophyll on the genotoxicity of 3-amino-1-methyl-5H-pyrido [4,3-b indole (Trp-P-2). Carcinogenesis 1989, 10, 145–149. [Google Scholar] [CrossRef]
- Negishi, T.; Rai, H.; Hayatsu, H. Antigenotoxic activity of natural chlorophylls. Mutat. Res. 1997, 376, 97–100. [Google Scholar] [CrossRef]
- Jubert, C.; Mata, J.; Bench, G.; Dashwood, R.; Pereira, C.; Tracewell, W.; Turteltaub, K.; Williams, D.; Bailey, G. Effects of chlorophyll and chlorophyllin on low-dose aflatoxin b1 pharmacokinetics in human volunteers. Cancer Prev. Res. 2009, 2, 1015–1022. [Google Scholar] [CrossRef] [Green Version]
- Demir, E.; Kocaoglu, S.; Kaya, B. Protective effects of chlorophyll against the genotoxicity of uvb in Drosophila smart assay. Fresen. Environ. Bull. 2008, 17, 2180–2186. [Google Scholar]
- Demir, E.; Kocaoglu, S.; Kaya, B. Protection against ultraviolet b-induced genotoxicity by the chlorophyllin in Drosoph melanogaster. Fresen. Environ. Bull. 2008, 17, 2187–2192. [Google Scholar]
- Pimentel, E.; Cruces, M.P.; Zimmering, S. On the persistence of the radioprotective effect of chlorophyllin (CHLN) in somatic cells of Drosophila. Mutat. Res. 1999, 446, 189–192. [Google Scholar] [CrossRef]
- Tajmir-Riahi, H.A.; Neault, J.-F.; Diamantoglou, S. DNA adducts with chlorophyll and chlorophyllin as antimutagenic agents: Synthesis, stability, and structural features. Methods Mol. Biol. 2004, 274, 159–171. [Google Scholar] [PubMed]
- Ferruzzi, M.G.; Bohm, V.; Courtney, P.D.; Schwartz, S.J. Antioxidant and antimutagenic activity of dietary chlorophyll derivatives determined by radical scavenging and bacterial reverse mutagenesis assays. J. Food Sci. 2002, 67, 2589–2595. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Yang, C.M.; Chen, C.M.; Chao, P.Y.; Hu, S.P. Effects of chlorophyll-related compounds on hydrogen peroxide induced dna damage within human lymphocytes. J. Agric. Food Chem. 2005, 53, 2746–2750. [Google Scholar] [CrossRef]
- De Cássia Bez, G.; Jordäol, B.Q.; Vicentini, V.E.P.; Mantovani, M.S. Differential protection of a and b chlorophyll in mmc-mediated induction of chromatid and lsochromatid breaks in human lymphocytes culture. Cytologia 2001, 66, 313–318. [Google Scholar]
- McQuistan, T.J.; Simonich, M.T.; Pratt, M.M.; Pereira, C.B.; Hendricks, J.D.; Dashwood, R.H.; Williams, D.E.; Bailey, G.S. Cancer chemoprevention by dietary chlorophylls: A 12,000-animal dose–dose matrix biomarker and tumor study. Food Chem. Toxicol. 2012, 50, 341–352. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Lee, D.H.; Won, C.H.; Kim, S.M.; Lee, S.; Lee, M.J.; Chung, J.H. Drink containing chlorophyll extracts improves signs of photoaging and increases type I procollagen in human skin in vivo. Korean J. Investig. Dermatol. 2006, 13, 111–119. [Google Scholar]



{kind=link}
{kind=link}
| Doses | No. of Wings (N) | Small Single Spots (1–2 Cells) (m = 2) | Large Single Spots (>2 Cells) (m = 5) | Twin Spots (m = 5) | Total mwh Spots a (m = 2) | Total Spots (m = 2) | Frequency of Clone Formation per 105 Cells b | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mwh/flr3 wings | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | ||
| 72 ± 4 h treatments | |||||||||||||||||
| Distilled water | 80 | 16 | 0.20 | 1 | 0.01 | 0 | 0.00 | 17 | 0.21 | 17 | 0.21 | 0.87 | |||||
| 1 mM EMS | 80 | 163 | 2.04 | + | 89 | 1.11 | + | 32 | 0.40 | + | 273 | 3.41 | + | 284 | 3.55 | + | 13.99 |
| %0.5 DMSO | 80 | 23 | 0.29 | - | 3 | 0.04 | i | 0 | 0.00 | i | 26 | 0.33 | i | 26 | 0.33 | i | 1.33 |
| Chlorophyll a 0.5 | 80 | 14 | 0.18 | - | 4 | 0.05 | i | 2 | 0.03 | i | 17 | 0.21 | - | 17 | 0.21 | - | 0.87 |
| Chlorophyll a 1 | 80 | 20 | 0.25 | - | 2 | 0.03 | - | 3 | 0.04 | i | 25 | 0.31 | - | 25 | 0.31 | - | 1.28 |
| Chlorophyll b 0.5 | 80 | 19 | 0.24 | - | 5 | 0.06 | i | 2 | 0.03 | i | 26 | 0.33 | - | 26 | 0.33 | - | 1.33 |
| Chlorophyll b 1 | 80 | 23 | 0.29 | - | 3 | 0.03 | i | 0 | 0.00 | i | 27 | 0.34 | - | 27 | 0.34 | - | 1.38 |
| 48 ± 4 h treatments | |||||||||||||||||
| Distilled water | 80 | 17 | 0.21 | 0 | 0.00 | 1 | 0.01 | 18 | 0.23 | 18 | 0.23 | 0.92 | |||||
| %0.5 DMSO | 80 | 22 | 0.26 | i | 1 | 0.01 | i | 0 | 0.00 | i | 22 | 0.26 | i | 23 | 0.29 | i | 1.13 |
| Chlorophyll a 0.5 | 80 | 11 | 0.14 | - | 0 | 0.00 | i | 0 | 0.00 | i | 11 | 0.14 | - | 11 | 0.14 | - | 0.56 |
| Chlorophyll a 1 | 80 | 10 | 0.13 | - | 2 | 0.03 | i | 0 | 0.00 | i | 12 | 0.15 | - | 12 | 0.15 | - | 0.61 |
| Chlorophyll b 0.5 | 80 | 8 | 0.10 | - | 1 | 0.01 | i | 0 | 0.00 | i | 9 | 0.11 | - | 9 | 0.11 | - | 0.46 |
| Chlorophyll b 1 | 80 | 11 | 0.14 | - | 1 | 0.01 | i | 0 | 0.00 | i | 12 | 0.15 | - | 12 | 0.15 | - | 0.61 |
| Doses | No. of Wings (N) | Small Single Spots (1–2 Cells) (m = 2) | Large Single Spots (>2 Cells) (m = 5) | Twin Spots (m = 5) | Total mwh Spots a (m = 2) | Total Spots (m = 2) | Frequency of Clone Formation per 105 Cells b | Recombination c (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mwh/flr3 wings | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | |||
| % 0.5 DMSO | 80 | 23 | 0.29 | - | 3 | 0.04 | i | 0 | 0.00 | i | 26 | 0.33 | i | 26 | 0.33 | i | 1.33 | |
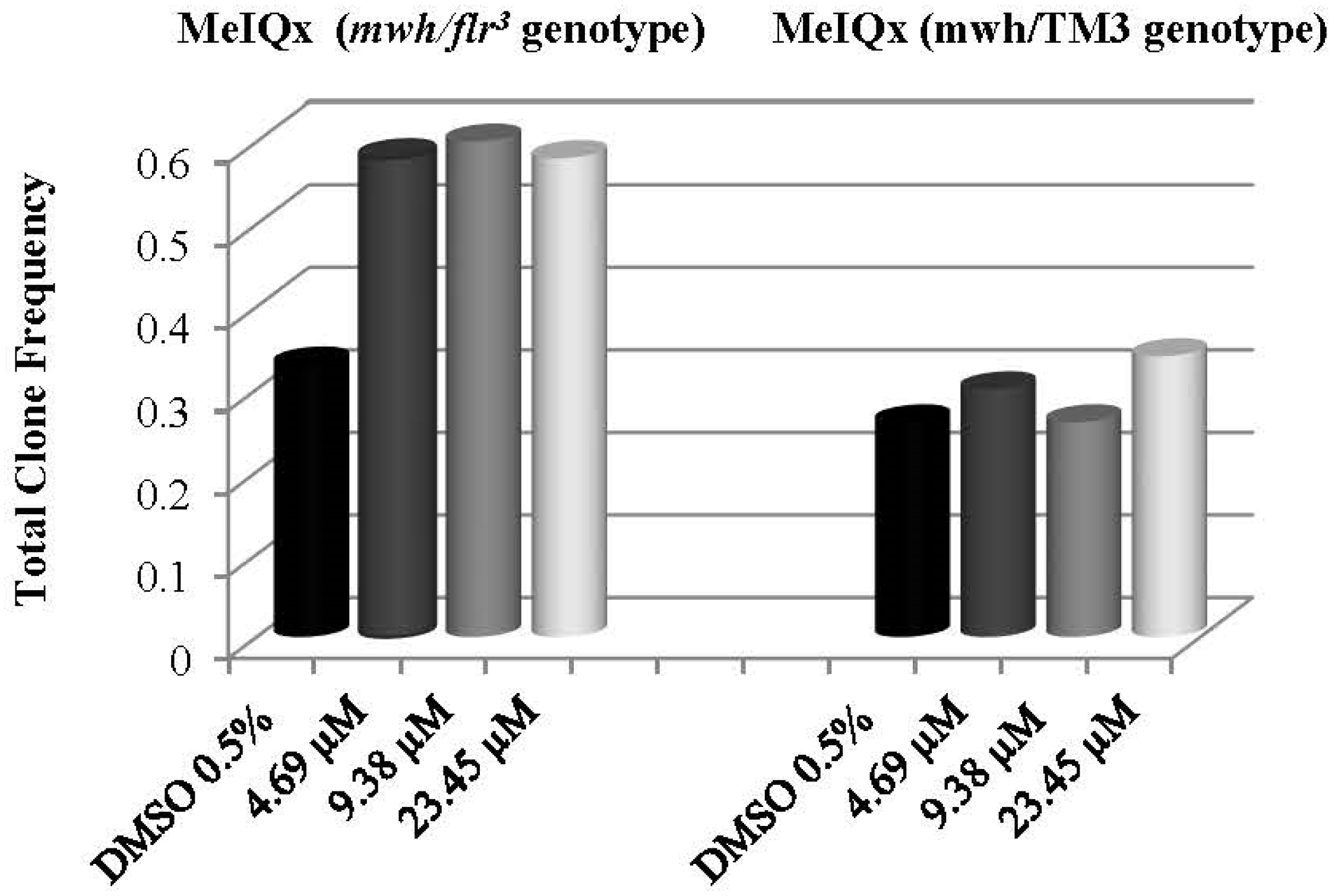
| 4.69 µM | 80 | 40 | 0.43 | + | 5 | 0.05 | i | 1 | 0.01 | i | 46 | 0.50 | + | 46 | 0.58 | + | 2.04 | 40 |
| 9.38 µM | 80 | 42 | 0.53 | + | 6 | 0.08 | i | 0 | 0.00 | i | 48 | 0.60 | + | 48 | 0.60 | + | 2.46 | 56,67 |
| 23.45 µM | 80 | 41 | 0.39 | + | 4 | 0.04 | i | 1 | 0.01 | i | 46 | 0.50 | + | 46 | 0.58 | + | 2.04 | 32 |
| mwh/TM3 wings | ||||||||||||||||||
| % 0.5 DMSO | 80 | 19 | 0.24 | - | 2 | 0.03 | - | 21 | 0.26 | - | 21 | 0.26 | - | 1.08 | ||||
| 4.69 µM | 80 | 23 | 0.29 | i | 1 | 0.01 | i | 24 | 0.30 | i | 24 | 0.30 | i | 1.23 | ||||
| 9.38 µM | 80 | 20 | 0.25 | i | 1 | 0.01 | i | 21 | 0.26 | i | 21 | 0.26 | - | 1.08 | ||||
| 23.45 µM | 80 | 26 | 0.33 | i | 1 | 0.01 | i | 27 | 0.34 | i | 27 | 0.34 | i | 1.38 | ||||
| Doses | No. of Wings (N) | Small Single Spots (1–2 Cells) (m = 2) | Large Single Spots (>2 Cells) (m = 5) | Twin Spots (m = 5) | Total mwh Spots a (m = 2) | Total Spots (m = 2) | Frequency of Clone Formation per 105 Cells b | Percentage of Inhibition (↓) and Induction (↑) (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mwh/flr3 wings | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | |||
| %0.5 DMSO | 80 | 23 | 0.29 | i | 3 | 0.04 | i | 0 | 0.00 | i | 26 | 0.33 | i | 26 | 0.33 | i | 1.54 | |
| 4.69 µM | 80 | 40 | 0.50 | i | 5 | 0.06 | i | 1 | 0.01 | i | 46 | 0.58 | + | 46 | 0.58 | + | 2.36 | |
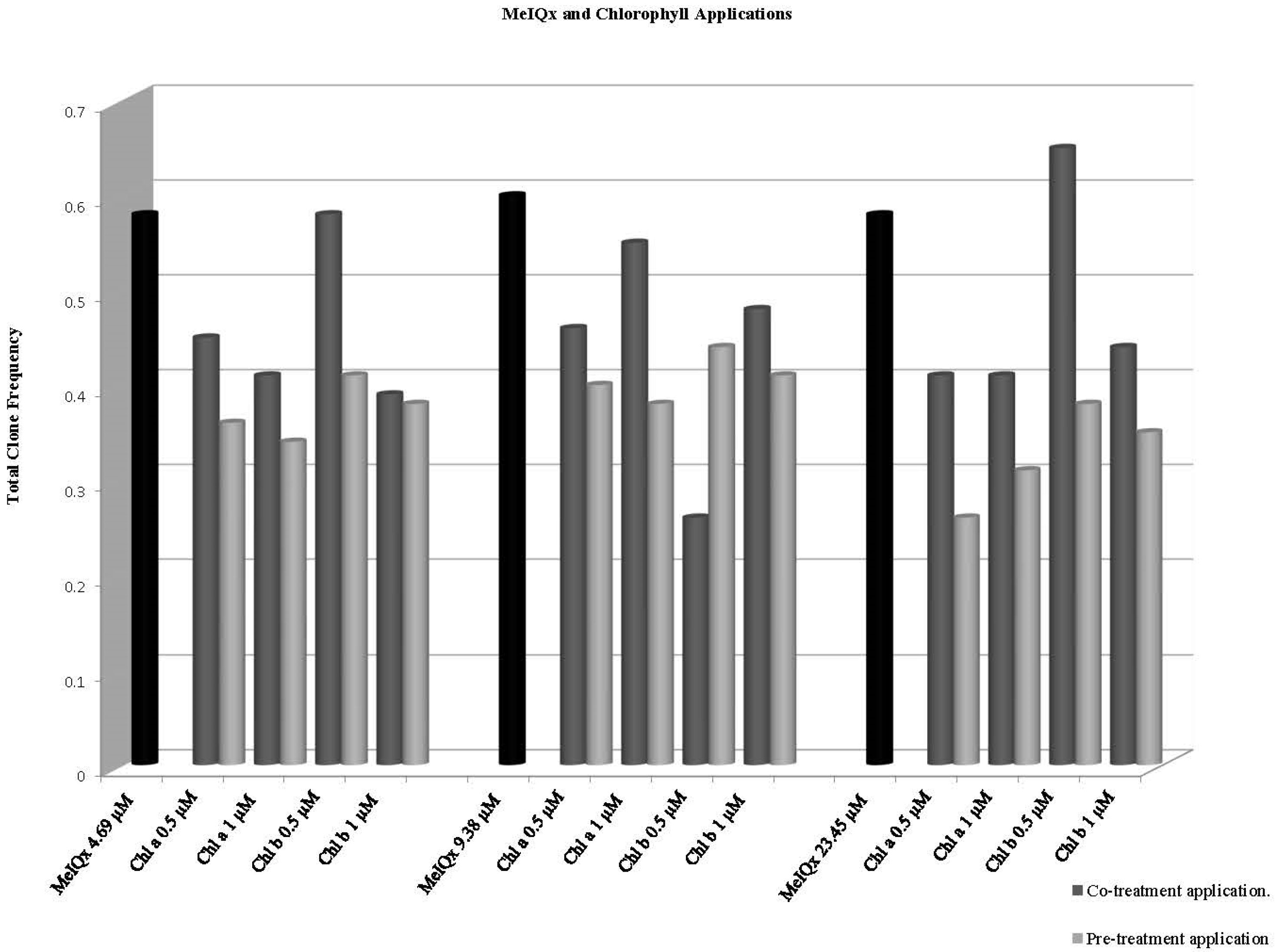
| Chlorophyll a 0.5 | 80 | 32 | 0.40 | i | 3 | 0.04 | - | 0 | 0.00 | i | 35 | 0.44 | - | 35 | 0.44 | - | 1.79 | ↓24.14 |
| Chlorophyll a 1 | 80 | 37 | 0.46 | - | 3 | 0.04 | - | 0 | 0.00 | i | 40 | 0.50 | - | 33 | 0.41 | - | 2.05 | ↓13.79 |
| Chlorophyll b 0.5 | 80 | 42 | 0.53 | i | 4 | 0.05 | i | 0 | 0.00 | i | 45 | 0.56 | - | 46 | 0.58 | i | 2.30 | 0 |
| Chlorophyll b 1 | 80 | 28 | 0.35 | - | 3 | 0.04 | - | 0 | 0.00 | i | 31 | 0.39 | - | 31 | 0.39 | - | 1.58 | ↓32.76 |
| 9.38 µM | 80 | 42 | 0.53 | + | 6 | 0.08 | i | 0 | 0.00 | i | 48 | 0.60 | + | 48 | 0.60 | + | 2.46 | |
| Chlorophyll a 0.5 | 80 | 33 | 0.41 | - | 5 | 0.06 | - | 0 | 0.00 | i | 36 | 0.45 | - | 38 | 0.48 | - | 1.84 | ↓17.24 |
| Chlorophyll a 1 | 80 | 43 | 0.54 | i | 1 | 0.01 | - | 0 | 0.00 | i | 43 | 0.54 | - | 44 | 0.55 | - | 2.25 | ↓5.17 |
| Chlorophyll b 0.5 | 80 | 21 | 0.26 | - | 0 | 0.00 | - | 0 | 0.00 | i | 21 | 0.26 | - | 21 | 0.26 | - | 1.07 | ↓55.17 |
| Chlorophyll b 1 | 80 | 37 | 0.46 | - | 1 | 0.01 | - | 0 | 0.00 | i | 38 | 0.48 | - | 38 | 0.48 | - | 1.94 | ↓17.24 |
| 23.45 µM | 80 | 41 | 0.51 | i | 4 | 0.05 | i | 1 | 0.01 | i | 46 | 0.58 | + | 46 | 0.58 | + | 2.36 | |
| Chlorophyll a 0.5 | 80 | 25 | 0.31 | - | 7 | 0.09 | i | 1 | 0.01 | i | 33 | 0.41 | - | 33 | 0.41 | - | 1.69 | ↓29.31 |
| Chlorophyll a 1 | 80 | 31 | 0.39 | i | 2 | 0.03 | - | 0 | 0.00 | i | 33 | 0.41 | - | 33 | 0.41 | - | 1.69 | ↓29.31 |
| Chlorophyll b 0.5 | 80 | 50 | 0.63 | i | 1 | 0.01 | - | 1 | 0.01 | i | 52 | 0.65 | i | 52 | 0.65 | i | 2.66 | ↑12.06 |
| Chlorophyll b 1 | 80 | 35 | 0.44 | i | 3 | 0.04 | - | 1 | 0.01 | i | 39 | 0.49 | - | 39 | 0.49 | - | 2.00 | ↓15.52 |
| Doses | No. of Wings (N) | Small Single Spots (1–2 Cells) (m = 2) | Large Single Spots (>2 Cells) (m = 5) | Twin Spots (m = 5) | Total mwh Spots (m = 2) a | Total Spots (m = 2) | Frequency of Clone Formation per 105 Cells b | Percentage of Inhibition (↓) and Induction (↑) (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mwh/flr3 wings | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | No | Fr | D | |||
| % 0.5 DMSO | 80 | 23 | 0.29 | i | 3 | 0.04 | i | 0 | 0.00 | i | 26 | 0.33 | i | 26 | 0.33 | i | 1.54 | |
| 4.69 µM | 80 | 40 | 0.50 | i | 5 | 0.06 | i | 1 | 0.01 | i | 46 | 0.58 | + | 46 | 0.58 | + | 2.36 | |
| Chlorophyll a 0.5 | 80 | 31 | 0.39 | - | 1 | 0.01 | - | 0 | 0.00 | i | 32 | 0.40 | - | 32 | 0.40 | - | 1.63 | ↓31.03 |
| Chlorophyll a 1 | 80 | 24 | 0.30 | - | 4 | 0.05 | - | 0 | 0.00 | i | 28 | 0.35 | - | 28 | 0.35 | - | 1.43 | ↓39.66 |
| Chlorophyll b 0.5 | 80 | 24 | 0.30 | - | 3 | 0.04 | - | 0 | 0.00 | i | 27 | 0.34 | - | 27 | 0.34 | - | 1.38 | ↓41.38 |
| Chlorophyll b 1 | 80 | 17 | 0.21 | - | 1 | 0.01 | - | 1 | 0.01 | i | 19 | 0.24 | - | 19 | 0.24 | - | 0.97 | ↓58.62 |
| 9.38 µM | 80 | 42 | 0.53 | + | 6 | 0.08 | i | 0 | 0.00 | i | 48 | 0.60 | + | 48 | 0.60 | + | 2.46 | |
| Chlorophyll a 0.5 | 80 | 25 | 0.31 | - | 0 | 0.00 | - | 1 | 0.01 | i | 26 | 0.33 | - | 26 | 0.33 | - | 1.33 | ↓43.10 |
| Chlorophyll a 1 | 80 | 22 | 0.28 | - | 1 | 0.01 | - | 0 | 0.00 | i | 23 | 0.29 | - | 23 | 0.29 | - | 1.18 | ↓50 |
| Chlorophyll b 0.5 | 80 | 26 | 0.33 | - | 5 | 0.06 | - | 1 | 0.01 | i | 32 | 0.40 | - | 32 | 0.40 | - | 1.63 | ↓31.03 |
| Chlorophyll b 1 | 80 | 21 | 0.26 | - | 0 | 0.00 | - | 0 | 0.00 | i | 21 | 0.26 | - | 21 | 0.26 | - | 1.07 | ↓55.17 |
| 23.45 µM | 80 | 41 | 0.51 | i | 4 | 0.05 | i | 1 | 0.01 | i | 46 | 0.58 | + | 46 | 0.58 | + | 2.36 | |
| Chlorophyll a 0.5 | 80 | 24 | 0.30 | - | 1 | 0.01 | - | 0 | 0.00 | i | 25 | 0.31 | - | 25 | 0.31 | - | 1.28 | ↓46.55 |
| Chlorophyll a 1 | 80 | 24 | 0.30 | - | 1 | 0.01 | - | 0 | 0.00 | i | 25 | 0.31 | - | 25 | 0.31 | - | 1.28 | ↓46.55 |
| Chlorophyll b 0.5 | 80 | 28 | 0.35 | - | 1 | 0.01 | - | 2 | 0.02 | i | 31 | 0.39 | - | 31 | 0.39 | - | 1.58 | ↓32.76 |
| Chlorophyll b 1 | 80 | 33 | 0.41 | - | 1 | 0.01 | - | 1 | 0.01 | i | 35 | 0.44 | - | 35 | 0.44 | - | 1.79 | ↓24.14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocaoğlu Cenkci, S.; Kaya, B. Effects of Chlorophyll a and b in Reducing Genotoxicity of 2-Amino-3,8-dimethylimidazo[4,5-F]quinoxaline (MeIQx). Biology 2022, 11, 602. https://doi.org/10.3390/biology11040602
Kocaoğlu Cenkci S, Kaya B. Effects of Chlorophyll a and b in Reducing Genotoxicity of 2-Amino-3,8-dimethylimidazo[4,5-F]quinoxaline (MeIQx). Biology. 2022; 11(4):602. https://doi.org/10.3390/biology11040602
Chicago/Turabian StyleKocaoğlu Cenkci, Serap, and Bülent Kaya. 2022. "Effects of Chlorophyll a and b in Reducing Genotoxicity of 2-Amino-3,8-dimethylimidazo[4,5-F]quinoxaline (MeIQx)" Biology 11, no. 4: 602. https://doi.org/10.3390/biology11040602
APA StyleKocaoğlu Cenkci, S., & Kaya, B. (2022). Effects of Chlorophyll a and b in Reducing Genotoxicity of 2-Amino-3,8-dimethylimidazo[4,5-F]quinoxaline (MeIQx). Biology, 11(4), 602. https://doi.org/10.3390/biology11040602