
The Effect of Hypothermia and Osmotic Shock on the Electrocardiogram of Adult Zebrafish

 , ,
, ,  ,
,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Strains and Husbandry
2.2. ECG Recording
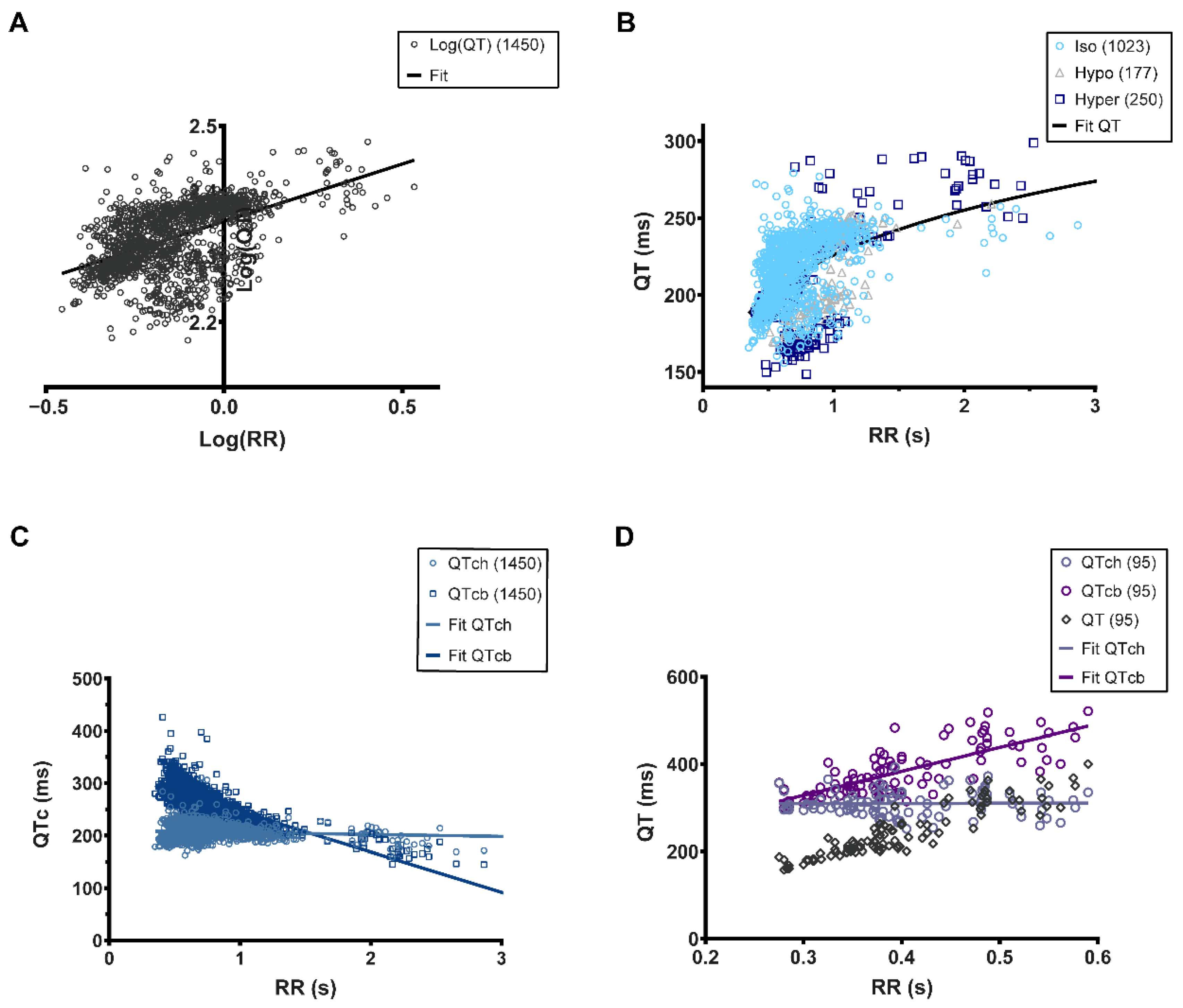
2.3. QT Analysis and Correction
2.4. Solutions and Experimental Protocols
2.5. Statistical Analysis
3. Results
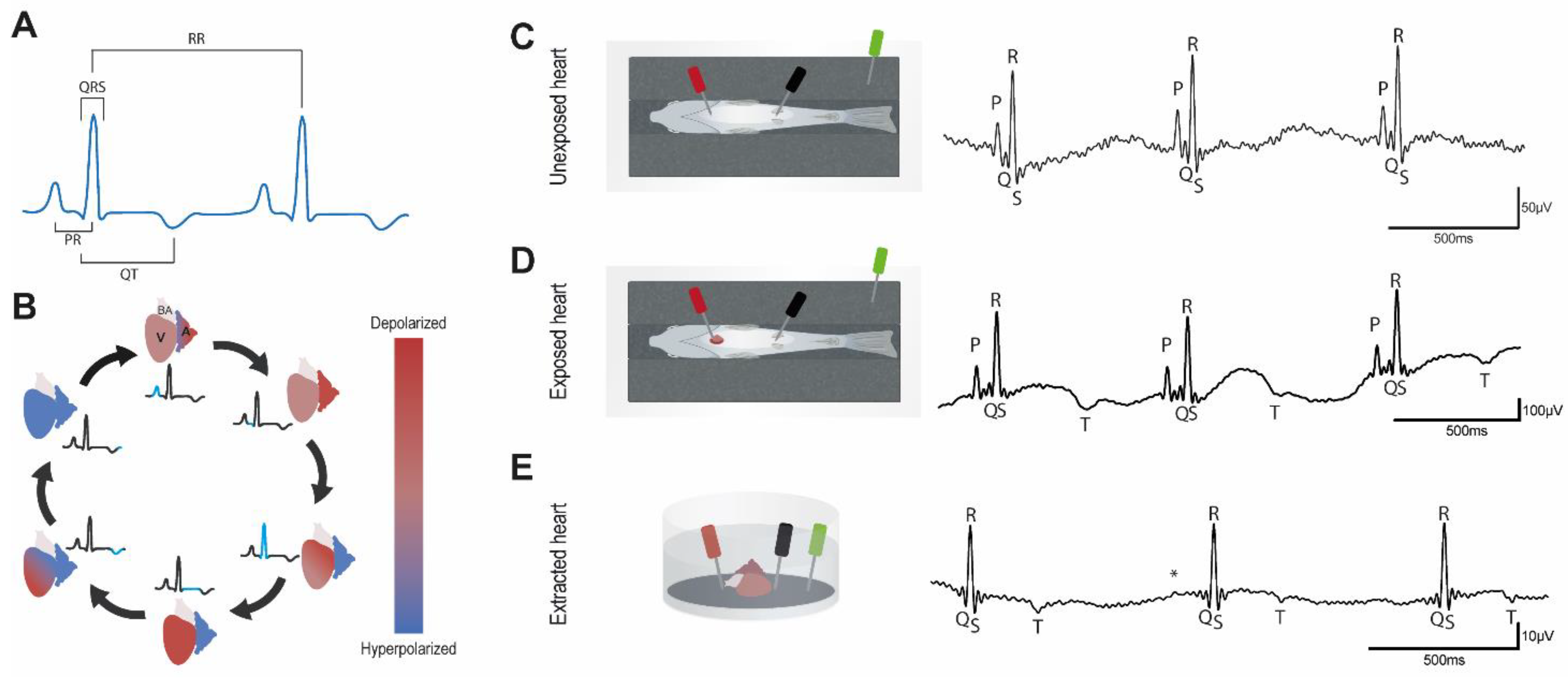
3.1. Basal Characteristics of Adult Zebrafish ECG
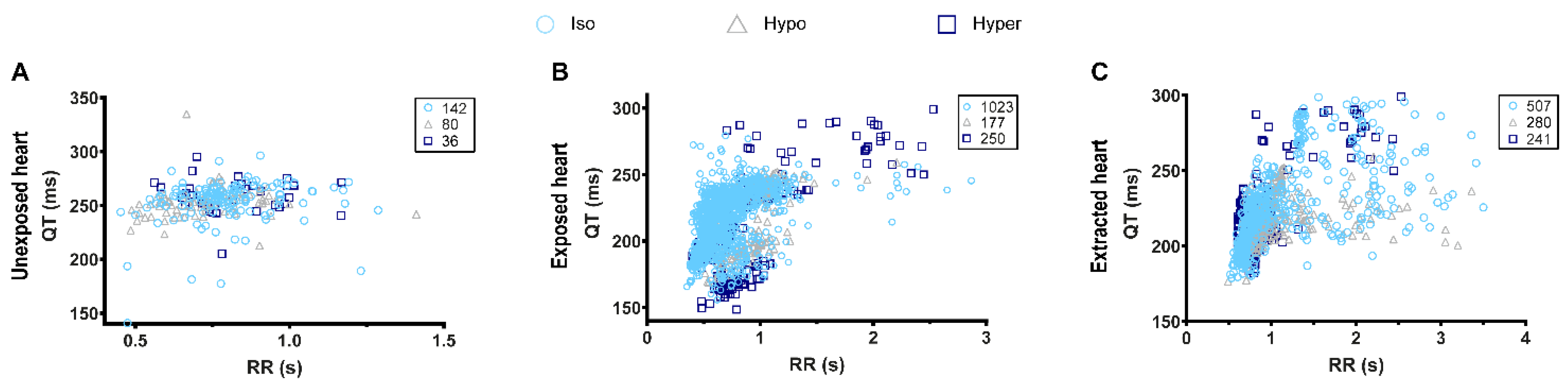
3.2. Effects of Osmotic Shock on Adult Zebrafish ECG Characteristics
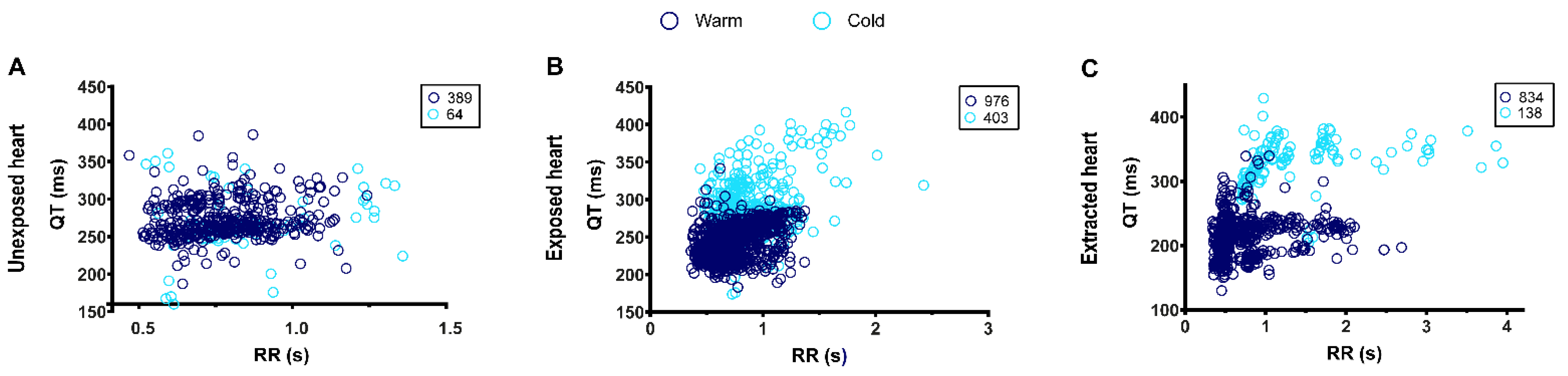
3.3. Effects of Hypothermia on Adult Zebrafish ECG Characteristics
3.4. QT-RR Relationship
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Milan, D.J.; Jones, I.L.; Ellinor, P.T.; MacRae, C.A. In Vivo Recording of Adult Zebrafish Electrocardiogram and Assessment of Drug-Induced QT Prolongation. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, H269–H273. [Google Scholar] [CrossRef] [PubMed]
- Arnaout, R.; Ferrer, T.; Huisken, J.; Spitzer, K.; Stainier, D.Y.R.; Tristani-Firouzi, M.; Chi, N.C. Zebrafish Model for Human Long QT Syndrome. Proc. Natl. Acad. Sci. USA 2007, 104, 11316–11321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaudhari, G.H.; Chennubhotla, K.S.; Chatti, K.; Kulkarni, P. Optimization of the Adult Zebrafish ECG Method for Assessment of Drug-Induced QTc Prolongation. J. Pharmacol. Toxicol. Methods 2013, 67, 115–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.C.; Li, L.; Lam, Y.W.; Siu, C.W.; Cheng, S.H. Improvement of Surface ECG Recording in Adult Zebrafish Reveals That the Value of This Model Exceeds Our Expectation. Sci. Rep. 2016, 6, 25073. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.-H.; Chou, H.-C.; Chen, Y.-F.; Liu, W.; Lee, C.-C.; Liu, L.Y.-M.; Chuang, Y.-J. Development of a Rapid and Economic in Vivo Electrocardiogram Platform for Cardiovascular Drug Assay and Electrophysiology Research in Adult Zebrafish. Sci. Rep. 2018, 8, 15986. [Google Scholar] [CrossRef]
- Echeazarra, L.; Hortigón-Vinagre, M.P.; Casis, O.; Gallego, M. Adult and Developing Zebrafish as Suitable Models for Cardiac Electrophysiology and Pathology in Research and Industry. Front. Physiol. 2020, 11, 607860. [Google Scholar] [CrossRef]
- Nemtsas, P.; Wettwer, E.; Christ, T.; Weidinger, G.; Ravens, U. Adult Zebrafish Heart as a Model for Human Heart? An Electrophysiological Study. J. Mol. Cell. Cardiol. 2010, 48, 161–171. [Google Scholar] [CrossRef]
- Brette, F.; Luxan, G.; Cros, C.; Dixey, H.; Wilson, C.; Shiels, H.A. Characterization of Isolated Ventricular Myocytes from Adult Zebrafish (Danio rerio). Biochem. Biophys. Res. Commun. 2008, 374, 143–146. [Google Scholar] [CrossRef]
- Holzgrefe, H.; Ferber, G.; Champeroux, P.; Gill, M.; Honda, M.; Greiter-Wilke, A.; Baird, T.; Meyer, O.; Saulnier, M. Preclinical QT Safety Assessment: Cross-Species Comparisons and Human Translation from an Industry Consortium. J. Pharmacol. Toxicol. Methods 2014, 69, 61–101. [Google Scholar] [CrossRef] [Green Version]
- Dickinson, D.F. The Normal ECG in Childhood and Adolescence. Heart Br. Card. Soc. 2005, 91, 1626–1630. [Google Scholar] [CrossRef] [Green Version]
- Raslan, A.; Bhardwaj, A. Medical Management of Cerebral Edema. Neurosurg. Focus 2007, 22, E12. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski, W.; Siwicka-Gieroba, D.; Robba, C.; Badenes, R.; Bialy, M.; Iwaniuk, P.; Schlegel, T.T.; Jaroszynski, A. Plasma Hyperosmolality Prolongs QTc Interval and Increases Risk for Atrial Fibrillation in Traumatic Brain Injury Patients. J. Clin. Med. 2020, 9, 1293. [Google Scholar] [CrossRef] [PubMed]
- Dabrowski, W.; Siwicka-Gieroba, D.; Robba, C.; Bielacz, M.; Sołek-Pastuszka, J.; Kotfis, K.; Bohatyrewicz, R.; Jaroszyński, A.; Malbrain, M.L.N.G.; Badenes, R. Potentially Detrimental Effects of Hyperosmolality in Patients Treated for Traumatic Brain Injury. J. Clin. Med. 2021, 10, 4141. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.N.; Prasad, N.; Glancy, J.M. QTc Prolongation during Therapeutic Hypothermia: Are We Giving It the Attention It Deserves? EP Europace 2010, 12, 266–270. [Google Scholar] [CrossRef]
- Rosol, Z.; Miranda, D.F.; Sandoval, Y.; Bart, B.A.; Smith, S.W.; Goldsmith, S.R. The Effect of Targeted Temperature Management on QT and Corrected QT Intervals in Patients with Cardiac Arrest. J. Crit. Care 2017, 39, 182–184. [Google Scholar] [CrossRef]
- Bazett, H.C. An Analysis of the Time-Relations of Electrocardiograms. Heart 1920, 7, 353–370. [Google Scholar] [CrossRef]
- Brönnimann, D.; Djukic, T.; Triet, R.; Dellenbach, C.; Saveljic, I.; Rieger, M.; Rohr, S.; Filipovic, N.; Djonov, V. Pharmacological Modulation of Hemodynamics in Adult Zebrafish In Vivo. PLoS ONE 2016, 11, e0150948. [Google Scholar] [CrossRef] [Green Version]
- Le, T.; Lenning, M.; Clark, I.; Bhimani, I.; Fortunato, J.; Mash, P.; Xu, X.; Cao, H. Acquisition, Processing and Analysis of Electrocardiogram in Awake Zebrafish. IEEE Sens. J. 2019, 19, 4283–4289. [Google Scholar] [CrossRef]
- Song, J.; Qiao, L.; Ji, L.; Ren, B.; Hu, Y.; Zhao, R.; Ren, Z. Toxic Responses of Zebrafish (Danio Rerio) to Thallium and Deltamethrin Characterized in the Electrocardiogram. Chemosphere 2018, 212, 1085–1094. [Google Scholar] [CrossRef]
- Zhao, Y.; James, N.A.; Beshay, A.R.; Chang, E.E.; Lin, A.; Bashar, F.; Wassily, A.; Nguyen, B.; Nguyen, T.P. Adult Zebrafish Ventricular Electrical Gradients as Tissue Mechanisms of ECG Patterns under Baseline vs. Oxidative Stress. Cardiovasc. Res. 2021, 117, 1891–1907. [Google Scholar] [CrossRef]
- Yu, F.; Li, R.; Parks, E.; Takabe, W.; Hsiai, T.K. Electrocardiogram Signals to Assess Zebrafish Heart Regeneration: Implication of Long QT Intervals. Ann. Biomed. Eng. 2010, 38, 2346–2357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chablais, F.; Veit, J.; Rainer, G.; Jaźwińska, A. The Zebrafish Heart Regenerates after Cryoinjury-Induced Myocardial Infarction. BMC Dev. Biol. 2011, 11, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Cao, H.; Kang, B.J.; Jen, N.; Yu, F.; Lee, C.-A.; Fei, P.; Park, J.; Bohlool, S.; Lash-Rosenberg, L.; et al. Hemodynamics and Ventricular Function in a Zebrafish Model of Injury and Repair. Zebrafish 2014, 11, 447–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Tai, J.; Park, J.; Tai, Y.-C. Flexible MEA for Adult Zebrafish ECG Recording Covering Both Ventricle and Atrium. In Proceedings of the 2014 IEEE 27th International Conference on Micro Electro Mechanical Systems (MEMS), San Francisco, CA, USA, 26–30 January 2014; pp. 841–844. [Google Scholar]
- Lenning, M.; Fortunato, J.; Le, T.; Clark, I.; Sherpa, A.; Yi, S.; Hofsteen, P.; Thamilarasu, G.; Yang, J.; Xu, X.; et al. Real-Time Monitoring and Analysis of Zebrafish Electrocardiogram with Anomaly Detection. Sensors 2017, 18, 61. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Yun, M.; Nguyen, S.A.; Tran, M.; Nguyen, T.P. In Vivo Surface Electrocardiography for Adult Zebrafish. J. Vis. Exp. JoVE 2019, 18, 61. [Google Scholar] [CrossRef]
- Sun, P.; Zhang, Y.; Yu, F.; Parks, E.; Lyman, A.; Wu, Q.; Ai, L.; Hu, C.-H.; Zhou, Q.; Shung, K.; et al. Micro-Electrocardiograms to Study Post-Ventricular Amputation of Zebrafish Heart. Ann. Biomed. Eng. 2009, 37, 890–901. [Google Scholar] [CrossRef]
- Zhang, X.; Beebe, T.; Jen, N.; Lee, C.-A.; Tai, Y.; Hsiai, T.K. Flexible and Waterproof Micro-Sensors to Uncover Zebrafish Circadian Rhythms: The next Generation of Cardiac Monitoring for Drug Screening. Biosens. Bioelectron. 2015, 71, 150–157. [Google Scholar] [CrossRef] [Green Version]
- Yu, F.; Zhao, Y.; Gu, J.; Quigley, K.L.; Chi, N.C.; Tai, Y.-C.; Hsiai, T.K. Flexible Microelectrode Arrays to Interface Epicardial Electrical Signals with Intracardial Calcium Transients in Zebrafish Hearts. Biomed. Microdevices 2012, 14, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Cao, H.; Yu, F.; Zhao, Y.; Zhang, X.; Tai, J.; Lee, J.; Darehzereshki, A.; Bersohn, M.; Lien, C.-L.; Chi, N.C.; et al. Wearable Multi-Channel Microelectrode Membranes for Elucidating Electrophysiological Phenotypes of Injured Myocardium. Integr. Biol. Quant. Biosci. Nano Macro 2014, 6, 789–795. [Google Scholar] [CrossRef] [Green Version]
- Meder, B.; Scholz, E.P.; Hassel, D.; Wolff, C.; Just, S.; Berger, I.M.; Patzel, E.; Karle, C.; Katus, H.A.; Rottbauer, W. Reconstitution of Defective Protein Trafficking Rescues Long-QT Syndrome in Zebrafish. Biochem. Biophys. Res. Commun. 2011, 408, 218–224. [Google Scholar] [CrossRef]
- Tsai, C.-T.; Wu, C.-K.; Chiang, F.-T.; Tseng, C.-D.; Lee, J.-K.; Yu, C.-C.; Wang, Y.-C.; Lai, L.-P.; Lin, J.-L.; Hwang, J.-J. In-Vitro Recording of Adult Zebrafish Heart Electrocardiogram—A Platform for Pharmacological Testing. Clin. Chim. Acta Int. J. Clin. Chem. 2011, 412, 1963–1967. [Google Scholar] [CrossRef] [PubMed]
- Stoyek, M.R.; Quinn, T.A.; Croll, R.P.; Smith, F.M. Zebrafish Heart as a Model to Study the Integrative Autonomic Control of Pacemaker Function. Am. J. Physiol. Heart Circ. Physiol. 2016, 311, H676–H688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwerte, T.; Prem, C.; Mairösl, A.; Pelster, B. Development of the Sympatho-Vagal Balance in the Cardiovascular System in Zebrafish (Danio rerio) Characterized by Power Spectrum and Classical Signal Analysis. J. Exp. Biol. 2006, 209, 1093–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stoyek, M.R.; Hortells, L.; Quinn, T.A. From Mice to Mainframes: Experimental Models for Investigation of the Intracardiac Nervous System. J. Cardiovasc. Dev. Dis. 2021, 8, 149. [Google Scholar] [CrossRef] [PubMed]
- Champeroux, P.; Le Guennec, J.Y.; Jude, S.; Laigot, C.; Maurin, A.; Sola, M.L.; Fowler, J.S.L.; Richard, S.; Thireau, J. The High Frequency Relationship: Implications for Torsadogenic HERG Blockers. Br. J. Pharmacol. 2016, 173, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Champéroux, P.; Fesler, P.; Judé, S.; Richard, S.; Le Guennec, J.-Y.; Thireau, J. High-Frequency Autonomic Modulation: A New Model for Analysis of Autonomic Cardiac Control. Br. J. Pharmacol. 2018, 175, 3131–3143. [Google Scholar] [CrossRef]
- Jopling, C.; Sleep, E.; Raya, M.; Martí, M.; Raya, A.; Izpisúa Belmonte, J.C. Zebrafish Heart Regeneration Occurs by Cardiomyocyte Dedifferentiation and Proliferation. Nature 2010, 464, 606–609. [Google Scholar] [CrossRef]
- Guzzolino, E.; Pellegrino, M.; Ahuja, N.; Garrity, D.; D’Aurizio, R.; Groth, M.; Baumgart, M.; Hatcher, C.J.; Mercatanti, A.; Evangelista, M.; et al. MiR-182-5p Is an Evolutionarily Conserved Tbx5 Effector That Impacts Cardiac Development and Electrical Activity in Zebrafish. Cell. Mol. Life Sci. CMLS 2020, 77, 3215–3229. [Google Scholar] [CrossRef]
- Boisen, A.M.Z.; Amstrup, J.; Novak, I.; Grosell, M. Sodium and Chloride Transport in Soft Water and Hard Water Acclimated Zebrafish (Danio rerio). Biochim. Biophys. Acta 2003, 1618, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Kennard, A.S.; Theriot, J.A. Osmolarity-Independent Electrical Cues Guide Rapid Response to Injury in Zebrafish Epidermis. eLife 2020, 9, e62386. [Google Scholar] [CrossRef]
- Jandu, S.; Sefa, N.; Sawyer, K.N.; Swor, R. Electrocardiographic Changes in Patients Undergoing Targeted Temperature Management. J. Am. Coll. Emerg. Physicians Open 2020, 1, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Hassel, D.; Scholz, E.P.; Trano, N.; Friedrich, O.; Just, S.; Meder, B.; Weiss, D.L.; Zitron, E.; Marquart, S.; Vogel, B.; et al. Deficient Zebrafish Ether-à-Go-Go-Related Gene Channel Gating Causes Short-QT Syndrome in Zebrafish Reggae Mutants. Circulation 2008, 117, 866–875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mersereau, E.J.; Poitra, S.L.; Espinoza, A.; Crossley, D.A.; Darland, T. The Effects of Cocaine on Heart Rate and Electrocardiogram in Zebrafish (Danio rerio). Comp. Biochem. Physiol. Toxicol. Pharmacol. CBP 2015, 172–173, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holden, L.A.; Brown, K.H. Baseline MRNA Expression Differs Widely between Common Laboratory Strains of Zebrafish. Sci. Rep. 2018, 8, 4780. [Google Scholar] [CrossRef] [PubMed]
- Duong, T.; Rose, R.; Blazeski, A.; Fine, N.; Woods, C.E.; Thole, J.F.; Sotoodehnia, N.; Soliman, E.Z.; Tung, L.; McCallion, A.S.; et al. Development and Optimization of an In Vivo Electrocardiogram Recording Method and Analysis Program for Adult Zebrafish. Dis. Model. Mech. 2021, 14, dmm048827. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Configuration | PR (ms) K | QRS (ms) K | QT (ms) K | RR (ms) K | HR (bpm) A |
|---|---|---|---|---|---|
| Unexposed Heart | 44.79 (1.90) (20) | 25.93 (1.02) (20) | 273.4 (7.65) (8) | 766.8 (30.92) (20) | 80.27 (3.07) (20) |
| Exposed Heart | 41.44 (2.22) (17) | 31.34 (1.54) * (20) | 234.8 (6.29) (16) | 699.9 (32.99) (20) | 86.46 (4.40) (20) |
| Extracted Heart | 49.87 (6.52) (5) | 34.09 (4.94) (17) | 213.4 (17.60) ** (13) | 910.1 (85.80) (18) | 74.68 (6.24) (18) |
| Configuration | Positive T Wave | Negative T Wave | Undetectable |
|---|---|---|---|
| Unexposed Heart | 0% (0/20) | 40% (8/20) | 60% (12/20) |
| Exposed Heart | 25% (5/20) | 55% (11/20) | 20% (4/20) |
| Extracted Heart | 28% (5/18) | 44% (8/18) | 28% (5/18) |
| Unexposed Heart | PR (ms) K | QRS (ms) A | QT (ms) NC | RR (ms) K | HR (bpm) A |
|---|---|---|---|---|---|
| Tyrode | 46.15 (3.34)(10) | 26.65 (1.24) (10) | 271.5 (9.01) (3) | 799.5 (41.08) (10) | 76.82 (4.01) (10) |
| Isosmotic | 57.01 (13.14) (10) | 25.86 (1.45) (10) | 265.1 (8.38) (3) | 756.5 (44.95) (10) | 81.34 (3.89) (10) |
| Hyposmotic | 47.08 (2.33) (9) | 26.11 (1.47) (10) | 266.8 (4.24) (3) | 727.7 (46.7) (10) | 84.85 (4.25) (10) |
| Wash isosmotic | 46.97 (1.95) (7) | 24.86 (1.79) (8) | NC (1) | 743.3 (38.04) (8) | 82.27 (4.36) (8) |
| Hyperosmotic | 46.59 (2.44) (8) | 25.95 (1.22) (10) | NC (2) | 766.2 (44.61) (10) | 80.50 (4.26) (10) |
| Exposed heart | PR (ms) A | QRS (ms) A | QT (ms) A | RR (ms) A | HR (bpm) A |
| Tyrode | 43.78 (4.04) (10) | 33.80 (1.46) (10) | 235.0 (9.99) (10) | 705.4 (45.32) (10) | 88.48 (5.86) (10) |
| Isosmotic | 42.13 (2.02) (9) | 31.39 (1.47) (10) | 241.3 (11.73) (8) | 733.2 (149.0) (10) | 85.3 (6.08) (10) |
| Hyposmotic | 41.07 (2.02) (9) | 29.27 (1.69) (10) | 244.9 (12.9) (7) | 716.6 (46.9) (10) | 87.43 (6.37) (10) |
| Wash isosmotic | 41.36 (1.98) (9) | 29.39 (1.74) (10) | 243.9 (14.02) (6) | 710.4 (51.48) (10) | 88.86 (6.92) (10) |
| Hyperosmotic | 40.70 (1.83) (9) | 31.07 (1.55) (10) | 245.2 (14.48) (5) | 729.6 (48.69) (10) | 85.88 (6.18) (10) |
| Extracted heart | PR (ms) NC | QRS (ms) K | QT (ms) A | RR (ms) A | HR (bpm) A |
| Tyrode | NC (0) | 42.82 (8.06) (9) | 233.9 (19.11) (6) | 954.0 (102.5) (10) | 69.79 (7.80) (10) |
| Isosmotic | NC (0) | 33.23 (5.01) (9) | 232.2 (6.18) (5) | 1239.0 (240.0) (10) | 67.24 (12.14) (10) |
| Hyposmotic | NC (0) | 32.00 (4.84) (9) | 224.4 (6.71) (7) | 1201.0 (202.6) (10) | 63.27 (9.95) (10) |
| Wash isosmotic | NC (0) | 31.99 (5.05) (9) | 215.7 (6.17) (7) | 980.7 (180.7) (10) | 77.25 (10.31) (10) |
| Hyperosmotic | NC (0) | 31.18 (4.14) (8) | 227.7 (8.44) (7) | 1132.0 (186.0) (10) | 64.46 (8.23) (10) |
| Unexposed Heart | PR (ms) A | QRS (ms) A | QT (ms) K | RR (ms) A | HR (bpm) A |
|---|---|---|---|---|---|
| Warm (control) | 43.42 (1.90) (10) | 25.22 (1.66) (10) | 273.4 (7.65) (5) | 734 (45.94) (10) | 83.72 (4.60) (10) |
| Cold | 78.47 (6.87) **** (10) | 37.59 (2.71) ** (10) | 252.2 (43.08) (2) | 1401 (196.9) ** (10) | 50.86 (7.30) ** (10) |
| Warm (washout) | 46.95 (2.23) (8) | 25.33 (2.11) (9) | 272.5 (26.66) (3) | 921.4 (81.02) (10) | 69.05 (5.84) (10) |
| Exposed heart | PR (ms) A | QRS (ms) A | QT (ms) A | RR (ms) A | HR (bpm) A |
| Warm (control) | 38.48 (1.91) (8) | 28.88 (2.55) (10) | 234.5 (4.07) (6) | 694.4 (50.34) (10) | 84.44 (6.80) (10) |
| Cold | 49.78 (2.96) ** (9) | 29.96 (2.35) (10) | 292.6 (11.86) ** (9) | 956.6 (76.04) * (10) | 65.36 (4.43) (10) |
| Warm (washout) | 41.32 (1.94) (9) | 25.99 (2.96) (10) | 244.5 (6.57) (8) | 762.9 (74.31) (10) | 83.74 (76) (10) |
| Extracted heart | PR (ms) A | QRS (ms) K | QT (ms) K | RR (ms) K | HR (bpm) A |
| Warm (control) | 49.87 (6.52) (5) | 24.26 (3) (8) | 195.8 (27.90) (7) | 855.1 (150.3) (8) | 80.79 (10.29) (8) |
| Cold | 56.23 (6.65) (4) | 38.80 (3.92) (8) | 341.8 (9.05) * (5) | 2038 (462.5) * (8) | 39.28 (6.97) * (8) |
| Warm (washout) | 41.42 (3.45) (3) | 34.52 (11.95) (8) | 220.5 (11.62) (7) | 846 (116.9) (8) | 77.68 (10.08) (8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arel, E.; Rolland, L.; Thireau, J.; Torrente, A.G.; Bechard, E.; Bride, J.; Jopling, C.; Demion, M.; Le Guennec, J.-Y. The Effect of Hypothermia and Osmotic Shock on the Electrocardiogram of Adult Zebrafish. Biology 2022, 11, 603. https://doi.org/10.3390/biology11040603
Arel E, Rolland L, Thireau J, Torrente AG, Bechard E, Bride J, Jopling C, Demion M, Le Guennec J-Y. The Effect of Hypothermia and Osmotic Shock on the Electrocardiogram of Adult Zebrafish. Biology. 2022; 11(4):603. https://doi.org/10.3390/biology11040603
Chicago/Turabian StyleArel, Elodie, Laura Rolland, Jérôme Thireau, Angelo Giovanni Torrente, Emilie Bechard, Jamie Bride, Chris Jopling, Marie Demion, and Jean-Yves Le Guennec. 2022. "The Effect of Hypothermia and Osmotic Shock on the Electrocardiogram of Adult Zebrafish" Biology 11, no. 4: 603. https://doi.org/10.3390/biology11040603
APA StyleArel, E., Rolland, L., Thireau, J., Torrente, A. G., Bechard, E., Bride, J., Jopling, C., Demion, M., & Le Guennec, J. -Y. (2022). The Effect of Hypothermia and Osmotic Shock on the Electrocardiogram of Adult Zebrafish. Biology, 11(4), 603. https://doi.org/10.3390/biology11040603