
Environmental Factors and the Risk of Developing Type 1 Diabetes—Old Disease and New Data

 ,
,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
1.1. Type 1 Diabetes Mellitus (T1DM)—Etiopathogenesis
1.2. Genetic Predisposition as an Etiological Factor of T1DM
1.3. The Accelerator Hypothesis, the β-Cell “Overload” and the Risk of Developing Diabetes
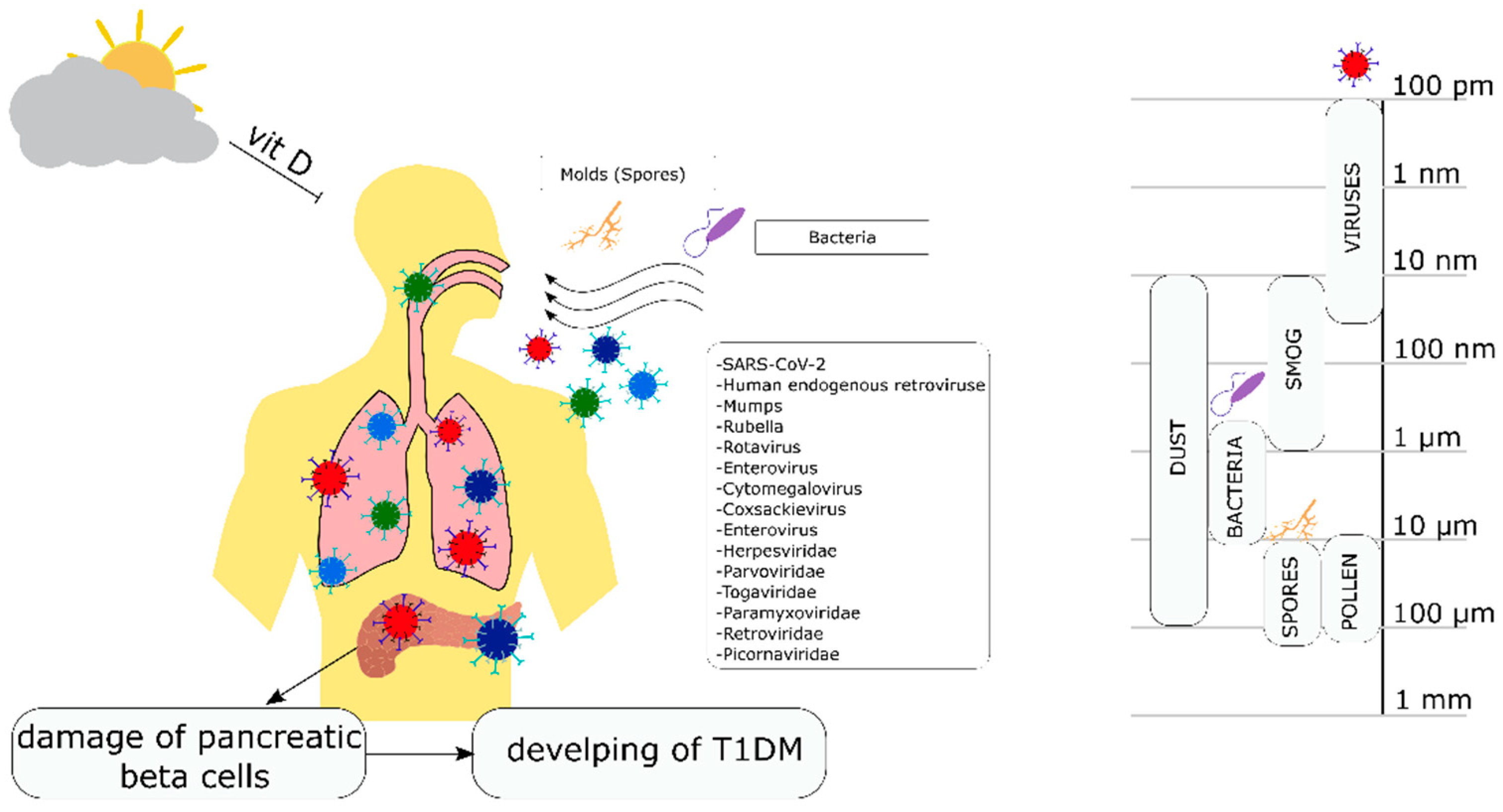
2. Environmental Factors
Climatic Conditions, Lack of Vitamin D Deficiency, Feeding with Cow’s Milk, and the Risk of Developing Type 1 Diabetes
3. Viral Infections and the Risk of Developing T1DM
4. The Effect of Bacteria and Yeast-Like Fungi on the Risk of Developing T1DM Diabetes
Limitations and Recommendations
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chobot, A.; Polanska, J.; Deja, G.; Jarosz-Chobot, P. Incidence of type 1 diabetes among Polish children ages 0–14 years from 1989–2012. Acta Diabetol. 2015, 52, 483–488. [Google Scholar] [CrossRef] [PubMed]
- Patterson, C.C.; Dahlquist, G.G.; Gyürüs, E.; Green, A.; Soltész, G. Incidence trends for childhood type 1 diabetes in Europe during 1989–2003 and predicted new cases 2005–20: A multicentre prospective registration study. Lancet 2009, 373, 2027–2033. [Google Scholar] [CrossRef]
- Altobelli, E.; Petrocelli, R.; Verrotti, A.; Chiarelli, F.; Marziliano, C. Genetic and environmental factors affect the onset of type 1 diabetes mellitus. Pediatr. Diabetes 2016, 17, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Pang, H.; Luo, S.; Huang, G.; Xia, Y.; Xie, Z.; Zhou, Z. Advances in Knowledge of Candidate Genes Acting at the Beta-Cell Level in the Pathogenesis of T1DM. Front. Endocrinol. 2020, 11, 119. [Google Scholar] [CrossRef] [PubMed]
- Michalska, M.; Zorena, K.; Wąż, P.; Bartoszewicz, M.; Brandt-Varma, A.; Ślęzak, D.; Robakowska, M. Gaseous Pollutants and Particulate Matter (PM) in Ambient Air and the Number of New Cases of Type 1 Diabetes in Children and Adolescents in the Pomeranian Voivodeship, Poland. Biomed. Res. Int. 2020, 2020, 1648264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedi, S.; Richardson, T.M.; Jia, B.; Saab, H.; Brinkman, F.S.L.; Westley, M. Similarities between bacterial GAD and human GAD65: Implications in gut mediated autoimmune type 1 diabetes. PLoS ONE 2022, 17, e0261103. [Google Scholar] [CrossRef]
- McGlacken-Byrne, S.M.; Drew, S.E.V.; Turner, K.; Peters, C.; Amin, R. The SARS-CoV-2 pandemic is associated with increased severity of presentation of childhood onset type 1 diabetes mellitus: A multi-centre study of the first COVID-19 wave. Diabet Med. 2021, 38, e14640. [Google Scholar] [CrossRef]
- Rewers, A.; Dong, F.; Slover, R.H.; Klingensmith, G.J.; Rewers, M. Incidence of diabetic ketoacidosis at diagnosis of type 1 diabetes in Colorado youth, 1998–2012. JAMA 2015, 313, 1570–1572. [Google Scholar] [CrossRef] [Green Version]
- Strollo, R.; Vinci, C.; Napoli, N.; Pozzilli, P.; Ludvigsson, J.; Nissim, A. Antibodies to post-translationally modified insulin as a novel biomarker for prediction of type 1 diabetes in children. Diabetologia 2017, 60, 1467–1474. [Google Scholar] [CrossRef]
- Pociot, F.; Lernmark, Å. Genetic risk factors for type 1 diabetes. Lancet 2016, 387, 2331–2339. [Google Scholar] [CrossRef]
- Cerna, M. Epigenetic Regulation in Etiology of Type 1 Diabetes Mellitus. Int. J. Mol. Sci. 2020, 21, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lammi, N.; Karvonen, M.; Tuomilehto, J. Do microbes have a causal role in type 1 diabetes? Med. Sci. Monit. 2005, 11, 63–69. [Google Scholar]
- Zipris, D. Epidemiology of type 1 diabetes and what animal models teach us about the role of viruses in disease mechanisms. Clin. Immunol. 2009, 131, 11–23. [Google Scholar] [CrossRef] [PubMed]
- La Marca, V.; Fierabracci, A. Insights into the Diagnostic Potential of Extracellular Vesicles and Their miRNA Signature from Liquid Biopsy as Early Biomarkers of Diabetic Micro/Macrovascular Complications. Int. J. Mol. Sci. 2017, 18, 1974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Liang, S.; Zhang, C. NK Cells in Autoimmune Diseases: Protective or Pathogenic? Front. Immunol. 2021, 12, 701. [Google Scholar] [CrossRef] [PubMed]
- Jaberi-Douraki, M.; Schnell, S.; Pietropaolo, M.; Khadra, A. Unraveling the contribution of pancreatic beta-cell suicide in autoimmune type 1 diabetes. J. Theor. Biol. 2015, 375, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Pugliese, A. Insulitis in the pathogenesis of type 1 diabetes. Pediatr. Diabetes 2016, 17, 31–36. [Google Scholar] [CrossRef]
- Zorena, K.; Kula, M.; Malinowska, E.; Raczyńska, D.; Myśliwiec, M.; Raczyńska, K. Threshold serum concentrations of tumour necrosis factor alpha (TNFα) as a potential marker of the presence of microangiopathy in children and adolescents with type 1 diabetes mellitus (T1DM). Hum Immunol. 2013, 74, 75–81. [Google Scholar] [CrossRef]
- Zorena, K.; Myśliwska, J.; Myśliwiec, M.; Balcerska, A.; Lipowski, P.; Raczyńska, K. Interleukin-12 and Tumour Necrosis Factor-α Equilibrium is a Prerequisite for Clinical Course Free from Late Complications in Children with Type 1 Diabetes Mellitus. Scand. J. Immunol. 2008, 67, 204–208. [Google Scholar] [CrossRef]
- Antonelli, A.; Fallahi, P.; Ferrari, S.M.; Pupilli, C.; D’Annunzio, G.; Lorini, R.; Vanelli, M.; Ferrannini, E. Serum Th1 (CXCL10) and Th2 (CCL2) chemokine levels in children with newly diagnosed Type 1 diabetes: A longitudinal study. Diabet. Med. 2008, 25, 1349–1353. [Google Scholar] [CrossRef]
- Ayabe, T.; Fukami, M.; Ogata, T.; Kawamura, T.; Urakami, T.; Kikuchi, N.; Yokota, I.; Ihara, K.; Takemoto, K.; Mukai, T.; et al. Variants associated with autoimmune Type 1 diabetes in Japanese children: Implications for age-specific effects of cis-regulatory haplotypes at 17q12-q21. Diabet. Med. 2016, 33, 1717–1722. [Google Scholar] [CrossRef] [PubMed]
- Fullerton, B.; Siebenhofer, A.; Jeitler, K.; Horvath, K.; Semlitsch, T.; Berghold, A.; Plank, J.; Pieber, T.R.; Gerlach, F.M. Short-acting insulin analogues versus regular human insulin for adults with type 1 diabetes mellitus. Cochrane Database Syst. Rev. 2016, 6, CD012161. [Google Scholar] [CrossRef] [PubMed]
- Rewers, M.; Ludvigsson, J. Environmental risk factors for type 1 diabetes. Lancet 2016, 387, 2340–2348. [Google Scholar] [CrossRef] [Green Version]
- Cieślak, M.; Cieślak, M. Role of purinergic signalling and proinflammatory cytokines in diabetes. Clin. Diabetol. 2017, 6, 90–100. [Google Scholar] [CrossRef]
- Principi, N.; Berioli, M.G.; Bianchini, S.; Esposito, S. Type 1 diabetes and viral infections: What is the relationship? J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2017, 96, 26–31. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.; Sonkar, S.K.; Sonkar, G.K.; Mahdi, A.A. Diabetic kidney disease: A systematic review on the role of epigenetics as diagnostic and prognostic marker. Diabetes. Metab. Res. Rev. 2019, 35, e3155. [Google Scholar] [CrossRef] [PubMed]
- Hyttinen, V.; Kaprio, J.; Kinnunen, L.; Koskenvuo, M.; Tuomilehto, J. Genetic liability of type 1 diabetes and the onset age among 22,650 young Finnish twin pairs: A nationwide follow-up study. Diabetes 2003, 52, 1052–1055. [Google Scholar] [CrossRef] [Green Version]
- Paschou, S.; Papadopoulou-Marketou, N.; Chrousos, G.; Kanaka-Gantenbein, C. On type 1 diabetes mellitus pathogenesis. Endocr. Connect. 2018, 7, R38–R46. [Google Scholar] [CrossRef]
- Siewko, K.; Popławska-Kita, A.; Telejko, B.; Maciulewski, R.; Zielińska, A.; Nikołajuk, A.; Górska, M.; Szelachowska, M. Prognostic markers for the development of type 1 diabetes in first-degree relatives of diabetic patients. Endokrynol. Pol. 2014, 65, 176–180. [Google Scholar] [CrossRef] [Green Version]
- Nerup, J.; Platz, P.; Andersen, O.O.; Christy, M.; Lyngsoe, J.; Poulsen, J.E.; Ryder, L.P.; Nielsen, L.S.; Thomsen, M.; Svejgaard, A. HL-A antigens and diabetes mellitus. Lancet 1974, 2, 864–866. [Google Scholar] [CrossRef]
- Farina, F.; Picascia, S.; Pisapia, L.; Barba, P.; Vitale, S.; Franzese, A.; Mozzillo, E.; Gianfrani, C.; Del Pozzo, G.G. HLA-DQA1 and HLA-DQB1 Alleles, Conferring Susceptibility to Celiac Disease and Type 1 Diabetes, Are More Expressed Than Non-Predisposing Alleles and Are Coordinately Regulated. Cells 2019, 8, 751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, D.C.; Porto, L.C.; Oliveira, R.V.; Secco, D.; Hanhoerderster, L.; Pizarro, M.H.; Barros, B.S.V.; Mello, L.G.N.; Muniz, L.H.; Silva, D.A.; et al. HLA class II genotyping of admixed Brazilian patients with type 1 diabetes according to self-reported color/race in a nationwide study. Sci. Rep. 2020, 10, 6628. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, H.; Noso, S.; Babaya, N.; Kawabata, Y. Genetics and pathogenesis of type 1 diabetes: Prospects for prevention and intervention. J. Diabetes Investig. 2011, 2, 415–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugliese, A. Genetics of type 1 diabetes. Endocrinol. Metab. Clin. N. Am. 2004, 33, 1–16. [Google Scholar] [CrossRef]
- Antosik, K.; Gnyś, P.; Jarosz-Chobot, P.; Myśliwiec, M.; Szadkowska, A.; Małecki, M.; Młynarski, W.; Borowiec, M. An analysis of the sequence of the BAD gene among patients with maturity-onset diabetes of the young (MODY). J. Pediatr. Endocrinol. Metab. 2017, 30, 97–100. [Google Scholar] [CrossRef]
- Yalcıntepe, S.; Ozguc Comlek, F.; Gurkan, H.; Demir, S.; Atlı, E.I.; Atlı, E.; Eker, D.; Tutunculer Kokenli, F. The Application of Next Generation Sequencing Maturity Onset Diabetes of the Young Gene Panel in Turkish Patients from Trakya Region. J. Clin. Res. Pediatr. Endocrinol. 2021, 13, 320–331. [Google Scholar] [CrossRef]
- Ivanoshchuk, D.E.; Shakhtshneider, E.V.; Rymar, O.D.; Ovsyannikova, A.K.; Mikhailova, S.V.; Fishman, V.S.; Valeev, E.S.; Orlov, P.S.; Voevoda, M.I. The Mutation Spectrum of Maturity Onset Diabetes of the Young (MODY)-Associated Genes among Western Siberia Patients. J. Pers. Med. 2021, 11, 57. [Google Scholar] [CrossRef]
- Ilonen, J.; Lempainen, J.; Veijola, R. The heterogeneous pathogenesis of type 1 diabetes mellitus. Nat. Rev. Endocrinol. 2019, 15, 635–650. [Google Scholar] [CrossRef]
- Michalska, M.; Bartoszewicz, M.; Wąż, P.; Korzeniowska, K.; Myśliwiec, M.; Zorena, K. The relation between concentration of particulate matter PM10 and the number of new cases of type 1 diabetes mellitus in children and adolescents in the Pomeranian Voivodeship. Med. Srod. 2018, 21, 25–27. [Google Scholar] [CrossRef]
- Kowalewska, B.; Zorena, K.; Szmigiero-Kawko, M.; Wąż, P.; Myśliwiec, M. High Interleukin-12 Levels May Prevent an Increase in the Amount of Fungi in the Gastrointestinal Tract during the First Years of Diabetes Mellitus Type 1. Dis. Markers 2016, 2016, 4685976. [Google Scholar] [CrossRef] [Green Version]
- Di Ciaula, A.; Portincasa, P. Relationships between emissions of toxic airborne molecules and type 1 diabetes incidence in children: An ecologic study. World J. Diabetes 2021, 12, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Michalska, M.; Kurpas, M.; Zorena, K.; Wąż, P.; Marks, R. Mold and Yeast-Like Fungi in the Seaside Air of the Gulf of Gdańsk (Southern Baltic) after an Emergency Disposal of Raw Sewage. J. Fungi 2021, 7, 219. [Google Scholar] [CrossRef] [PubMed]
- Hugerth, L.W.; Larsson, J.; Alneberg, J.; Lindh, M.V.; Legrand, C.; Pinhassi, J.; Andersson, A.F. Metagenome-assembled genomes uncover a global brackish microbiome. Genome Biol. 2015, 16, 279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogensen, T.H.; Paludan, S.R. Molecular pathways in virus-induced cytokine production. Microbiol. Mol. Biol. Rev. 2001, 65, 131–150. [Google Scholar] [CrossRef] [Green Version]
- Cheon, C.K. Understanding of type 1 diabetes mellitus: What we know and where we go. Korean J. Pediatr. 2018, 61, 307–314. [Google Scholar] [CrossRef] [Green Version]
- Myśliwiec, M.; Balcerska, A.; Zorena, K.; Jędrzejczyk, A.; Malinowska, E.; Myśliwska, J. Increasing incidence of diabetes mellitus type 1 in children: The role of environmental factors. Pol. J. Environ. Stud. 2007, 16, 109–112. Available online: https://www.pjoes.com/pdf-87966-21825?filename=Increasing%20Incidence%20of.pdf (accessed on 26 March 2022).
- Ibrahim Saber, A.-Z.A.-B.; Mohammed, A.H. The role of Human Cytomegalovirus and Epstein-Barr virus in type 1 Diabetes Mellitus. Ann. Trop Public Health 2019, 22, S267. [Google Scholar] [CrossRef]
- Murdaca, G.; Greco, M.; Borro, M.; Gangemi, S. Hygiene hypothesis and autoimmune diseases: A narrative review of clinical evidences and mechanisms. Autoimmun. Rev. 2021, 20, 102845. [Google Scholar] [CrossRef]
- Kibirige, M.; Metcalf, B.; Renuka, R.; Wilkin, T.J. Testing the accelerator hypothesis: The relationship between body mass and age at diagnosis of type 1 diabetes. Diabetes Care 2003, 26, 2865–2870. [Google Scholar] [CrossRef] [Green Version]
- Leclair, E.; de Kerdanet, M.; Riddell, M.; Heyman, E. Type 1 Diabetes and Physical Activity in Children and Adolescents. J. Diabetes Metab. 2013, 1, S10. [Google Scholar] [CrossRef] [Green Version]
- Fourlanos, S.; Harrison, L.C.; Colman, P.G. The accelerator hypothesis and increasing incidence of type 1 diabetes. Curr. Opin. Endocrinol. Diabetes Obes. 2008, 15, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Dahlquist, G. Can we slow the rising incidence of childhood-onset autoimmune diabetes? The overload hypothesis. Diabetologia 2006, 49, 20–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secher, A.L.; Parellada, C.B.; Ringholm, L.; Asbjörnsdóttir, B.; Damm, P.; Mathiesen, E.R. Higher gestational weight gain is associated with increasing offspring birth weight independent of maternal glycemic control in women with type 1 diabetes. Diabetes Care 2014, 37, 2677–2684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasyl-Nawrot, B.; Wójcik, M.; Nazim, J.; Skupień, J.; Starzyk, J.B. Increased Incidence of Type 1 Diabetes in Children and No Change in the Age of Diagnosis and BMI-SDS at the Onset—Is the Accelerator Hypothesis not Working? J. Clin. Res. Pediatr. Endocrinol. 2020, 12, 281–286. [Google Scholar] [CrossRef]
- Kuchlbauer, V.; Vogel, M.; Gausche, R.; Kapellen, T.; Rothe, U.; Vogel, C.; Pfäffle, R.; Kiess, W. High birth weights but not excessive weight gain prior to manifestation are related to earlier onset of diabetes in childhood: ‘accelerator hypothesis’ revisited. Pediatr. Diabetes 2014, 15, 428–435. [Google Scholar] [CrossRef]
- Waernbaum, I.; Dahlquist, G. Low mean temperature rather than few sunshine hours are associated with an increased incidence of type 1 diabetes in children. Eur. J. Epidemiol. 2016, 31, 61–65. [Google Scholar] [CrossRef]
- Takiishi, T.; Gysemans, C.; Bouillon, R.; Mathieu, C. Vitamin D and diabetes. Rheum. Dis. Clin. N. Am. 2012, 38, 179–206. [Google Scholar] [CrossRef]
- Thorsteinsdottir, F.; Cardoso, I.; Keller, A.; Stougaard, M.; Frederiksen, P.; Cohen, A.S.; Maslova, E.; Jacobsen, R.; Backer, V.; Heitmann, B.L. Neonatal Vitamin D Status and Risk of Asthma in Childhood: Results from the D-Tect Study. Nutrients 2020, 12, 842. [Google Scholar] [CrossRef] [Green Version]
- Lai, G.-R.; Lee, Y.-F.; Yan, S.-J.; Ting, H.-J. Active vitamin D induces gene-specific hypomethylation in prostate cancer cells developing vitamin D resistance. Am. J. Physiol. Cell Physiol. 2020, 318, C836–C847. [Google Scholar] [CrossRef]
- Leão, A.A.P.; Fritz, C.K.; Dias, M.R.M.G.; Carvalho, J.A.R.; Mascarenhas, L.P.G.; Cat, M.N.L.; Radominski, R.; Nesi-França, S. Bone mass and dietary intake in children and adolescents with type 1 diabetes mellitus. J. Diabetes Complicat. 2020, 34, 107573. [Google Scholar] [CrossRef]
- Hyppönen, E. Vitamin D and increasing incidence of type 1 diabetes—evidence for an association? Diabetes Obes. Metab. 2010, 12, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Li, J.; Zhai, M.; Yao, X.; Liu, H.; Deng, T.; Cai, H.; Zhang, W.; Zhang, W.; Lou, J.; et al. 1,25-(OH)2D3 protects Schwann cells against advanced glycation end products-induced apoptosis through PKA-NF-κB pathway. Life Sci. 2019, 225, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, I.M.; Joner, G.; Jenum, P.A.; Eskild, A.; Brunborg, C.; Torjesen, P.A.; Stene, L.C. Vitamin D-binding protein and 25-hydroxyvitamin D during pregnancy in mothers whose children later developed type 1 diabetes. Diabetes. Metab. Res. Rev. 2016, 32, 883–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacobsen, R.; Frederiksen, P.; Heitmann, B.L. Exposure to sunshine early in life prevented development of type 1 diabetes in Danish boys. J. Pediatr. Endocrinol. Metab. 2016, 29, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Valizadeh, M.; Piri, Z.; Mohammadian, F.; Kamali, K.; Amir Moghadami, H.R. The Impact of Vitamin D Supplementation on Post-Partum Glucose Tolerance and Insulin Resistance in Gestational Diabetes: A Randomized Controlled Trial. Int. J. Endocrinol. Metab. 2016, 14, e34312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karras, S.N.; Wagner, C.L.; Castracane, V.D. Understanding vitamin D metabolism in pregnancy: From physiology to pathophysiology and clinical outcomes. Metabolism. 2018, 86, 112–123. [Google Scholar] [CrossRef]
- Mohr, S.; Garland, C.; Gorham, E.; Garland, F. The association between ultraviolet B irradiance, vitamin D status and incidence rates of type 1 diabetes in 51 regions worldwide. Diabetologia 2008, 51, 1391–1398. [Google Scholar] [CrossRef] [Green Version]
- Cherrie, M.P.C.; Wheeler, B.W.; White, M.P.; Sarran, C.E.; Osborne, N.J. Coastal climate is associated with elevated solar irradiance and higher 25(OH)D level. Environ. Int. 2015, 77, 76–84. [Google Scholar] [CrossRef]
- Abela, A.G.; Fava, S. Incidence of type 1 diabetes and distance from the sea: A descriptive epidemiological study. J. Diabetes 2019, 11, 345–347. [Google Scholar] [CrossRef] [Green Version]
- Ahadi, M.; Tabatabaeiyan, M.; Moazzami, K. Association between environmental factors and risk of type 1 diabetes—A case-control study. Endokrynol. Pol. 2011, 62, 134–137. [Google Scholar]
- Awadalla, N.; Hegazy, A.; El-Salam, M.; Elhady, M. Environmental Factors Associated with Type 1 Diabetes Development: A Case Control Study in Egypt. Int. J. Environ. Res. Public Health 2017, 14, 615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargese, E.; Samuel, S.M.; Liskova, A.; Kubatka, P.; Busselberg, D. Diabetes and coronavirus (SARS-CoV-2): Molecular mechanism of metformin intervention and the scientific basis of drug repurposing. PLoS Pathog. 2021, 17, e1009634. [Google Scholar] [CrossRef] [PubMed]
- Bathina, S.; Das, U.N. Resolvin D1 Decreases Severity of Streptozotocin-Induced Type 1 Diabetes Mellitus by Enhancing BDNF Levels, Reducing Oxidative Stress, and Suppressing Inflammation. Int. J. Mol. Sci. 2021, 22, 1516. [Google Scholar] [CrossRef] [PubMed]
- Michalska, M.; Wąż, P.; Zorena, K.; Bartoszewicz, M.; Korzeniowska, K.; Krawczyk, S.; Beń-Skowronek, I.; Myśliwiec, M. Potential effects of microbial air quality on the number of new cases of diabetes type 1 in children in two regions of Poland: A pilot study. Infect. Drug Resist. 2019, 12, 2323–2334. [Google Scholar] [CrossRef] [Green Version]
- Suwanwongse, K.; Shabarek, N. Newly diagnosed diabetes mellitus, DKA, and COVID-19: Causality or coincidence? A report of three cases. J. Med. Virol. 2021, 93, 1150–1153. [Google Scholar] [CrossRef]
- Zorena, K.; Kowalewska, B.; Szmigiero-Kawko, M.; Wąż, P.; Myśliwiec, M. No effect of yeast-like fungi on lipid metabolism and vascular endothelial growth factor level in children and adolescents with type 1 diabetes mellitus. Ital. J. Pediatr. 2016, 42, 107. [Google Scholar] [CrossRef] [Green Version]
- Altobelli, E.; Petrocelli, R.; Verrotti, A.; Valenti, M. Infections and risk of type I diabetes in childhood: A population-based case-control study. Eur. J. Epidemiol. 2003, 18, 425–430. [Google Scholar] [CrossRef]
- Costa, F.R.C.; Françozo, M.C.S.; de Oliveira, G.G.; Ignacio, A.; Castoldi, A.; Zamboni, D.S.; Ramos, S.G.; Câmara, N.O.; de Zoete, M.R.; Palm, N.W.; et al. Gut microbiota translocation to the pancreatic lymph nodes triggers NOD2 activation and contributes to T1D onset. J. Exp. Med. 2016, 213, 1223–1239. [Google Scholar] [CrossRef]
- Rubino, F.; Amiel, S.A.; Zimmet, P.; Alberti, G.; Bornstein, S.; Eckel, R.H.; Mingrone, G.; Boehm, B.; Cooper, M.E.; Chai, Z.; et al. New-Onset Diabetes in Covid-19. N. Engl. J. Med. 2020, 383, 789–790. [Google Scholar] [CrossRef]
- Netea, M.G.; van Crevel, R. BCG-induced protection: Effects on innate immune memory. Semin. Immunol. 2014, 26, 512–517. [Google Scholar] [CrossRef]
- Gosiewski, T.; Salamon, D.; Szopa, M.; Sroka, A.; Malecki, M.T.; Bulanda, M. Quantitative evaluation of fungi of the genus Candida in the feces of adult patients with type 1 and 2 diabetes—A pilot study. Gut Pathog. 2014, 6, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kowalewska, B.; Kawko, M.; Zorena, K.; Myśliwiec, M. Yeast-like fungi in the gastrointestinal tract in children and adolescents with diabetes type 1. Pediatr. Endocrinol. Diabetes Metab. 2014, 20, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Kowalewska, B.; Zorena, K.; Szmigiero-Kawko, M.; Wąż, P.; Myśliwiec, M. Higher diversity in fungal species discriminates children with type 1 diabetes mellitus from healthy control. Patient Prefer. Adherence 2016, 10, 591–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esposito, S.; Toni, G.; Tascini, G.; Santi, E.; Berioli, M.G.; Principi, N. Environmental Factors Associated With Type 1 Diabetes. Front. Endocrinol. 2019, 10, 592. [Google Scholar] [CrossRef]
- Shirakawa, J. Pancreatic β-cell fate in subjects with COVID-19. J. Diabetes Investig. 2021, 12, 2126–2128. [Google Scholar] [CrossRef]
- Krogvold, L.; Edwin, B.; Buanes, T.; Frisk, G.; Skog, O.; Anagandula, M.; Korsgren, O.; Undlien, D.; Eike, M.C.; Richardson, S.J.; et al. Detection of a Low-Grade Enteroviral Infection in the Islets of Langerhans of Living Patients Newly Diagnosed with Type 1 Diabetes. Diabetes 2015, 64, 1682–1687. [Google Scholar] [CrossRef] [Green Version]
- Kalliora, M.I.; Vazeou, A.; Delis, D.; Bozas, E.; Thymelli, I.; Bartsocas, C.S. Seasonal variation of type 1 diabetes mellitus diagnosis in Greek children. Hormones 2011, 10, 67–71. [Google Scholar] [CrossRef] [Green Version]
- Kostopoulou, E.; Papachatzi, E.; Skiadopoulos, S.; Rojas Gil, A.-P.; Dimitriou, G.; Spiliotis, B.E.; Varvarigou, A. Seasonal variation and epidemiological parameters in children from Greece with type 1 diabetes mellitus (T1DM). Pediatr. Res. 2021, 89, 574–578. [Google Scholar] [CrossRef]
- Nagafuchi, S. Regulation of Viral Infection in Diabetes. Biology 2021, 10, 529. [Google Scholar] [CrossRef]
- Honkanen, H.; Oikarinen, S.; Nurminen, N.; Laitinen, O.H.; Huhtala, H.; Lehtonen, J.; Ruokoranta, T.; Hankaniemi, M.M.; Lecouturier, V.; Almond, J.W.; et al. Detection of enteroviruses in stools precedes islet autoimmunity by several months: Possible evidence for slowly operating mechanisms in virus-induced autoimmunity. Diabetologia 2017, 60, 424–431. [Google Scholar] [CrossRef] [Green Version]
- Vehik, K.; Lynch, K.F.; Wong, M.C.; Tian, X.; Ross, M.C.; Gibbs, R.A.; Ajami, N.J.; Petrosino, J.F.; Rewers, M.; Toppari, J.; et al. Prospective virome analyses in young children at increased genetic risk for type 1 diabetes. Nat. Med. 2019, 25, 1865–1872. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, T.; Nakano, Y.; Hayakawa, K.; Yoshimatu, H.; Hori, Y.; Yamanishi, K.; Yamanishi, H.; Ota, T.; Fujimoto, T. Fulminant type 1 diabetes mellitus associated with Coxsackievirus type B1 infection during pregnancy: A case report. J. Med. Case Rep. 2019, 13, 186. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, O.A.J.; Szereday, L. The “Three Amigos” lurking behind type 1 diabetes: Hygiene, gut microbiota and viruses. Acta Microbiol. Immunol. Hung. 2018, 65, 421–438. [Google Scholar] [CrossRef] [PubMed]
- Mine, K.; Nagafuchi, S.; Mori, H.; Takahashi, H.; Anzai, K. SARS-CoV-2 Infection and Pancreatic β Cell Failure. Biology 2021, 11, 22. [Google Scholar] [CrossRef] [PubMed]
- Cuschieri, S.; Grech, S. COVID-19 and diabetes: The why, the what and the how. J. Diabetes Complicat. 2020, 34, 107637. [Google Scholar] [CrossRef] [PubMed]
- Zubkiewicz-Kucharska, A.; Seifert, M.; Stępkowski, M.; Noczyńska, A. Diagnosis of type 1 diabetes during the SARS-CoV-2 pandemic: Does lockdown affect the incidence and clinical status of patients? Adv. Clin. Exp. Med. 2021, 30, 127–134. [Google Scholar] [CrossRef]
- Filippi, C.; von Herrath, M. How viral infections affect the autoimmune process leading to type 1 diabetes. Cell Immunol. 2005, 233, 125–132. [Google Scholar] [CrossRef]
- Christen, U.; Bender, C.; von Herrath, M.G. Infection as a cause of type 1 diabetes? Curr. Opin. Rheumatol. 2012, 24, 417–423. [Google Scholar] [CrossRef]
- Butalia, S.; Kaplan, G.G.; Khokhar, B.; Rabi, D.M. Environmental Risk Factors and Type 1 Diabetes: Past, Present, and Future. Can. J. Diabetes 2016, 40, 586–593. [Google Scholar] [CrossRef]
- Rodriguez-Calvo, T. Enterovirus infection and type 1 diabetes: Unraveling the crime scene. Clin. Exp. Immunol. 2019, 195, 15–24. [Google Scholar] [CrossRef] [Green Version]
- Hodik, M.; Anagandula, M.; Fuxe, J.; Krogvold, L.; Dahl-Jørgensen, K.; Hyöty, H.; Sarmiento, L.; Frisk, G. Consortium, =POD-V Coxsackie-adenovirus receptor expression is enhanced in pancreas from patients with type 1 diabetes. BMJ Open Diabetes Res. Care 2016, 4, e000219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyalwidhe, J.O.; Jurczyk, A.; Satish, B.; Redick, S.; Qaisar, N.; Trombly, M.I.; Vangala, P.; Racicot, R.; Bortell, R.; Harlan, D.M.; et al. Proteomic and Transcriptional Profiles of Human Stem Cell-Derived β Cells Following Enteroviral Challenge. Microorganisms 2020, 8, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lietzén, N.; Hirvonen, K.; Honkimaa, A.; Buchacher, T.; Laiho, J.E.; Oikarinen, S.; Mazur, M.A.; Flodström-Tullberg, M.; Dufour, E.; Sioofy-Khojine, A.-B.; et al. Coxsackievirus B Persistence Modifies the Proteome and the Secretome of Pancreatic Ductal Cells. Iscience 2019, 19, 340–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aarnisalo, J.; Veijola, R.; Vainionpää, R.; Simell, O.; Knip, M.; Ilonen, J. Cytomegalovirus infection in early infancy: Risk of induction and progression of autoimmunity associated with type 1 diabetes. Diabetologia 2008, 51, 769–772. [Google Scholar] [CrossRef] [Green Version]
- Yoneda, S.; Imagawa, A.; Fukui, K.; Uno, S.; Kozawa, J.; Sakai, M.; Yumioka, T.; Iwahashi, H.; Shimomura, I. A Histological Study of Fulminant Type 1 Diabetes Mellitus Related to Human Cytomegalovirus Reactivation. J. Clin. Endocrinol. Metab. 2017, 102, 2394–2400. [Google Scholar] [CrossRef]
- Ekman, I.; Vuorinen, T.; Knip, M.; Veijola, R.; Toppari, J.; Hyöty, H.; Kinnunen, T.; Ilonen, J.; Lempainen, J. Early childhood CMV infection may decelerate the progression to clinical type 1 diabetes. Pediatr. Diabetes 2019, 20, 73–77. [Google Scholar] [CrossRef] [Green Version]
- Al-Hakami, A.M. Pattern of thyroid, celiac, and anti-cyclic citrullinated peptide autoantibodies coexistence with type 1 diabetes mellitus in patients from Southwestern Saudi Arabia. Saudi Med. J. 2016, 37, 386–391. [Google Scholar] [CrossRef]
- Nishiumi, T.; Okamoto, K.; Inamoto, S.; Inoue, N.; Ohnishi, K.; Ohji, G.; Hirota, Y.; Ogawa, W. Case of fulminant type 1 diabetes mellitus associated with parvovirus B19 infection. J. Diabetes Investig. 2014, 5, 472–473. [Google Scholar] [CrossRef] [Green Version]
- Selver Eklioglu, B.; Atabek, M.; Akyürek, N.; Gümüş, M. Parvovirus Infection in a Child Complicated with Diabetic Ketoacidosis and Acute Fulminant Hepatitis: A Case Report. J. Pediatr. Infect. 2017, 11, 92–94. [Google Scholar] [CrossRef]
- O’Bryan, T.A.; Beck, M.J.; Demers, L.M.; Naides, S.J. Human parvovirus B19 infection in children with new onset Type 1 diabetes mellitus. Diabet. Med. 2005, 22, 1778–1779. [Google Scholar] [CrossRef]
- Honeyman, M. How robust is the evidence for viruses in the induction of type 1 diabetes? Curr. Opin. Immunol. 2005, 17, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Blumenfeld, O.; Hampe, C.S.; Shulman, L.; Chen, R.; Laron, Z. For Debate: The Controversy whether Rotavirus Vaccination Attenuates the Incidence of Childhood Type 1 Diabetes. Pediatr. Endocrinol. Rev. 2020, 17, 284–286. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.M.; Tate, J.E.; Jiang, B.; Parashar, U.D. Rotavirus and Type 1 Diabetes—Is There a Connection? A Synthesis of the Evidence. J. Infect. Dis. 2020, 222, 1076–1083. [Google Scholar] [CrossRef] [PubMed]
- Harrison, L.C.; Perrett, K.P.; Jachno, K.; Nolan, T.M.; Honeyman, M.C. Does rotavirus turn on type 1 diabetes? PLoS Pathog. 2019, 15, e1007965. [Google Scholar] [CrossRef]
- Vaarala, O.; Jokinen, J.; Lahdenkari, M.; Leino, T. Rotavirus Vaccination and the Risk of Celiac Disease or Type 1 Diabetes in Finnish Children at Early Life. Pediatr. Infect. Dis. J. 2017, 36, 674–675. [Google Scholar] [CrossRef]
- Glanz, J.M.; Clarke, C.L.; Xu, S.; Daley, M.F.; Shoup, J.A.; Schroeder, E.B.; Lewin, B.J.; McClure, D.L.; Kharbanda, E.; Klein, N.P.; et al. Association Between Rotavirus Vaccination and Type 1 Diabetes in Children. JAMA Pediatr. 2020, 174, 455–462. [Google Scholar] [CrossRef]
- Inns, T.; Fleming, K.M.; Iturriza-Gomara, M.; Hungerford, D. Paediatric rotavirus vaccination, coeliac disease and type 1 diabetes in children: A population-based cohort study. BMC Med. 2021, 19, 147. [Google Scholar] [CrossRef]
- Ramondetti, F.; Sacco, S.; Comelli, M.; Bruno, G.; Falorni, A.; Iannilli, A.; d’Annunzio, G.; Iafusco, D.; Songini, M.; Toni, S.; et al. Type 1 diabetes and measles, mumps and rubella childhood infections within the Italian Insulin-dependent Diabetes Registry. Diabet. Med. 2012, 29, 761–766. [Google Scholar] [CrossRef]
- Korkmaz, H.A.; Ermiş, Ç. A case of immune-mediated type 1 diabetes mellitus due to congenital rubella ınfection. Ann. Pediatr. Endocrinol. Metab. 2019, 24, 68–70. [Google Scholar] [CrossRef] [Green Version]
- Gale, E.A.M. Congenital rubella: Citation virus or viral cause of type 1 diabetes? Diabetologia 2008, 51, 1559–1566. [Google Scholar] [CrossRef] [Green Version]
- Vuorinen, T.; Nikolakaros, G.; Simell, O.; Hyypiä, T.; Vainionpää, R. Mumps and Coxsackie B3 virus infection of human fetal pancreatic islet-like cell clusters. Pancreas 1992, 7, 460–464. [Google Scholar] [CrossRef] [PubMed]
- Precechtelova, J.; Borsanyiova, M.; Sarmirova, S.; Bopegamage, S. Type I diabetes mellitus: Genetic factors and presumptive enteroviral etiology or protection. J. Pathog. 2014, 2014, 738512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levet, S.; Medina, J.; Joanou, J.; Demolder, A.; Queruel, N.; Réant, K.; Normand, M.; Seffals, M.; Dimier, J.; Germi, R.; et al. An ancestral retroviral protein identified as a therapeutic target in type-1 diabetes. JCI Insight 2017, 2, e94387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rathnaiah, G.; Zinniel, D.K.; Bannantine, J.P.; Stabel, J.R.; Gröhn, Y.T.; Collins, M.T.; Barletta, R.G. Pathogenesis, Molecular Genetics, and Genomics of Mycobacterium avium subsp. paratuberculosis, the Etiologic Agent of Johne’s Disease. Front. Vet. Sci. 2017, 4, 187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niegowska, M.; Wajda-Cuszlag, M.; Stępień-Ptak, G.; Trojanek, J.; Michałkiewicz, J.; Szalecki, M.; Sechi, L.A. Anti-HERV-W(Env) antibodies are correlated with seroreactivity against Mycobacterium avium subsp. paratuberculosis in children and youths at T1D risk. Sci. Rep. 2019, 9, 6282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ata, A.; Jalilova, A.; Kırkgöz, T.; Işıklar, H.; Demir, G.; Altınok, Y.A.; Özkan, B.; Zeytinlioğlu, A.; Darcan, Ş.; Özen, S.; et al. Does COVID-19 predispose patients to type 1 diabetes mellitus? Clin. Pediatr. Endocrinol. 2022, 31, 33–37. [Google Scholar] [CrossRef]
- Lança, A.; Rodrigues, C.; Diamantino, C.; Fitas, A.L. COVID-19 in two children with new-onset diabetes: Case reports. BMJ Case Rep. 2022, 15, e247309. [Google Scholar] [CrossRef]
- Figueroa-Pizano, M.D.; Campa-Mada, A.C.; Carvajal-Millan, E.; Martinez-Robinson, K.G.; Chu, A.R. The underlying mechanisms for severe COVID-19 progression in people with diabetes mellitus: A critical review. AIMS Public Health 2021, 8, 720–742. [Google Scholar] [CrossRef]
- Demeterco-Berggren, C.; Ebekozien, O.; Rompicherla, S.; Jacobsen, L.; Accacha, S.; Gallagher, M.P.; Todd Alonso, G.; Seyoum, B.; Vendrame, F.; Haw, J.S.; et al. Age and Hospitalization Risk in People with Type 1 Diabetes and COVID-19: Data From the T1D Exchange Surveillance Study. J. Clin. Endocrinol. Metab. 2022, 107, 410–418. [Google Scholar] [CrossRef]
- Deinhardt-Emmer, S.; Wittschieber, D.; Sanft, J.; Kleemann, S.; Elschner, S.; Haupt, K.F.; Vau, V.; Häring, C.; Rödel, J.; Henke, A.; et al. Early postmortem mapping of SARS-CoV-2 RNA in patients with COVID-19 and the correlation with tissue damage. Elife 2021, 10, e60361. [Google Scholar] [CrossRef]
- Ding, Y.; He, L.; Zhang, Q.; Huang, Z.; Che, X.; Hou, J.; Wang, H.; Shen, H.; Qiu, L.; Li, Z.; et al. Organ distribution of severe acute respiratory syndrome(SARS) associated coronavirus(SARS-CoV) in SARS patients: Implications for pathogenesis and virus transmission pathways. J. Pathol. 2004, 203, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Tashiro, M.; Izumikawa, K.; Minematsu, A.; Hirano, K.; Iwanaga, N.; Ide, S.; Mihara, T.; Hosogaya, N.; Takazono, T.; Morinaga, Y.; et al. Antifungal Susceptibilities of Aspergillus fumigatus Clinical Isolates Obtained in Nagasaki, Japan. Antimicrob. Agents Chemother. 2011, 56, 584–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarkar, A.; Mukherjee, A.; Ghoshal, A.; Kundu, S.; Mitra, S. Occurrence of allergic bronchopulmonary mycosis in patients with asthma: An Eastern India experience. Lung India 2010, 27, 212–216. [Google Scholar] [CrossRef] [PubMed]
- Kallawicha, K.; Chuang, Y.-C.; Lung, S.-C.C.; Wu, C.-F.; Han, B.-C.; Ting, Y.-F.; Chao, H.J. Outpatient Visits for Allergic Diseases are Associated with Exposure to Ambient Fungal Spores in the Greater Taipei Area. Aerosol Air Qual. Res. 2018, 18, 2077–2085. [Google Scholar] [CrossRef]
- Xie, Z.; Li, Y.; Lu, R.; Li, W.; Fan, C.; Liu, P.; Wang, J.; Wang, W. Characteristics of total airborne microbes at various air quality levels. J. Aerosol Sci. 2018, 116, 57–65. [Google Scholar] [CrossRef]
- Walser, S.M.; Gerstner, D.G.; Brenner, B.; Bünger, J.; Eikmann, T.; Janssen, B.; Kolb, S.; Kolk, A.; Nowak, D.; Raulf, M.; et al. Evaluation of exposure-response relationships for health effects of microbial bioaerosols—A systematic review. Int. J. Hyg. Environ. Health 2015, 218, 577–589. [Google Scholar] [CrossRef]
- Yang, C.; Yang, X.; Zhao, B. Person to person droplets transmission characteristics in unidirectional ventilated protective isolation room: The impact of initial droplet size. Build. Simul. 2016, 9, 597–606. [Google Scholar] [CrossRef]
- Wei, J.; Li, Y. Airborne spread of infectious agents in the indoor environment. Am. J. Infect. Control 2016, 44, S102–S108. [Google Scholar] [CrossRef]
- Sturm, R. Modeling the deposition of bioaerosols with variable size and shape in the human respiratory tract—A review. J. Adv. Res. 2012, 3, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Michalska, M.; Bartoszewicz, M.; Wąż, P.; Kozaczuk, S.; Beń-Skowronek, I.; Zorena, K. PM10 concentration and microbiological assessment of air in relation to the number of acute cases of type 1 diabetes mellitus in the Lubelskie Voivodeship. Preliminary report. Pediatr. Endocrinol. Diabetes Metab. 2017, 23, 70–76. [Google Scholar] [CrossRef]
- Knutsen, A.P. Allergic bronchopulmonary aspergillosis in asthma. Expert Rev. Clin. Immunol. 2017, 13, 11–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chowdhary, A.; Sharma, C.; Kathuria, S.; Hagen, F.; Meis, J.F. Azole-resistant Aspergillus fumigatus with the environmental TR46/Y121F/T289A mutation in India. J. Antimicrob. Chemother. 2014, 69, 555–557. [Google Scholar] [CrossRef] [PubMed]
- McCrory, M.C.; Moore, B.A.; Nakagawa, T.A.; Givner, L.B.; Jason, D.R.; Palavecino, E.L.; Ajizian, S.J. Disseminated mucormycosis in an adolescent with newly diagnosed diabetes mellitus. Pediatr. Infect. Dis. J. 2014, 33, 1094–1096. [Google Scholar] [CrossRef] [PubMed]
- Dayal, D.; Bakshi, J.; Jain, P.; Shivaprakash, M.R.; Singhi, S. Outcome of Rhino-Sinus Mucormycosis in Children with Type 1 Diabetes. Indian J. Pediatr. 2015, 82, 651–652. [Google Scholar] [CrossRef]
- Pennell, E.; Pecson, I.; Nakamura, C.; Galvis, A.E. Pulmonary mucormycosis in an adolescent female with type 1 diabetes mellitus. IDCases 2018, 14, e00474. [Google Scholar] [CrossRef]
- Durazzo, M.; Ferro, A.; Gruden, G. Gastrointestinal Microbiota and Type 1 Diabetes Mellitus: The State of Art. J. Clin. Med. 2019, 8, 1843. [Google Scholar] [CrossRef] [Green Version]
- Zheng, P.; Li, Z.; Zhou, Z. Gut microbiome in type 1 diabetes: A comprehensive review. Diabetes. Metab. Res. Rev. 2018, 34, e3043. [Google Scholar] [CrossRef]
- Fallani, M.; Young, D.; Scott, J.; Norin, E.; Amarri, S.; Adam, R.; Aguilera, M.; Khanna, S.; Gil, A.; Edwards, C.A.; et al. Intestinal Microbiota of 6-week-old Infants Across Europe: Geographic Influence Beyond Delivery Mode, Breast-feeding, and Antibiotics. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 77–84. [Google Scholar] [CrossRef]
- Kostic, A.D.; Gevers, D.; Siljander, H.; Vatanen, T.; Hyötyläinen, T.; Hämäläinen, A.-M.; Peet, A.; Tillmann, V.; Pöhö, P.; Mattila, I.; et al. The Dynamics of the Human Infant Gut Microbiome in Development and in Progression toward Type 1 Diabetes. Cell Host Microbe 2015, 17, 260–273. [Google Scholar] [CrossRef] [Green Version]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The role of inflammation in diabetes: Current concepts and future perspectives. Eur. Cardiol. Rev. 2019, 14, 50–59. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Ley, R.E.; Volchkov, P.Y.; Stranges, P.B.; Avanesyan, L.; Stonebraker, A.C.; Hu, C.; Wong, F.S.; Szot, G.L.; Bluestone, J.A.; et al. Innate immunity and intestinal microbiota in the development of Type 1 diabetes. Nature 2008, 455, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Patterson, E.; Marques, T.; O’Sullivan, O.; Fitzgerald, P.; Ross, R.P.; Fitzgerald, G.; Cotter, P.; Dinan, T.; Cryan, J.; Stanton, C. Streptozotocin-induced type-1-diabetes disease onset in rats is associated with altered intestinal microbiota composition coupled with decreased diversity. Microbiology 2015, 161, 182–193. [Google Scholar] [CrossRef] [PubMed]
- Sonnenburg, E.D.; Zheng, H.; Joglekar, P.; Higginbottom, S.K.; Firbank, S.J.; Bolam, D.N.; Sonnenburg, J.L. Specificity of Polysaccharide Use in Intestinal Bacteroides Species Determines Diet-Induced Microbiota Alterations. Cell 2010, 141, 1241–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Shea, E.F.; Cotter, P.D.; Stanton, C.; Ross, R.P.; Hill, C. Production of bioactive substances by intestinal bacteria as a basis for explaining probiotic mechanisms: Bacteriocins and conjugated linoleic acid. Int. J. Food Microbiol. 2012, 152, 189–205. [Google Scholar] [CrossRef]
- Devaraj, S.; Hemarajata, P.; Versalovic, J. The Human Gut Microbiome and Body Metabolism: Implications for Obesity and Diabetes. Clin. Chem. 2013, 59, 617–628. [Google Scholar] [CrossRef] [Green Version]
- Woting, A.; Blaut, M. The Intestinal Microbiota in Metabolic Disease. Nutrients 2016, 8, 202. [Google Scholar] [CrossRef]
- Chen, X.; Devaraj, S. Gut Microbiome in Obesity, Metabolic Syndrome, and Diabetes. Curr. Diab. Rep. 2018, 18, 129. [Google Scholar] [CrossRef]
- Carvalho, B.M.; Guadagnini, D.; Tsukumo, D.M.L.; Schenka, A.A.; Latuf-Filho, P.; Vassallo, J.; Dias, J.C.; Kubota, L.T.; Carvalheira, J.B.C.; Saad, M.J.A. Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia 2012, 55, 2823–2834. [Google Scholar] [CrossRef]
- Brown, K.; DeCoffe, D.; Molcan, E.; Gibson, D.L. Diet-Induced Dysbiosis of the Intestinal Microbiota and the Effects on Immunity and Disease. Nutrients 2012, 4, 1095–1119. [Google Scholar] [CrossRef] [Green Version]
- Calcinaro, F.; Dionisi, S.; Marinaro, M.; Candeloro, P.; Bonato, V.; Marzotti, S.; Corneli, R.B.; Ferretti, E.; Gulino, A.; Grasso, F.; et al. Oral probiotic administration induces interleukin-10 production and prevents spontaneous autoimmune diabetes in the non-obese diabetic mouse. Diabetologia 2005, 48, 1565–1575. [Google Scholar] [CrossRef] [Green Version]
- Pahwa, R.; Balderas, M.; Jialal, I.; Chen, X.; Luna, R.A.; Devaraj, S. Gut Microbiome and Inflammation: A Study of Diabetic Inflammasome-Knockout Mice. J. Diabetes Res. 2017, 2017, 6519785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roesch, L.F.; Lorca, G.L.; Casella, G.; Giongo, A.; Naranjo, A.; Pionzio, A.M.; Li, N.; Mai, V.; Wasserfall, C.H.; Schatz, D.; et al. Culture-independent identification of gut bacteria correlated with the onset of diabetes in a rat model. ISME J. 2009, 3, 536–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tai, N.; Peng, J.; Liu, F.; Gulden, E.; Hu, Y.; Zhang, X.; Chen, L.; Wong, F.S.; Wen, L. Microbial antigen mimics activate diabetogenic CD8 T cells in NOD mice. J. Exp. Med. 2016, 213, 2129–2146. [Google Scholar] [CrossRef] [PubMed]
- Patel, M.K.; Antoni, S.; Danovaro-Holliday, M.C.; Desai, S.; Gacic-Dobo, M.; Nedelec, Y.; Kretsinger, K. The epidemiology of rubella, 2007–2018: An ecological analysis of surveillance data. Lancet Glob. Health 2020, 8, e1399–e1407. [Google Scholar] [CrossRef]
- Adamo, G.; Baccolini, V.; Massimi, A.; Barbato, D.; Cocchiara, R.; Di Paolo, C.; Mele, A.; Cianfanelli, S.; Angelozzi, A.; Castellani, F.; et al. Towards elimination of measles and rubella in Italy: Progress and challenges. PLoS ONE 2019, 14, e0226513. [Google Scholar] [CrossRef]
- Zhang, M.; Zhu, Y. Association between particulate matter pollution and the incidence of mumps in 31 provinces from China. Environ. Sci. Pollut. Res. 2021, 28, 51210–51216. [Google Scholar] [CrossRef]
- Mishra, S.; Wang, S.; Nagpal, R.; Miller, B.; Singh, R.; Taraphder, S.; Yadav, H. Probiotics and Prebiotics for the amelioration of type 1 diabetes: Present and future perspectives. Microorganisms 2019, 7, 67. [Google Scholar] [CrossRef] [Green Version]


{kind=link}
| No. | Authors | Climatic Conditions, Lack of Vitamin D Deficiency, Cow’s Milk Feeding | Consequences |
|---|---|---|---|
| 56 | Waernbaum and Dahlquist, 2016 | Climate/air temperature and the number of sunshine hours | Low mean temperature rather than few sunshine hours are associated with an increased incidence of type 1 diabetes in children. |
| 63 | Sørensen et al., 2016 | Vitamin D deficiency in pregnancy | Low level of 25 (OH) D during the third trimester of pregnancy may be associated with a higher risk of developing diabetes in the offspring. |
| 64 | Jacobsen et al., 2016 | Sun exposure/Vitamin D deficiency in pregnancy | Higher sun exposure in the third trimester of pregnancy is associated with a lower risk (HR) of T1D at 5 to 9 years of age. |
| 67 | Mohr et al., 2008 | Intensity of UVB radiation and the number of hours of sunshine per day | The low intensity of UVB radiation at higher latitudes in both hemispheres (R2 for latitude = 0.25, p < 0.0001) and the number of hours of sunshine per day were associated with a much higher incidence of T1D in childhood. |
| 69 | Abela and Fava, 2019 | Climatic conditions | A significant negative association was found between T1D incidence and shortest distance from sea, mean hours of sunshine, and mean temperature, and a positive association was found between T1D incidence and latitude. |
| 70 | Ahadi et al., 2011 | Association between environmental factors and risk of type 1 diabetes | Maternal age > 35 years at delivery, duration of > 6 months of cow milk feeding, lack of vitamin D supplementation, and cesarean delivery were significantly associated with diabetes type 1. |
| 88 | Kostopoulou, et al., 2021 | Seasonal variation of T1DM onset | Seasonality of the T1DM diagnosis is shown, with a predominance in the colder months of the year. |
| No. | Authors | Viruses | Consequences |
|---|---|---|---|
| Viruses | |||
| [23] | (Rewers and Ludvigsson, 2016) | Enterovirus (EV) | Development of β-cell autoimmunity, the presence of enterovirus in pancreatic islets of type 1 diabetic patients. |
| [84] | (Esposito, S.; 2014) | Herpesviridae, ParvoviridaeTogaviridae, Paramyxoviridae, Retroviridae, Picornaviridae, | Induce islet autoimmunity and ß-cell damage and reduce insulin production, leading to full-blown T1DM. |
| [86] | (Krogvold et al., 2015) | Enterovirus (EV) | The presence of enterovirus in pancreatic islets of type 1 diabetic patients. |
| [91] | (Vehik et al., 2019) | Enterovirus A, B, mast adenovirus C, Coxsackievirus, adenovirus | Enterovirus B infections may be involved in the development of islet autoimmunity, but not T1DM; in some young children, coxsackie and adenovirus receptor (CXADR) genes independently correlated with islet autoimmunity. |
| [92] | (Hayakawa et al., 2019) | Coxsackievirus B1 | Fulminant T1DM in pregnancy may be associated with Coxsackievirus B1 infection. |
| [99] | (Butalia et al., 2016) | Mumps, Rubella, Rotavirus, Rnterovirus, Cytomegalovirus | Development of β-cell autoimmunity, molecular mimicry, in vitro, viruses may induce markers of inflammation and alter HLA class I molecule expression. |
| [101] | (Hodik et al., 2016) | Coxsackievirus | Thecoxsackie–adenovirusreceptor (CAR) is expressed in pancreatic islets of patients with T1DM. |
| [104] | (Aarnisalo et al., 2008) | Cytomegalovirus (CMV) | Development of beta-cell autoimmunity. |
| [105] | (Yoneda et al., 2017) | Cytomegalovirus (CMV) | Significantly increased numbers of alpha cells expressing RIG-I and IRF3 development and progression of T1DM. |
| [106] | (Ekman, et al., 2019) | Cytomegalovirus (CMV) | Development and progression of T1DM. |
| [107] | (Al-Hakami, 2016) | Cytomegalovirus (CMV) | No correlation between T1DMandvirus infectivity. |
| [108] | (Nishiumi et al., 2014) | Parvovirus B19 | Fulminant type 1 diabetes mellitus associated with parvovirus B 19. |
| [109] | (Selver Eklioglu et al., 2017) | Parvovirus B19 | Diabetic ketoacidosis (DKA) and acute fulminan hepatitis. |
| [110] | (O’Bryan et al., 2005) | Parvovirus B19 | No association between parvovirus B19 infection and the development of T1DM. |
| [111] | (Honeyman, 2005) | Rotavirus | Molecular mimicry, pancreatic β cell destruction. |
| [114] | (Harrison et al., 2019) | Rotavirus | Molecular mimicry, development of β-cell autoimmunity. |
| [115] | (Vaarala et al., 2017) | Rotavirus | Molecular mimicry, development and progression of T1DM. |
| [116] | (Glanz et al., 2020) | Rotavirus | Molecular mimicry, development of β-cell autoimmunity, rotavirus vaccination does not appear to be associated with T1DM in children. |
| [118] | (Ramondetti et al., 2012) | Rubella virus, Mumps virus | Mumps and rubella viral infections are associated with T1DM. |
| [119] | (Korkmaz and Ermiş, 2019) | Rubella virus | Rubella viral infections are associated with T1DM. |
| [120] | (Gale, 2008) | Rubella virus | Rubella infections predispose to autoimmunity. |
| [121] | (Vuorinen et al., 1992) | Mumps Virus, Coxsackievirus | In vitro model indicated that mumps and coxsackie B3 viruses infect human fetal pancreatic endocrine cells and are able to alter beta-cell function. |
| [122] | (Precechtelova et al., 2014) | Human Cytomegalovirus, Parvovirus, Rotavirus, Coxsackievirus, Human Parechovirus, Enteric Cytopathic Human Orphan viruses, Mumps virus, Rubella virus | Persistent infection, molecular mimicry, autoimmune destruction of pancreatic ≤ β-cells, congenital infection, loss of regulatory T-cells. Infection by rubella virus during pregnancy has been related to increased risk of diabetes in the offspring suffering from congenital rubella syndrome. |
| [123] | (Levet et al., 2017) | Human endogenous retroviruses (HERV) | Pancreatic β cell destruction. |
| [132] | (Parkkonen et al., 1992) | Mumps virus | The infection is associated with an increase in the expression of HLA class I molecules. |
| [133] | (Al-Hakami, 2016) | Viracela, measles | Hemagglutinin peptide and Hsp60 peptide induce the cellular immune response; varicella and measles are risk factors in developing type 1 diabetes. |
| [79] | (Rubino et al., 2020) | SARS-CoV-2 | SARS-CoV-2 virus leads to diabetes via binding to its cellular entry—ACE-2 receptors, which are abundant in pancreatic beta cells and adipose tissue, leading to glucose metabolism abnormalities, and pancreatic beta cells destruction. |
| [75] | (Suwanwongse and Shabarek, 2021) | SARS-CoV-2 | The aberrant immunity caused by SARS-CoV-2 may induce an auto-immune attack on the pancreatic islet cells mimicking the pathogenesis of insulin-dependent DM. |
| [127] | (Lança et al., 2022) | SARS-CoV-2 | Delayed diagnosis, low socioeconomic status, and infection have been associated with diabetic ketoacidosis (DKA) in type 1 diabetes mellitus. |
| No. | Authors | Bacteria and Yeast-Like Fungi | Consequences |
|---|---|---|---|
| Bacteria | |||
| [74] | (Michalska et al., 2019) | Micrococcus luteus, M. lylar, Sarcina luteaKocuriarosea, Staphylococcus aureus, S. epidermidis, S. saprophyticus, Bacillussubtilis, B. cereus, B. mycoides, B. macerans, Pseudomonas aeruginosa, Citrobacter freundi, Enterobacter aerogenes, Aeromonas hydrophila | A relationship between the number of children with recently diagnosed T1DM and the mean concentration of psychrophilic and mesophilic bacteria in the Pomeranian and Lubelskie Voivodeships (p < 0.001). |
| [125] | (Niegowska et al., 2019) | Mycobacterium avium paratuberculosis (MAP) | MAP infection leading to HERV-W antigen expression and enhancing the production of autoantibodies in T1D. |
| Mold fungi | |||
| [74] | (Michalska et al., 2019) | Rhodotorulamucilaginosa, Penicillium chrysogenum, P. viridicatum, Aspergillus niger, A. flavus, Alternaria alternata, Mucor mucedo, Rhizopus nigricans, Geotrychum sp., Biopolaris sp., Chrysosporium sp., Paecilomyces sp. | A significant relationship between the number of new cases of T1DM in children and the mean concentration of fungi in bioaerosols in the Lubelskie Voivodeship (p < 0.001) but not in the Pomeranian Voivodeship (p = NS). |
| [140] | (Michalska et al., 2017) | Penicillium chrysogenum, P. viridicatum, Aspergillus niger, A. flavus, Alternaria alternata, Mucor mucedo, Rhizopus nigricans, Geotrychum sp., Biopolaris sp., Chrysosporium sp., Paecilomyces sp. | A relation between the number of new cases of T1DM and the number of mold fungi (β = 2.923; p < 0.001). |
| [143] | (McCrory et al., 2014) | Mucor sp. | Mucormycosis in children with poorly controlled diabetes and ketoacidosis. |
| [144] | (Dayal et al., 2015) | Mucor sp. | Mucormycosis may extend into the orbit and brain and result in high mortality in children with T1DM. |
| [145] | (Pennell et al., 2018) | Rhizopus sp., Rhizomucor sp., Apophysomyces | Mucormycosis in children with T1DM. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zorena, K.; Michalska, M.; Kurpas, M.; Jaskulak, M.; Murawska, A.; Rostami, S. Environmental Factors and the Risk of Developing Type 1 Diabetes—Old Disease and New Data. Biology 2022, 11, 608. https://doi.org/10.3390/biology11040608
Zorena K, Michalska M, Kurpas M, Jaskulak M, Murawska A, Rostami S. Environmental Factors and the Risk of Developing Type 1 Diabetes—Old Disease and New Data. Biology. 2022; 11(4):608. https://doi.org/10.3390/biology11040608
Chicago/Turabian StyleZorena, Katarzyna, Małgorzata Michalska, Monika Kurpas, Marta Jaskulak, Anna Murawska, and Saeid Rostami. 2022. "Environmental Factors and the Risk of Developing Type 1 Diabetes—Old Disease and New Data" Biology 11, no. 4: 608. https://doi.org/10.3390/biology11040608
APA StyleZorena, K., Michalska, M., Kurpas, M., Jaskulak, M., Murawska, A., & Rostami, S. (2022). Environmental Factors and the Risk of Developing Type 1 Diabetes—Old Disease and New Data. Biology, 11(4), 608. https://doi.org/10.3390/biology11040608