
IL-10 Producing Regulatory B Cells Mediated Protection against Murine Malaria Pathogenesis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
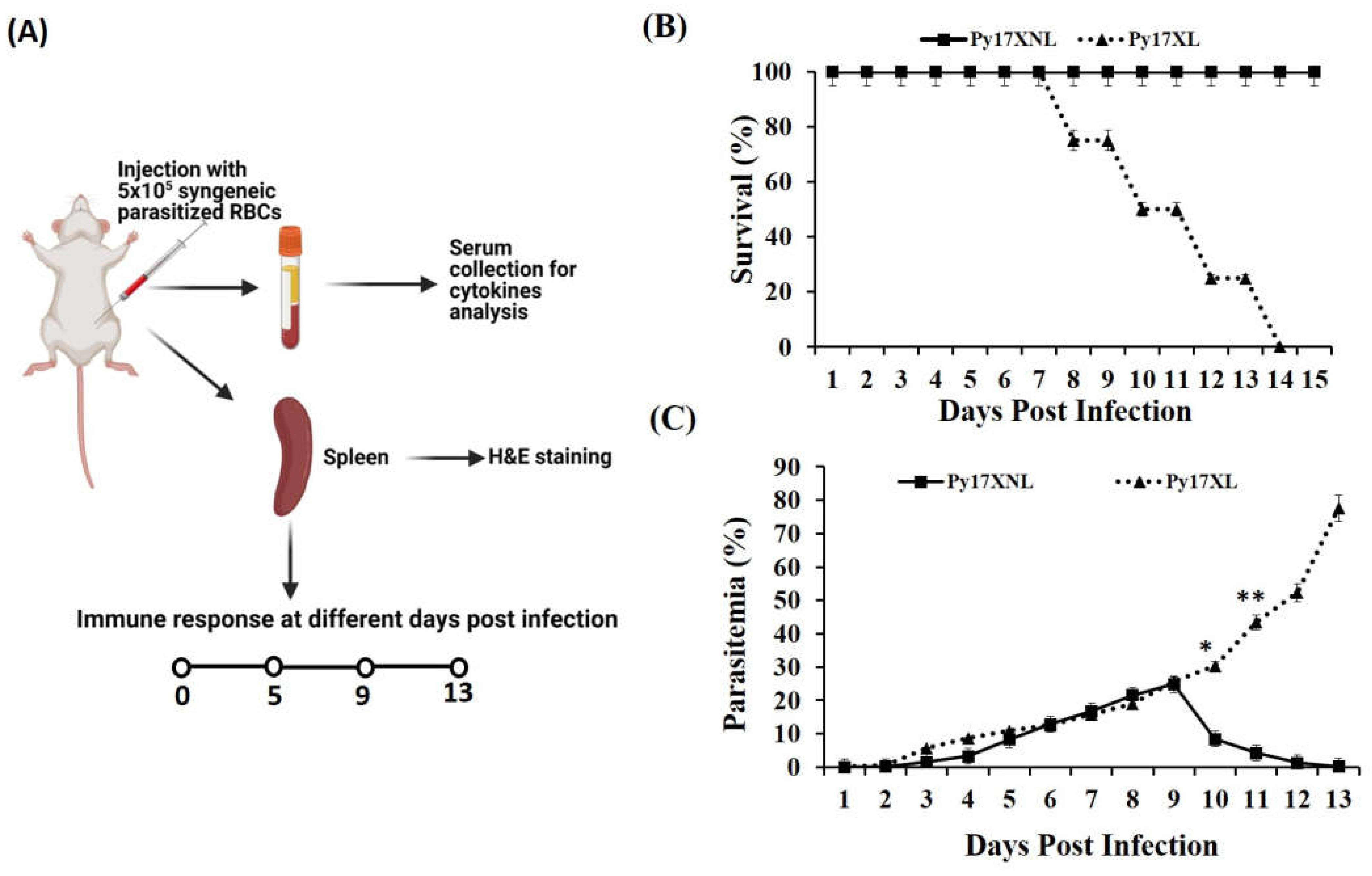
3.1. Parasitaemia and Survival during Lethal and Non-Lethal Plasmodium yoelii Infection
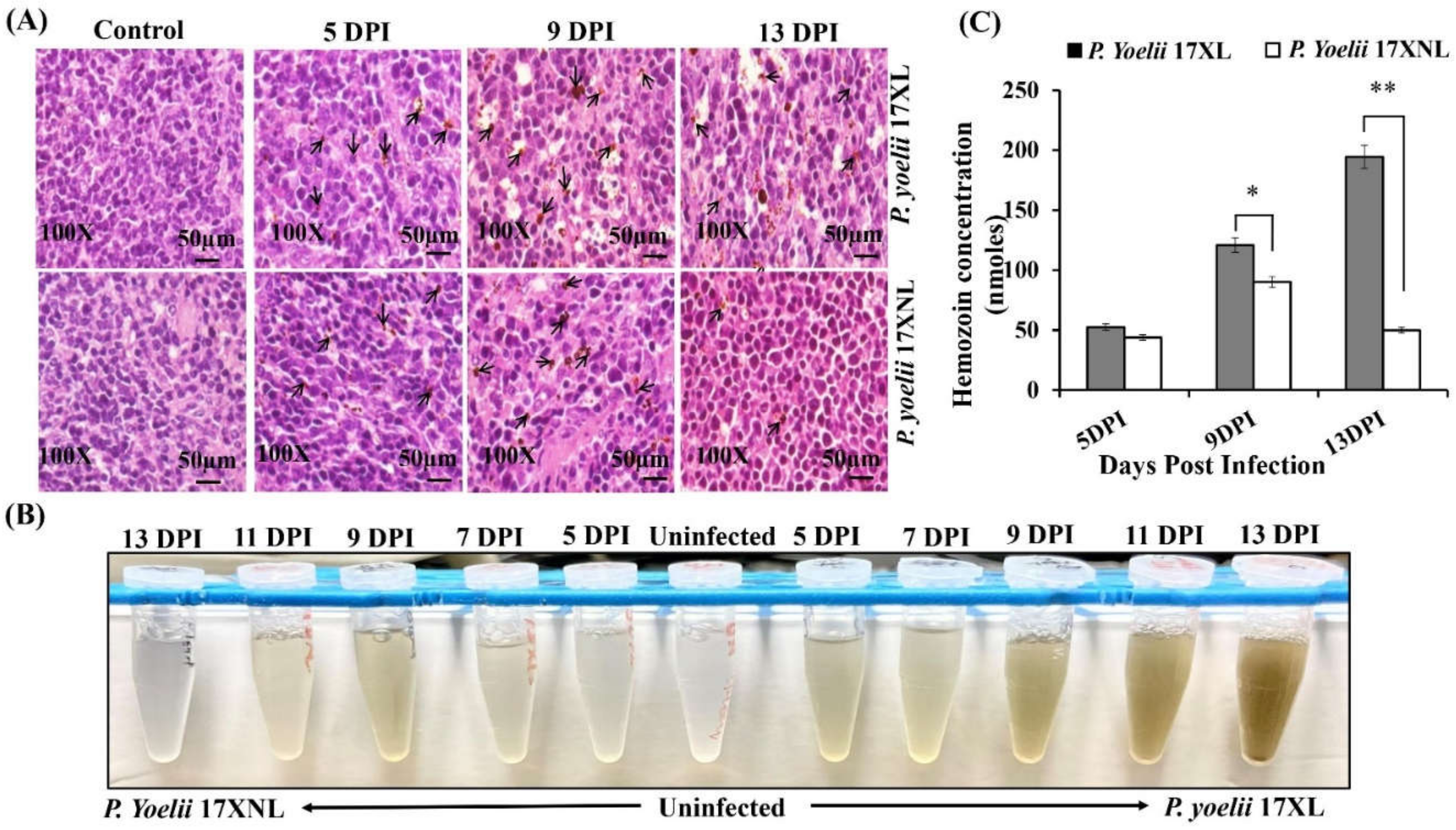
3.2. Spleen Histopathology and Hemozoin Content Analysis during Py 17XL and Py 17XNL Infection
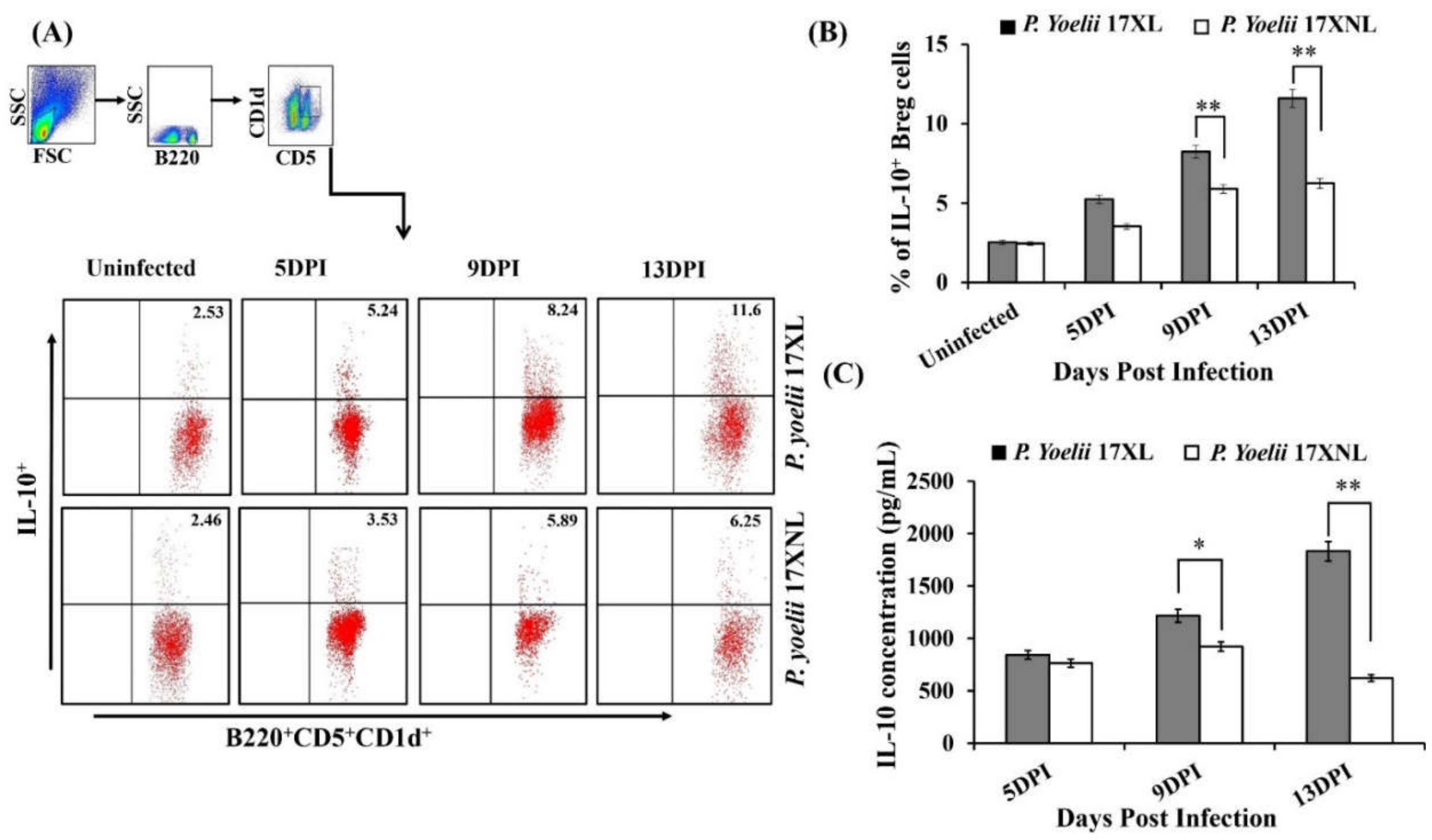
3.3. Dynamics and Expansion of B Cells and IL-10 Producing Regulatory B Cells during Py 17XL and Py 17XNL Infection
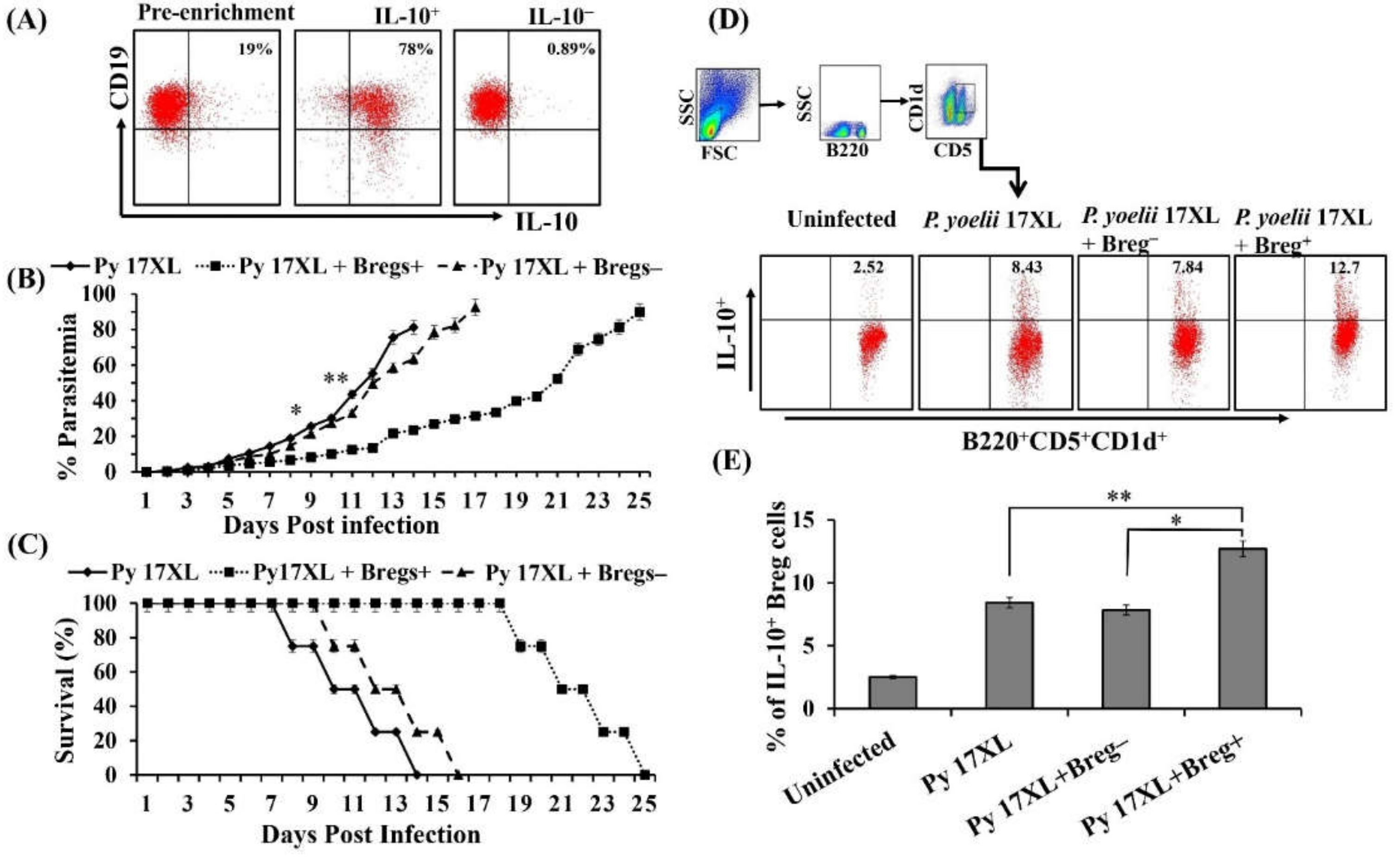
3.4. Adoptive Transferred Regulatory B Cells Mediating Protection
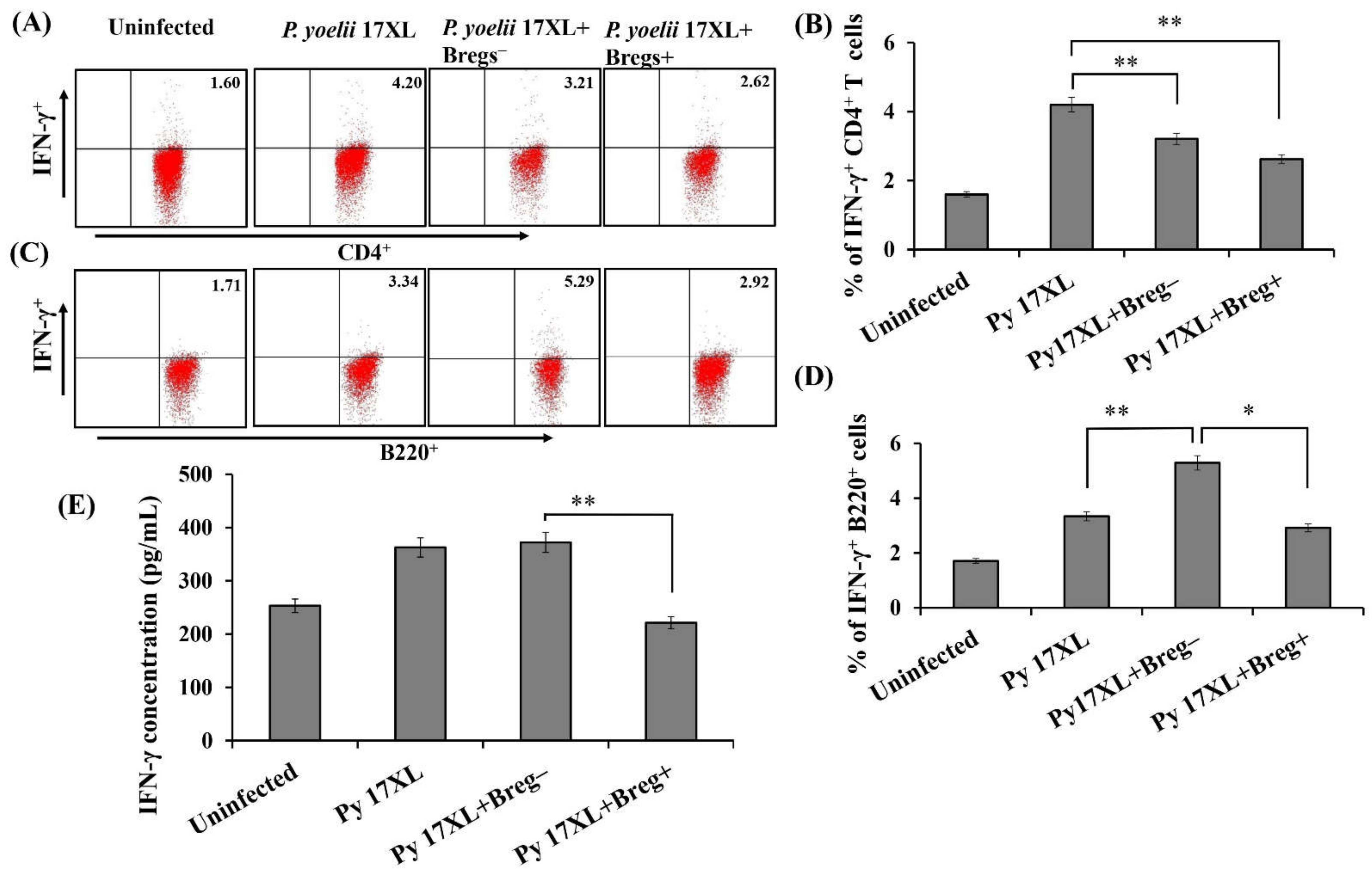
3.5. Effect of Adoptive Transfer of Regulatory B Cells on Pro-Inflammatory Immune Response
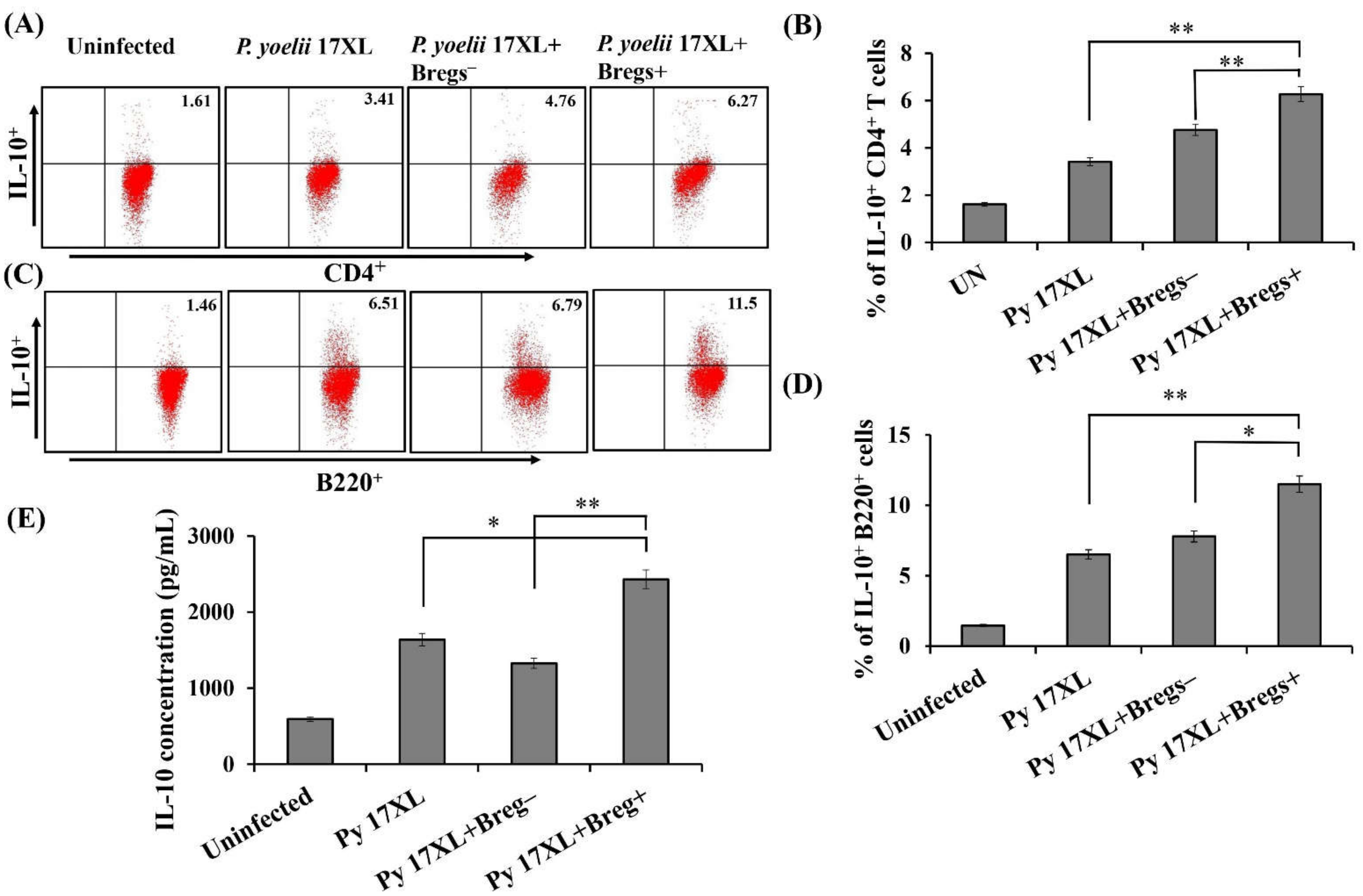
3.6. Effect of Adoptive Transfer of Regulatory B Cells on Anti-Inflammatory Immune Response
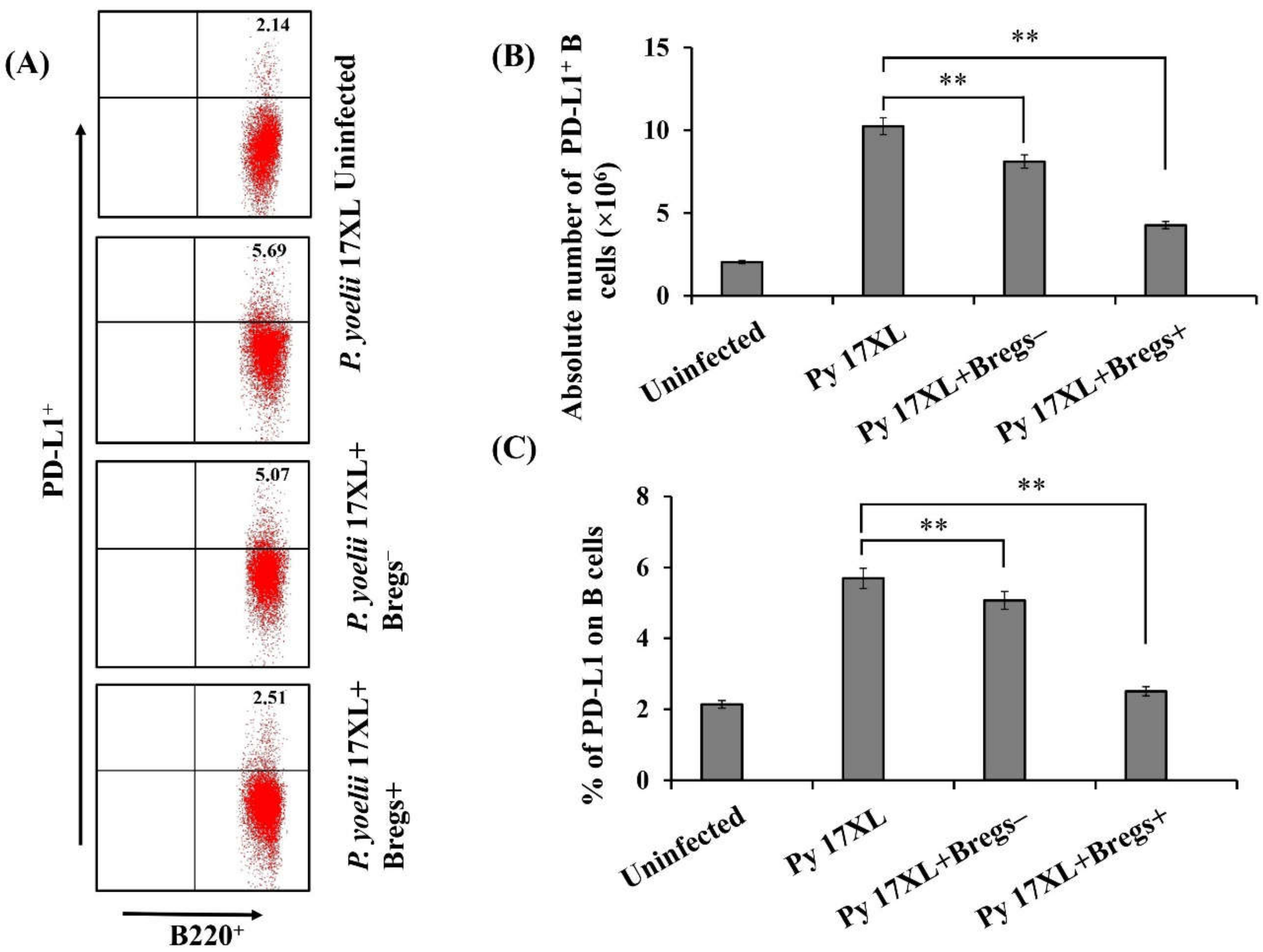
3.7. Effect of Adoptive Transfer of Bregs on PD-L1 Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. World Malaria Report 2021; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Day, N.P.; Hien, T.T.; Schollaardt, T.; Loc, P.P.; Chuong, L.V.; Hong Chau, T.T.; Hoang Mai, N.T.; Phu, N.H.; Sinh, D.X.; White, N.J. The prognostic and pathophysiologic role of pro-and antiinflammatory cytokines in severe malaria. J. Infect. Dis. 1999, 180, 1288–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCall, M.B.; Sauerwein, R.W. Interferon-γ—central mediator of protective immune responses against the pre-erythrocytic and blood stage of malaria. J. Leukoc. Biol. 2010, 88, 1131–1143. [Google Scholar] [CrossRef] [PubMed]
- Claser, C.; De Souza, J.B.; Thorburn, S.G.; Grau, G.E.; Riley, E.M.; Rénia, L.; Hafalla, J.C. Host resistance to Plasmodium-induced acute immune pathology is regulated by interleukin-10 receptor signaling. Infect. Immun. 2017, 85, e00941-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Corraliza, I.; Langhorne, J. A defect in interleukin-10 leads to enhanced malarial disease in Plasmodium chabaudi chabaudi infection in mice. Infect. Immun. 1999, 67, 4435–4442. [Google Scholar] [CrossRef] [Green Version]
- Popa, G.L.; Popa, M.I. Recent Advances in Understanding the Inflammatory Response in Malaria: A Review of the Dual Role of Cytokines. J. Immunol. Res. 2021, 2021, 7785180. [Google Scholar] [CrossRef]
- Riley, E.; Wahl, S.; Perkins, D.; Schofield, L. Regulating immunity to malaria. Parasite Immunol. 2006, 28, 35–49. [Google Scholar] [CrossRef]
- Horne-Debets, J.M.; Faleiro, R.; Karunarathne, D.S.; Liu, X.Q.; Lineburg, K.E.; Poh, C.M.; Grotenbreg, G.M.; Hill, G.R.; MacDonald, K.P.; Good, M.F. PD-1 dependent exhaustion of CD8+ T cells drives chronic malaria. Cell Rep. 2013, 5, 1204–1213. [Google Scholar] [CrossRef] [Green Version]
- Saeidi, A.; Zandi, K.; Cheok, Y.Y.; Saeidi, H.; Wong, W.F.; Lee, C.Y.Q.; Cheong, H.C.; Yong, Y.K.; Larsson, M.; Shankar, E.M. T-Cell Exhaustion in Chronic Infections: Reversing the State of Exhaustion and Reinvigorating Optimal Protective Immune Responses. Front. Immunol. 2018, 9, 2569. [Google Scholar] [CrossRef]
- Rao, M.; Valentini, D.; Dodoo, E.; Zumla, A.; Maeurer, M. Anti-PD-1/PD-L1 therapy for infectious diseases: Learning from the cancer paradigm. Int. J. Infect. Dis. 2017, 56, 221–228. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, M.M.; Riley, E.M. Innate immunity to malaria. Nat. Rev. Immunol. 2004, 4, 169–180. [Google Scholar] [CrossRef]
- Langhorne, J.; Ndungu, F.M.; Sponaas, A.-M.; Marsh, K. Immunity to malaria: More questions than answers. Nat. Immunol. 2008, 9, 725–732. [Google Scholar] [CrossRef]
- Coban, C.; Ishii, K.J.; Horii, T.; Akira, S. Manipulation of host innate immune responses by the malaria parasite. Trends Microbiol. 2007, 15, 271–278. [Google Scholar] [CrossRef]
- Do Rosário, A.P.F.; Lamb, T.; Spence, P.; Stephens, R.; Lang, A.; Roers, A.; Muller, W.; O’Garra, A.; Langhorne, J. IL-27 promotes IL-10 production by effector Th1 CD4+ T cells: A critical mechanism for protection from severe immunopathology during malaria infection. J. Immunol. 2012, 188, 1178–1190. [Google Scholar] [CrossRef] [Green Version]
- Jagannathan, P.; Eccles-James, I.; Bowen, K.; Nankya, F.; Auma, A.; Wamala, S.; Ebusu, C.; Muhindo, M.K.; Arinaitwe, E.; Briggs, J. IFNγ/IL-10 co-producing cells dominate the CD4 response to malaria in highly exposed children. PLoS Pathog. 2014, 10, e1003864. [Google Scholar] [CrossRef] [Green Version]
- Couper, K.N.; Blount, D.G.; Riley, E.M. IL-10: The Master Regulator of Immunity to Infection. J. Immunol. 2008, 180, 5771–5777. [Google Scholar] [CrossRef]
- Saraiva, M.; O’garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Guthmiller, J.J.; Graham, A.C.; Zander, R.A.; Pope, R.L.; Butler, N.S. Cutting Edge: IL-10 Is Essential for the Generation of Germinal Center B Cell Responses and Anti-Plasmodium Humoral Immunity. J. Immunol. 2017, 198, 617–622. [Google Scholar] [CrossRef]
- Kossodo, S.; Monso, C.; Juillard, P.; Velu, T.; Goldman, M.; Grau, G.E. Interleukin-10 modulates susceptibility in experimental cerebral malaria. Immunology 1997, 91, 536–540. [Google Scholar] [CrossRef]
- Lyke, K.; Burges, R.; Cissoko, Y.; Sangare, L.; Dao, M.; Diarra, I.; Kone, A.; Harley, R.; Plowe, C.; Doumbo, O. Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1β), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12 (p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect. Immun. 2004, 72, 5630–5637. [Google Scholar] [CrossRef] [Green Version]
- Wenisch, C.; Parschalk, B.; Narzt, E. Looareesuwan S, Graninger W: Elevated serum levels of IL-10 and IFN-γ in patients with acute Plasmodium falciparum malaria. Clin. Immunol. Immunopathol. 1995, 74, 115–117. [Google Scholar] [CrossRef]
- DiLillo, D.J.; Matsushita, T.; Tedder, T.F. B10 cells and regulatory B cells balance immune responses during inflammation, autoimmunity, and cancer. Ann. N. Y. Acad. Sci. 2010, 1183, 38–57. [Google Scholar] [CrossRef]
- Rosser, E.C.; Mauri, C. Regulatory B cells: Origin, phenotype, and function. Immunity 2015, 42, 607–612. [Google Scholar] [CrossRef] [Green Version]
- Ran, Z.; Yue-Bei, L.; Qiu-Ming, Z.; Huan, Y. Regulatory B Cells and Its Role in Central Nervous System Inflammatory Demyelinating Diseases. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Han, X.; Yang, J.; Zhang, Y.; Zhang, Y.; Cao, H.; Cao, Y.; Qi, Z. Potential role for regulatory B cells as a major source of interleukin-10 in spleen from Plasmodium chabaudi-infected mice. Infect. Immun. 2018, 86, e00016–e00018. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Chen, Y.; Li, Z.; Han, Y.; Sun, Y.; Wang, Q.; Liu, B.; Su, Z. Role of IL-10-producing regulatory B cells in control of cerebral malaria in P lasmodium berghei infected mice. Eur. J. Immunol. 2013, 43, 2907–2918. [Google Scholar] [CrossRef]
- Thakur, R.S.; Tousif, S.; Awasthi, V.; Sanyal, A.; Atul, P.K.; Punia, P.; Das, J. Mesenchymal stem cells play an important role in host protective immune responses against malaria by modulating regulatory T cells. Eur. J. Immunol. 2013, 43, 2070–2077. [Google Scholar] [CrossRef]
- Thakur, R.S.; Awasthi, V.; Sanyal, A.; Chatterjee, S.; Rani, S.; Chauhan, R.; Kalkal, M.; Tiwari, M.; Pande, V.; Das, J. Mesenchymal stem cells protect against malaria pathogenesis by reprogramming erythropoiesis in the bone marrow. Cell Death Discov. 2020, 6, 125. [Google Scholar] [CrossRef]
- Pisciotta, J.M.; Scholl, P.F.; Shuman, J.L.; Shualev, V.; Sullivan, D.J. Quantitative characterization of hemozoin in Plasmodium berghei and vivax. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Del Portillo, H.A.; Ferrer, M.; Brugat, T.; Martin-Jaular, L.; Langhorne, J.; Lacerda, M.V.G. The role of the spleen in malaria. Cell. Microbiol. 2012, 14, 343–355. [Google Scholar] [CrossRef] [PubMed]
- Olivier, M.; Van Den Ham, K.; Shio, M.; Kassa, F.; Fougeray, S. Malarial Pigment Hemozoin and the Innate Inflammatory Response. Front. Immunol. 2014, 5, 25. [Google Scholar] [CrossRef] [PubMed]
- Harris, D.P.; Goodrich, S.; Gerth, A.J.; Peng, S.L.; Lund, F.E. Regulation of IFN-γ Production by B Effector 1 Cells: Essential Roles for T-bet and the IFN-γ Receptor. J. Immunol. 2005, 174, 6781–6790. [Google Scholar] [CrossRef] [Green Version]
- Bao, Y.; Liu, X.; Han, C.; Xu, S.; Xie, B.; Zhang, Q.; Gu, Y.; Hou, J.; Qian, L.; Qian, C.; et al. Identification of IFN-γ-producing innate B cells. Cell Res. 2014, 24, 161–176. [Google Scholar] [CrossRef]
- Bénard, A.; Sakwa, I.; Schierloh, P.; Colom, A.; Mercier, I.; Tailleux, L.; Jouneau, L.; Boudinot, P.; Al-Saati, T.; Lang, R. B cells producing type I IFN modulate macrophage polarization in tuberculosis. Am. J. Respir. Crit. Care Med. 2018, 197, 801–813. [Google Scholar] [CrossRef]
- Beeson, J.G.; Osier, F.H.; Engwerda, C.R. Recent insights into humoral and cellular immune responses against malaria. Trends Parasitol. 2008, 24, 578–584. [Google Scholar] [CrossRef]
- Kalkal, M.; Tiwari, M.; Thakur, R.S.; Awasthi, V.; Pande, V.; Chattopadhyay, D.; Das, J. Mesenchymal Stem Cells: A Novel Therapeutic Approach to Enhance Protective Immunomodulation and Erythropoietic Recovery in Malaria. Stem Cell Rev. Rep. 2021, 17, 1993–2002. [Google Scholar] [CrossRef]
- Artavanis-Tsakonas, K.; Tongren, J.; Riley, E. The war between the malaria parasite and the immune system: Immunity, immunoregulation and immunopathology. Clin. Exp. Immunol. 2003, 133, 145–152. [Google Scholar] [CrossRef]
- King, T.; Lamb, T. Interferon-γ: The Jekyll and Hyde of malaria. PLoS Pathog. 2015, 11, e1005118. [Google Scholar] [CrossRef]
- Chauhan, R.; Awasthi, V.; Thakur, R.S.; Pande, V.; Chattopadhyay, D.; Das, J. CD4+ICOS+Foxp3+: A sub-population of regulatory T cells contribute to malaria pathogenesis. Malaria J. 2022, 21, 32. [Google Scholar] [CrossRef]
- Silveira, E.L.; Dominguez, M.R.; Soares, I.S. To B or not to B: Understanding B cell responses in the development of malaria infection. Front. Immunol. 2018, 9, 2961. [Google Scholar] [CrossRef] [Green Version]
- Von der Weid, T.; Honarvar, N.; Langhorne, J. Gene-targeted mice lacking B cells are unable to eliminate a blood stage malaria infection. J. Immunol. 1996, 156, 2510–2516. [Google Scholar]
- Bouaziz, J.D.; Yanaba, K.; Tedder, T.F. Regulatory B cells as inhibitors of immune responses and inflammation. Immunol. Rev. 2008, 224, 201–214. [Google Scholar] [CrossRef]
- Dai, Y.C.; Zhong, J.; Xu, J.F. Regulatory B cells in infectious disease (Review). Mol. Med. Rep. 2017, 16, 3–10. [Google Scholar] [CrossRef] [Green Version]
- Jansen, K.; Cevhertas, L.; Ma, S.; Satitsuksanoa, P.; Akdis, M.; van de Veen, W. Regulatory B cells, A to Z. Allergy 2021, 76, 2699–2715. [Google Scholar] [CrossRef]
- Asito, A.S.; Moormann, A.M.; Kiprotich, C.; Ng’ang’a, Z.W.; Ploutz-Snyder, R.; Rochford, R. Alterations on peripheral B cell subsets following an acute uncomplicated clinical malaria infection in children. Malar. J. 2008, 7, 238. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, R.T.; Ssewanyana, I.; Wamala, S.; Nankya, F.; Jagannathan, P.; Tappero, J.W.; Mayanja-Kizza, H.; Muhindo, M.K.; Arinaitwe, E.; Kamya, M. B cell sub-types following acute malaria and associations with clinical immunity. Malar. J. 2016, 15, 1–10. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalkal, M.; Chauhan, R.; Thakur, R.S.; Tiwari, M.; Pande, V.; Das, J. IL-10 Producing Regulatory B Cells Mediated Protection against Murine Malaria Pathogenesis. Biology 2022, 11, 669. https://doi.org/10.3390/biology11050669
Kalkal M, Chauhan R, Thakur RS, Tiwari M, Pande V, Das J. IL-10 Producing Regulatory B Cells Mediated Protection against Murine Malaria Pathogenesis. Biology. 2022; 11(5):669. https://doi.org/10.3390/biology11050669
Chicago/Turabian StyleKalkal, Meenu, Rubika Chauhan, Reva Sharan Thakur, Mrinalini Tiwari, Veena Pande, and Jyoti Das. 2022. "IL-10 Producing Regulatory B Cells Mediated Protection against Murine Malaria Pathogenesis" Biology 11, no. 5: 669. https://doi.org/10.3390/biology11050669
APA StyleKalkal, M., Chauhan, R., Thakur, R. S., Tiwari, M., Pande, V., & Das, J. (2022). IL-10 Producing Regulatory B Cells Mediated Protection against Murine Malaria Pathogenesis. Biology, 11(5), 669. https://doi.org/10.3390/biology11050669