
Utilization of Legume-Nodule Bacterial Symbiosis in Phytoremediation of Heavy Metal-Contaminated Soils



Abstract
:Simple Summary
Abstract
1. Introduction
2. Toxic Effect of Metal Stress on Plants
3. Legume–Rhizobium Symbiotic System for Stress Condition Tolerance
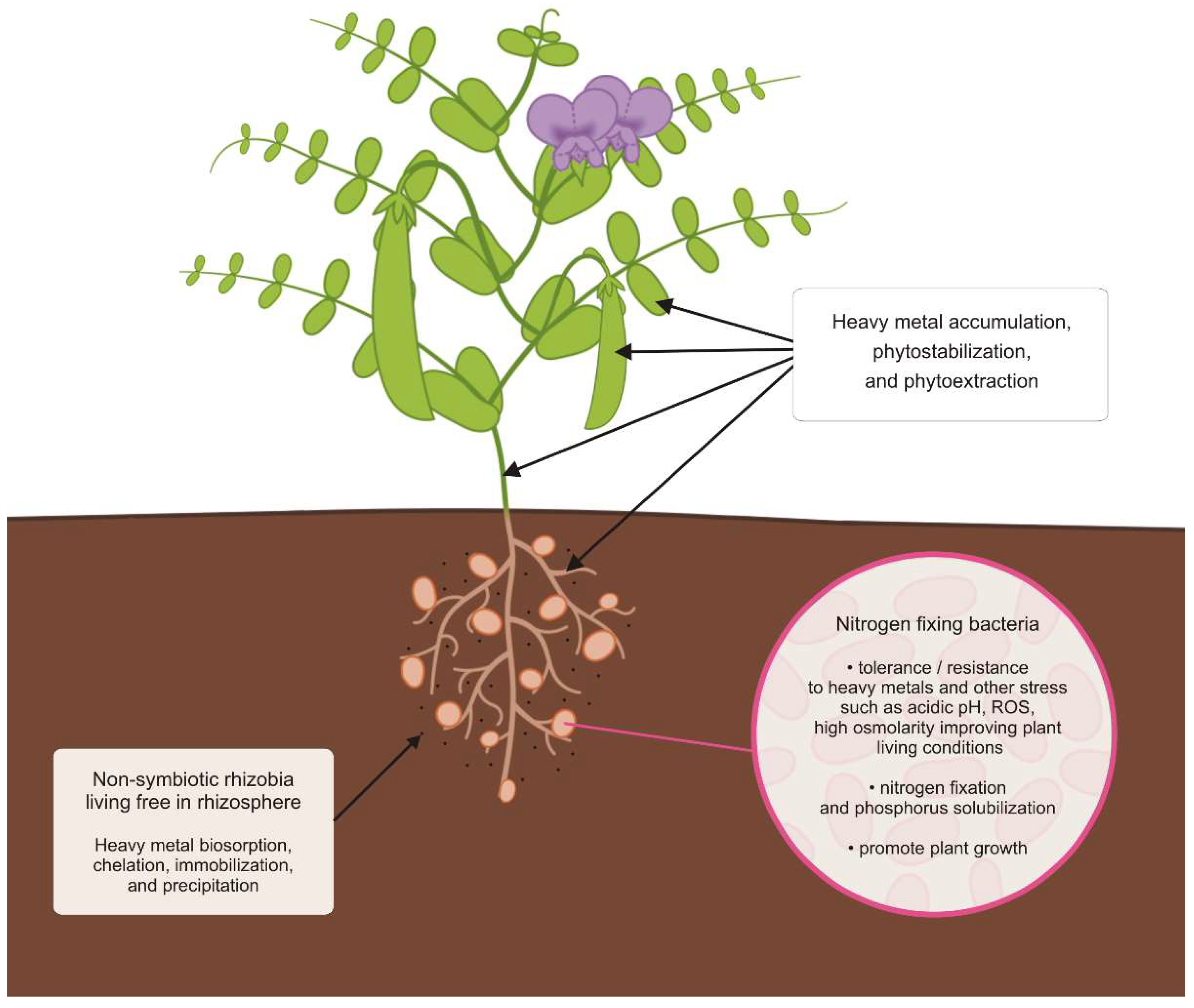
3.1. Plant Growth-Promoting Rhizobia Assisted Legumes for Metal Phytoremediation
3.2. Nitrogen Fixation Bacteria
3.3. Mechanism of the Legume–Rhizobium Interaction
3.4. The Rhizobial Tolerance to Various Stress Conditions
3.5. The Legume–Rhizobium Symbiosis in Heavy Metal Stress Response
4. Phytoremediation by Legume and Associated Rhizobia
4.1. Legume Growth, Nodulation, and Nitrogen Fixation under Heavy Metal Stress
4.2. The Success of Biorhizoremediation of Heavy Metals
- Improve the soil properties to allow other plants to grow through immobilization of contaminants, enhancing organic content, and modifying rhizosphere population;
- Increase diversity of microorganisms, especially rhizobacteria and arbuscular mycorrhiza fungi to improve and stabilize the ecology of the polluted and contaminated field soils;
- Provide additional nitrogen and phosphorus compounds to the field soil to improve its fertility and ability to support biological growth;
- Improve plant living conditions to allow legumes and other plants to grow in HM and other stress conditions;
- Promote plant growth.
4.3. The Enhancement of Heavy Metal Stress Response by other PGPB in Addition to the Legume–Rhizobium Symbiosis
5. Genetic Engineering in Improvement of Leguminous Plants and Their Rhizobial Partners for Phytoremediation
5.1. General Strategies of Plant Transgenesis for Increasing HM Tolerance and Accumulation

{kind=link}
| Gene (s) | Gene Function | Gene Origin | Gene Host | Effect Comparing to the Wild-Type (WT) Control | References |
|---|---|---|---|---|---|
| GST and CYP2E1 | Glutathione S-transferase Cytochrome 450 2E1 | Human | Medicago sativa L. | Improved tolerance of plants towards Cd/trichloroethylene (TCE) mixture | [333] |
| GST and CYP2E1 | Glutathione S-transferase Cytochrome 450 2E1 | Human | M. sativa L. | Improved tolerance of plants towards Hg/TCE mixture. Ameliorated plant growth with a longer root system enhanced accumulation of pullulans | [334] |
| MtTdp2α | Tyrosyl-DNA phosphodiesterase II | Medicago truncatula Gaertn. | M. truncatula Gaertn. | Improved Cu tolerance. Reduction in necrosis volume, decrease in double strand breaks in DNA | [324] |
| mt4a | Metallothionein | Arabidopsis thaliana L. | M. truncatula (composite plants) | Increased Cu tolerance, reduction in oxidative stress. Improved nodulation | [119] |
| mt4a | Metallothionein | A. thaliana L. | M. truncatula (composite plants) | Enhanced Cu tolerance, reduction in oxidative stress. Improved plant growth and nodulation | [332] |
| ATPS | ATP sulfurylase | A. thaliana L. | M. sativa L. | Increased tolerance towards a mixture of Cd, Ni, W, Cu, and Pb. Enhanced metal uptake and accumulation in roots and shoots | [322] |
5.2. Transgenesis of Rhizobia for Improving Phytoremediation in Symbiosis with Legumes
| Gene(s) | Genome Function | Gene Origin | Gene Host | Legume Partner | Effect Comparing to the Wild-Type (WT) Control | References |
|---|---|---|---|---|---|---|
| MTL4 | Metallothionein | Arabidopsis thaliana L. | Mesorhizobium huakuii subsp. rengei B3 | Astragalus sinicus Thunb. | Increased Cd accumulation in free-living cells and nodules | [337] |
| PCs | Phytochelatin synthase | A. thaliana L. | Mesorhizobium huakuii subsp. rengei B3 | A. sinicus Thunb. | Increased Cd accumulation in free-living cells and nodules | [338] |
| MTL4 and/or AtPCS | Metallothionein Phytochelatin synthase | A. thaliana L. | Mesorhizobium huakuii subsp. rengei B3 | A. sinicus Thunb. | Increased Cd accumulation in free-living cells and root nodules in hydroponic culture. Additive effect in the case of expression of both genes. Increased Cd accumulation in nodules and roots of A. sinicus in rice paddy soil | [339] |
| PsMT1 | Metallothionein | Pisum sativum L. | Rhizobium leguminosarum bv. viciae | Pisum sativum SGE (WT) or Pisum sativum SGECdt (mutant with increased tolerance to Cd) | Positive effect on WT plant biomass during growth in the presence of Cd. Reduction in the Cd content in shoots of both WT and mutant plants, proper organization of nodules | [340] |
| PSMT2 | Metallothionein | P. sativum L. | R. leguminosarum bv. viciae | Pisum sativum SGE (WT) or P. sativum SGECdt (mutant with increased tolerance to Cd) | Increased Cd accumulation in mutant plants | [340] |
| copAB | Cu+-ATPase | Pseudomonas fluorescens | Ensifer medicae MA11 | Medicago truncatula L. | Alleviation of the toxic effect of Cu in plants. Increased plant growth, nodule numbers, nitrogen contents, and photosynthetic efficiency. Increased Cu accumulation in nodules and roots with decreased accumulation in shoots | [341] |
| copAB | Copper resistance proteins; Cu+-ATPase | P. fluorescens | E. medicae MA11 | M. truncatula composite mutant expressing mt4a gene coding metallothionein | Enhanced Cu tolerance in plants. Improved Cu accumulation in roots, diminished Cu translocation from roots to shoots | [332] |
| CrarsM | S-adenosyl methionine methyl- transferase | Chlamydomonas reinhardtii | Rhizobium leguminosarum bv. trifolii R3 | Trifolium pratense L. | Capability to methylate As by free-living bacteria and in symbiosis with plants. Methylated As forms detected both in nodules and shoots. Lesser amounts of methylated As species were volatilized. | [343] |
5.3. Other Genetic Engineering Methods to Be Used for Improving Bioremediation in Legumes
6. Conclusions and Further Perspective
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ABA | Abscisic acid |
| ABC | ATP-binding cassette |
| ACC | 1-aminocyclopropane 1-carboxylic acid |
| AMPs | Anti-microbial peptides |
| APX | Ascorbate peroxidase |
| ATPS | ATP-sulfurylase |
| C-GCS | c-glutamyl cysteine synthetase |
| CCs | Cover crops |
| CDF | Cation diffusion facilitator |
| EPS | Exopolysaccharides |
| EPS-1 | Polysaccharide succinoglycan |
| GHG | Greenhouse gases |
| GMO | Genetically modified organism |
| GSH | Glutathione |
| GST | Glutathione S-transferase |
| HM | Heavy metal |
| IAA | Indole-3-acetic acid |
| IAM | Indole-3-acetamide |
| IPyA | Indole-3-pyruvic acid |
| MBPs | metal-binding proteins |
| HM | Heavy metal |
| MT | Metallothionein |
| MTP | Metal tolerance proteins |
| NFB | Nitrogen fixating bacteria |
| NRAMP | Natural resistant associated macrophage protein |
| NRE | Non-rhizobium endophytes |
| PC | Phytochelatin |
| PCS | Phytochelatin synthetase |
| PGP | Plant growth-promoting |
| PGPB | Beneficial plant growth-promoting rhizobacteria |
| ROS | Reactive oxygen species |
| SOD | Superoxide dismutase |
| TCA | Tricarboxylic acid |
| TCE | Trichloroethylene |
| Tra | Tryptamine |
| WT | Wild-type |
| ZIP | Zinc–iron permease |
References
- Lewis, G.P.; Schrire, B.; Mackinder, B.; Lock, M. Legumes of the World; Royal Botanic Gardens: Kew, UK, 2005. [Google Scholar]
- Voisin, A.S.; Guéguen, J.; Huyghe, C.; Jeuffroy, M.H.; Magrini, M.B.; Meynard, J.M.; Mougel, C.; Pellerin, S.; Pelzer, E. Legumes for feed, food, biomaterials and bioenergy in Europe: A review. Agron. Sustain. Dev. 2014, 34, 361–380. [Google Scholar] [CrossRef]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Semba, R.D.; Ramsing, R.; Rahman, N.; Kraemer, K.; Bloem, M.W. Legumes as a sustainable source of protein in human diets. Glob. Food Sec. 2021, 28, 100520. [Google Scholar] [CrossRef]
- Maphosa, Y.; Jideani, V.A. The Role of Legumes in Human Nutrition. In Functional Food-Improve Health through Adequate Food; Hueda, M.V., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- Tharanathan, R.N.; Mahadevamma, S. Grain legumes—A boon to human nutrition. Trends Food Sci. Technol. 2003, 14, 507–518. [Google Scholar] [CrossRef]
- Zander, P.; Amjath-Babu, T.S.; Preissel, S.; Reckling, M.; Bues, A.; Schläfke, N.; Kuhlman, T.; Bachinger, J.; Uthes, S.; Stoddard, F.; et al. Grain legume decline and potential recovery in European agriculture: A review. Agron. Sustain. Dev. 2016, 36, 26. [Google Scholar] [CrossRef]
- Duranti, M. Grain legume proteins and nutraceutical properties. Fitoterapia 2006, 77, 67–82. [Google Scholar] [CrossRef]
- Butkutė, B.; Benetis, R.; Padarauskas, A.; Cesevičienė, J.; Dagilytė, A.; Taujenis, L.; Rodovičius, H.; Lemežienė, N. Young herbaceous legumes—A natural reserve of bioactive compounds and antioxidants for healthy food and supplements. J. Appl. Bot. Food Qual. 2017, 90, 346–353. [Google Scholar]
- Budhwar, S.; Chakraborty, M. Novel Dietary and Nutraceutical Supplements from Legumes. Sustain. Agro. Rev. 2020, 45, 53–70. [Google Scholar]
- Kocira, A.; Staniak, M.; Tomaszewska, M.; Kornas, R.; Cymerman, J.; Panasiewicz, K.; Lipińska, H. Legume Cover Crops as One of the Elements of Strategic Weed Management and Soil Quality Improvement. A Review. Agriculture 2020, 10, 394. [Google Scholar] [CrossRef]
- White, P.M.; Williams, G.; Viator, H.P.; Viator, R.P.; Webber, C.L. Legume Cover Crop Effects on Temperate Sugarcane Yields and Their Decomposition in Soil. Agronomy 2020, 10, 703. [Google Scholar] [CrossRef]
- Sharma, P.; Singh, A.; Kachlon, C.S.; Brar, A.S.; Grover, K.K.; Dia, M.; Steiner, R.L. The Role of Cover Crops towards Sustainable Soil Health and Agriculture—A Review Paper. Am. J. Plant Sci. 2018, 9, 1935–1951. [Google Scholar] [CrossRef] [Green Version]
- Meena, B.L.; Fagodiya, R.K.; Prajapat, K.; Dotaniya, M.L.; Kaledhonkar, M.J.; Sharma, P.C.; Meena, R.S.; Mitran, T.; Kumar, S. Legume Green Manuring: An Option for Soil Sustainability. In Legumes for Soil Health and Sustainable Management; Meena, R., Das, A., Yadav, G., Lal, R., Eds.; Springer Nature: Singapore, 2018; Volume 1, pp. 387–408. [Google Scholar]
- Kakraliya, S.K.; Singh, U.; Bohar, A.; Choudhary, K.K.; Kumar, S.; Meena, R.S.; Jat, M.L. Nitrogen and Legumes: A Meta-analysis. In Legumes for Soil Health and Sustainable Management; Meena, R.S., Das, A., Yadav, G.S., Lal, R., Eds.; Springer Nature: Cham, Switzerland, 2018; pp. 277–314. [Google Scholar]
- Velázquez, E.; García-Fraile, P.; Ramírez-Bahena, M.-H.; Rivas, R.; Martínez-Molina, E. Bacteria Involved in Nitrogen-Fixing Legume Symbiosis: Current Taxonomic Perspective. In Microbes for Legume Improvement; Khan, M.S., Musarrat, J., Zaidi, A., Eds.; Springer: Vienna, Austria, 2010; Volume 1, pp. 1–25. [Google Scholar]
- Reinprecht, Y.; Schram, L.; Marsolais, F.; Smith, T.H.; Hill, B.; Pauls, K.P. Effects of Nitrogen application on nitrogen fixation in common bean production. Front. Plant Sci. 2020, 11, 1172. [Google Scholar] [CrossRef] [PubMed]
- Preissel, S.; Reckling, R.; Schläfke, N.; Zander, P. Magnitude and farm-economic value of grain legume pre-crop benefits in Europe: A review. Field Crops Res. 2015, 175, 64–79. [Google Scholar] [CrossRef] [Green Version]
- Carranca, C.; Torres, M.O.; Madeira, M. Underestimated role of legume roots for soil N fertility. Agron. Sustain. Dev. 2015, 35, 1095–1102. [Google Scholar] [CrossRef] [Green Version]
- Blanco-Canqui, H.; Shaver, T.M.; Lindquist, J.L.; Shapiro, C.A.; Elmore, R.W.; Francis, C.A.; Hergert, G.W. Cover crops and ecosystem services: Insights from studies in temperate soils. Agron. J. 2015, 107, 2449–2474. [Google Scholar] [CrossRef] [Green Version]
- Mirsky, S.B.; Curran, W.S.; Mortensen, D.A.; Ryan, M.R.; Shumway, D.L. Timing of Cover-Crop Management Effects on weed suppression in no-till planted soybean using a roller crimper. Weed Sci. 2011, 59, 380–389. [Google Scholar] [CrossRef]
- Elsalahy, H.; Döring, T.; Bellingrath-Kimura, S.; Arends, D. Weed Suppression in Only-Legume Cover Crop Mixtures. Agronomy 2019, 9, 648. [Google Scholar] [CrossRef] [Green Version]
- Köpke, U.; Nemecek, T. Ecological services of faba bean. Field Crops Res. 2010, 115, 217–233. [Google Scholar] [CrossRef]
- Jensen, E.S.; Peoples, M.B.; Boddey, R.M.; Gresshoff, P.M.; Hauggaard-Nielsen, H.; Alves, B.J.; Morrison, M.J. Legumes for mitigation of climate change and the provision of feedstock for biofuels and biorefineries. A review. Agron. Sustain. Dev. 2012, 32, 329–364. [Google Scholar] [CrossRef] [Green Version]
- Jeuffroy, M.H.; Baranger, E.; Carrouée, B.; Chezelles, E.D.; Gosme, M.; Hénault, C.; Schneider, A.; Cellier, P. Nitrous oxide emissions from crop rotations including wheat, oilseed rape and dry peas. Biogeosciences 2013, 10, 1787–1797. [Google Scholar] [CrossRef] [Green Version]
- Abdu, N.; Abdullahi, A.A. Review of heavy metal remediation of contaminated soils (ii): Legume-rhizobia symbiosis, mycorrhiza and other microbes: A review. Niger. J. Sci. 2017, 16, 775–780. [Google Scholar]
- Gerhardt, K.E.; Huang, X.-D.; Glick, B.R.; Greenberg, B.M. Phytoremediation and rhizoremediation of organics oil contaminants: Potential and challenges. Plant Sci. J. 2009, 176, 20–30. [Google Scholar] [CrossRef]
- Tóth, G.; Hermann, T.; Szatmári, G.; Pásztor, L. Maps of heavy metals in the soils of the European Union and proposed priority areas for detailed assessment. Sci. Total Environ. 2016, 565, 1054–1062. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Shu, W.; Lan, C.; Wong, M. Soil seed bank as an input of seed source in revegetation of lead/zinc mine tailings. Restor. Ecol. 2001, 9, 378–385. [Google Scholar] [CrossRef]
- Manoj, S.R.; Karthik, C.; Kadirvelu, K.; Arulselvi, P.I.; Shanmugasundaram, T.; Bruno, B.; Rajkumar, M. Understanding the molecular mechanisms for the enhanced phytoremediation of heavy metals through plant growth promoting rhizobacteria: A review. J. Environ. Manag. 2020, 254, 109779. [Google Scholar] [CrossRef]
- Tóth, G.; Hermann, T.; Da Silvac, M.R.; Montanarella, L. Heavy metals in agricultural soils of the European Union with implications for food safety. Environ. Int. 2016, 88, 299–309. [Google Scholar] [CrossRef]
- Mandal, S.M.; Bhattacharyya, R. Rhizobium–Legume Symbiosis: A Model System for the Recovery of Metal-Contaminated Agricultural Land. In Toxicity of Heavy Metals to Legumes and Bioremediation; Zaidi, A., Wani, P., Khan, M., Eds.; Springer: Vienna, Austria, 2012; Volume 1, pp. 115–127. [Google Scholar]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals—concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef]
- Neilson, S.; Rajakaruna, N. Phytoremediation of Agricultural Soils: Using Plants to Clean Metal-Contaminated Arable Land. In Phytoremediation; Ansari, A., Gill, S., Gill, R., Lanza, G., Newman, L., Eds.; Springer: Cham, Denmark, 2015; Volume 1, pp. 159–168. [Google Scholar]
- Gómez-Sagasti, M.T.; Marino, D. PGPRs and nitrogen-fixing legumes: A perfect team for efficient Cd phytoremediation? Front. Plant Sci. 2015, 6, 81. [Google Scholar]
- Pajuelo, E.; Dary, M.; Palomares, A.J.; Rodriguez-Llorente, I.D.; Carrasco, J.A.; Chamber, M.A. Biorhizoremediation of heavy metals toxicity using rhizobium-legume symbioses. In Biological Nitrogen Fixation: Towards Poverty Alleviation through Sustainable Agriculture. Current Plant Science and Biotechnology in Agriculture; Dakora, F.D., Chimphango, S.B.M., Valentine, A.J., Elmerich, C., Newton, W.E., Eds.; Springer: Dordrecht, The Netherlands, 2008; Volume 42, pp. 101–104. [Google Scholar]
- Lebrazi, S.; Fikri-Benbrahim, K. Rhizobium-legume symbioses: Heavy metal effects and principal approaches for bioremediation of contaminated soil. In Legumes for Soil Health and Sustainable Management; Meena, R., Das, A., Yadav, G., Lal, R., Eds.; Springer: Heidelberg, Germany, 2018; Volume 1, pp. 205–233. [Google Scholar]
- Fagorzi, C.; Checcucci, A.; DiCenzo, G.C.; Debiec-Andrzejewska, K.; Dziewit, Ł.; Pini, F.; Mengoni, A. Harnessing Rhizobia to Improve Heavy-Metal Phytoremediation by Legumes. Genes 2018, 9, 542. [Google Scholar] [CrossRef] [Green Version]
- Bradl, H.B. Heavy Metals in the Environment: Origin, Interaction and Remediation; Elsevier: London, UK, 2005. [Google Scholar]
- Luo, H.S.; Xue, Y.; Wang, Y.L.; Cang, L.; Xu, B.; Ding, J. Source identification and apportionment of heavy metals in urban soil profiles. Chemosphere 2015, 127, 152–157. [Google Scholar] [CrossRef]
- Singh, U.K.; Kumar, B. Pathways of heavy metals contamination and associated human health risk in Ajay River basin, India. Chemosphere 2017, 174, 183–199. [Google Scholar] [CrossRef] [PubMed]
- Masindi, V.; Muedi, K.L. Environmental Contamination by Heavy Metals. In Heavy Metals; Hosam El-Din, M., Aglam, S., Aglam, R.F., Eds.; IntechOpen: London, UK, 2018; Volume 1, pp. 115–132. [Google Scholar]
- Dudka, S.; Miller, W.P. Accumulation of potentially toxic elements in plants and their transfer to human food chain. J. Environ. Sci. Health B 1999, 34, 681–708. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.S.; Kapri, A.; Goel, R. Heavy metal pollution: Source, impact, and remedies. In Biomanagement of Metal-Contaminated Soils. Environmental Pollution; Khan, M., Zaidi, A., Goel, R., Musarrat, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 1, pp. 1–28. [Google Scholar]
- Pujari, M.; Kapoor, D. Heavy metals in the ecosystem: Sources and their effects. In Heavy Metals in the Environment; Kumar, V., Sharma, A., Cerdà, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2021; Volume 1, pp. 1–7. [Google Scholar]
- Friedlová, M. The influence of heavy metals on soil biological and chemical properties. Soil Water Res. 2010, 5, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Nagajyoti, P.C.; Lee, K.D.; Sreekanth, T.V.M. Heavy metals, occurrence and toxicity for plants: A review. Environ. Chem. Lett. 2010, 8, 199–216. [Google Scholar] [CrossRef]
- Arora, S.; Jain, C.K.; Lokhande, R.S. Review of Heavy Metal Contamination in Soil. Int. J. Environ. Sci. Nat. Res. 2017, 3, 139–144. [Google Scholar] [CrossRef]
- Crowley, D. Impacts of Metals and Metalloids on Soil Microbial Diversity and Ecosystem Function. In Proceedings of the 5th International Symposium ISMOM, Pucón, Chile, 24–28 November 2008; pp. 1–7. [Google Scholar]
- Chu, D. Effects of heavy metals on soil microbial community. IOP Conf. Ser. Earth Environ. Sci. 2018, 113, 012009. [Google Scholar] [CrossRef]
- Angelovičová, L.; Bobulská, L.; Fazekašová, D. Toxicity of heavy metals to soil biological and chemical properties in conditions of environmentally polluted area middle Spiš (Slovakia). Carpathian J. Earth Environ. Sci. 2015, 10, 193–201. [Google Scholar]
- Wang, Y.P.; Shi, J.Y.; Wang, H.; Lin, Q.; Chen, X.C.; Chen, Y.X. The influence of soil heavy metals pollution on soil microbial biomass, enzyme activity, and community composition near a copper smelter. Ecotoxicol. Environ. Saf. 2007, 67, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Järup, J. Hazards of heavy metal contamination. Br. Med. Bull. 2003, 68, 167–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morais, S.; Costa, F.G.; de Lourdes Pereira, M. Heavy metals and human health. In Environmental Health—Emerging Issues and Practice; Oosthuizen, J., Ed.; InTechOpen: Rijeka, Croatia, 2012; Volume 1, pp. 227–246. [Google Scholar]
- Rai, P.K.; Lee, S.S.; Zhang, M.; Tsang, Y.F.; Kim, K.-H. Heavy metals in food crops: Health risks, fate, mechanisms, and management. Environ. Int. 2019, 25, 365–385. [Google Scholar] [CrossRef] [PubMed]
- Alengebawy, A.; Abdelkhalek, S.T.; Qureshi, S.R.; Wang, M.-G. Heavy Metals and Pesticides Toxicity in Agricultural Soil and Plants: Ecological Risks and Human Health Implications. Toxics 2021, 9, 42. [Google Scholar] [CrossRef] [PubMed]
- Asati, A.; Pichhode, M.; Nikhil, K. Effect of Heavy Metals on Plants: An Overview. Int. J. Appl. Innov. Eng. Manag. 2016, 5, 56–66. [Google Scholar]
- El-Kady, A.A.; Abdel-Wahhab, M.A. Occurrence of trace metals in foodstuffs and their health impact. Trends Food Sci. Technol. 2018, 75, 36–45. [Google Scholar] [CrossRef]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e04691. [Google Scholar] [CrossRef]
- Wang, S.; Shi, X. Molecular mechanisms of metal toxicity and carcinogenesis. Mol. Cell. Biochem. 2001, 222, 3–9. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjon, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy Metal Toxicity and Environment. In Molecular, Clinical and Environmental Toxicology; Luch, A., Ed.; Springer: Basel, Switzerland, 2012; pp. 133–164. [Google Scholar]
- Chibuike, G.U.; Obiora, S.C. Heavy Metal Polluted Soils: Effect on Plants and Bioremediation Methods. Appl. Environ. Soil Sci. 2014, 2014, 752708. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rinklebe, J.; Tack, F.M.G.; Hou, D. A review of green remediation strategies for heavy metal contaminated soil. Soil Use Manag. 2021, 27, 936–963. [Google Scholar] [CrossRef]
- Mosa, K.A.; Saadoun, I.; Kumar, K.; Helmy, M.; Dhankher, O.P. Potential Biotechnological Strategies for the Cleanup of Heavy Metals and Metalloids. Front. Plant Sci. 2016, 7, 303. [Google Scholar] [CrossRef] [Green Version]
- Chandra, R.; Saxena, G.; Kumar, V. Phytoremediation of Environmental Pollutants: An Eco-Sustainable Green Technology to Environmental Management. In Advances in Biodegradation and Bioremediation of Industrial Waste; Chandra, R., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 1–29. [Google Scholar]
- Ullah, A.; Heng, S.; Munis, M.F.H.; Fahad, S.; Yang, X. Phytoremediation of heavy metals assisted by plant growth promoting (PGP) bacteria: A review. Environ. Exp. Bot. 2015, 117, 28–40. [Google Scholar] [CrossRef]
- Rengel, Z. Heavy Metals as Essential Nutrients. In Heavy Metal Stress in Plants; Prasad, M.N.V., Ed.; Springer: Berlin/Heidelberg, Germany, 1999; Volume 1, pp. 271–294. [Google Scholar]
- Wuana, R.A.; Okieimen, F.E. Heavy Metals in Contaminated Soils: A Review of Sources, Chemistry, Risks and Best Available Strategies for Remediation. ISRN Ecol. 2011, 2011, 402647. [Google Scholar] [CrossRef] [Green Version]
- Peralta-Videa, J.R.; Lopez, M.L.; Narayan, M.; Saupe, G.; Gardea-Torresdey, J. The biochemistry of environmental heavy metal uptake by plants: Implications for the food chain. Int. J. Biochem. Cell Biol. 2009, 41, 1665–1677. [Google Scholar] [CrossRef] [PubMed]
- Arif, N.; Yadav, V.; Singh, S.; Singh, S.; Ahmad, P.; Mishra, R.K.; Sharma, S.; Tripathi, D.K.; Dubey, N.K.; Chauhan, D.K. Influence of High and Low Levels of Plant-Beneficial Heavy Metal Ions on Plant Growth and Development. Front. Environ. Sci. 2016, 4, 69. [Google Scholar] [CrossRef]
- Oves, M.; Khan, S.; Qari, H.; Felemban, N.; Almeelbi, T. Heavy Metals: Biological Importance and Detoxification Strategies. J. Bioremed. Biodegrad. 2016, 7, 1–15. [Google Scholar]
- Valko, M.; Morris, H.; Cronin, M.T.D. Metals, Toxicity and Oxidative Stress. Curr. Med. Chem. 2005, 12, 1161–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Cobbina, S.J.; Mao, G.; Xu, H.; Zhang, Z.; Yang, L. A review of toxicity and mechanisms of individual and mixtures of heavy metals in the environment. Environ. Sci. Pollut. Res. Int. 2016, 23, 8244–8259. [Google Scholar] [CrossRef]
- Li, H.F.; Gray, C.; Mico, C.; Zhao, F.J.; McGrath, S.P. Phytotoxicity and bioavailability of cobalt to plants in a range of soils. Chemosphere 2009, 75, 979–986. [Google Scholar] [CrossRef]
- Shah, F.U.R.; Ahmad, N.; Masood, K.R.; Peralta-Videa, J.R.; Ahmad, F.D. Heavy Metal Toxicity in Plants. In Plant Adaptation and PhytoremediaKhantion; Ashraf, M., Ozturk, M., Ahmad, M., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 71–97. [Google Scholar]
- Khan, M.R.; Khan, M.M. Effect of Varying Concentration of Nickel and Cobalt on the Plant Growth and Yield of Chickpea. Aust. J. Basic Appl. Sci. 2010, 4, 1036–1046. [Google Scholar]
- Yadav, S. Heavy metals toxicity in plants: An overview on the role of glutathione and phytochelatins in heavy metal stress tolerance of plants. S. Afr. J. Bot. 2010, 76, 167–179. [Google Scholar] [CrossRef] [Green Version]
- Jibril, S.A.; Hassan, S.A.; Ishak, C.F.; Megat Wahab, P.E. Cadmium Toxicity Affects Phytochemicals and Nutrient Elements Composition of Lettuce (Lactuca sativa L.). Adv. Agric. 2017, 2017, 1236830. [Google Scholar] [CrossRef] [Green Version]
- Nascimento, V.; Nogueira, G.; Monteiro, G.; Júnior, W.; Numes de Freitas, J.M.; Neto, C. Influence of Heavy Metals on the Nitrogen Metabolism in Plants. In Nitrogen in Agriculture; Ohyana, T., Ed.; IntechOpen: London, UK, 2021; Volume 1, pp. 120–197. [Google Scholar]
- Hayat, M.T.; Nauman, M.; Nazir, N.; Ali, S.; Bangash, N. Environmental Hazards of Cadmium: Past, Present, and Future. In Cadmium Toxicity and Tolerance in Plants: From Physiology to Remediation; Hasanuzzaman, M., Prasad, M.N.V., Fujita, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 163–183. [Google Scholar]
- Nazar, R.; Iqbal, N.; Masood, A.; Khan, M.I.R.; Syeed, S.; Khan, N.A. Cadmium Toxicity in Plants and Role of Mineral Nutrients in Its Alleviation. Am. J. Plant Sci. 2012, 3, 1476–1489. [Google Scholar] [CrossRef] [Green Version]
- Rout, J.R.; Ram, S.S.; Das, R.; Chakraborty, A.; Sudarshan, M.; Sahoo, S.L. Copper-stress induced alterations in protein profile and antioxidant enzymes activities in the in vitro grown Withania somnifera L. Physiol. Mol. Biol. Plants 2013, 19, 353–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, J.C.; Henriques, F.S. Biochemical, physiological, and structural effects of excess copper in plants. Bot. Rev. 1991, 57, 246–273. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Zia-ur-Rehman, M.; Irshad, M.K.; Bharwana, S.A. The effect of excess copper on growth and physiology of important food crops: A review. Environ. Sci. Pollut. Res. 2015, 22, 8148–8162. [Google Scholar] [CrossRef] [PubMed]
- Shakya, K.; Chettri, M.K.; Sawidis, T. Impact of Heavy Metals (Copper, Zinc, and Lead) on the Chlorophyll Content of Some Mosses. Arch. Environ. Contam. Toxicol. 2008, 54, 412–421. [Google Scholar] [CrossRef] [PubMed]
- Manivasagaperumal, R.; Vijayarengan, P.; Balamurugan, S.; Thiyagarajan, G. Effect of copper on growth, dry matter yield and nutrient content of Vigna radiata (L.) Wilczek. J. Phytol. 2011, 3, 53–62. [Google Scholar]
- Mukhopadhyay, M.; Das, A.; Subba, P.; Bantawa, P.; Sarkar, B.; Ghosh, P.; Mondal, T.K. Structural, physiological, and biochemical profiling of tea plantlets under zinc stress. Biol. Plant. 2013, 57, 474–480. [Google Scholar] [CrossRef]
- Rout, G.; Das, P. Effect of Metal Toxicity on Plant Growth and Metabolism: I. Zinc. Agron. EDP Sci. 2003, 23, 3–11. [Google Scholar] [CrossRef]
- Warne, M.S.J.; Heemsbergen, D.; Stevens, D.; McLaughlin, M.; Cozens, G.; Whatmuff, M.; Broos, K.; Barry, G.; Bell, M.; Nash, D.; et al. Modeling the toxicity of copper and zinc salts to wheat in 14 soils. Envirol. Toxicol. Chem. 2008, 27, 786–792. [Google Scholar] [CrossRef]
- Kaur, H.; Garg, N. Zinc toxicity in plants: A review. Planta 2021, 253, 129. [Google Scholar] [CrossRef]
- Abedin, M.J.; Cotter-Howells, J.; Meharg, A.A. Arsenic uptake and accumulation in rice (Oryza sativa L.) irrigated with contaminated water. Plant Soil 2002, 240, 311–319. [Google Scholar] [CrossRef]
- Finnegan, P.M.; Chen, W. Arsenic Toxicity: The Effects on Plant Metabolism. Front. Physiol. 2012, 3, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbas, G.; Murtaza, B.; Bibi, I.; Shahid, M.; Niazi, N.K.; Khan, M.I.; Amjad, M.; Hussain, M.; Natasha, N. Arsenic Uptake, Toxicity, Detoxification, and Speciation in Plants: Physiological, Biochemical, and Molecular Aspects. Int. J. Environ. Res. Public Health 2018, 15, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armendariz, A.L.; Talano, M.A.; Travaglia, C.; Reinoso, H.; Oller, A.L.W.; Agostini, E. Arsenic toxicity in soybean seedlings and their attenuation mechanisms. Plant Physiol. Biochem. 2016, 98, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Ghasemi, F.; Heidari, R.; Jameii, R.; Purakbar, L. Effects of Ni2+ toxicity on Hill reaction and membrane functionality in maize. J. Stress Physiol. Biochem. 2012, 8, 55–61. [Google Scholar]
- Rahman, H.; Sabreen, S.; Alam, S.; Kawai, S. Effects of nickel on growth and composition of metal micronutrients in barley plants grown in nutrient solution. J. Plant Nutr. 2005, 28, 393–404. [Google Scholar] [CrossRef]
- Bhalerao, S.A.; Sharma, A.S.; Poojar, A.C. Toxicity of Nickel in Plants. Int. J. Pure Appl. Biosci. 2015, 3, 345–355. [Google Scholar]
- Lan, M.M.; Khan, C.; Liu, S.J.; Qiu, R.L.; Tang, Y.T. Phytostabilization of cd and pb in highly polluted farmland soils using ramie and amendments. Int. J. Environ. Res. Public Health 2020, 17, 1661. [Google Scholar] [CrossRef] [Green Version]
- Kushwaha, A.; Hans, N.; Kumar, S.; Rani, R. A critical review on speciation, mobilization and toxicity of lead in soil-microbe-plant system and bioremediation strategies. Ecotoxicol. Environ. Saf. 2018, 147, 1035–1045. [Google Scholar] [CrossRef]
- Ali, M.; Nas, F.S. The effect of lead on plants in terms of growing and biochemical parameters: A review. MOJ Ecol. Environ. Sci. 2018, 3, 265–268. [Google Scholar] [CrossRef]
- Anjum, S.A.; Askraf, U.; Khan, I.; Tanverr, M.; Shahid, M.; Shakoor, A.; Wang, L. Phyto-Toxicity of Chromium in Maize: Oxidative Damage, Osmolyte Accumulation, Anti-Oxidative Defense and Chromium Uptake. Pedosphere 2017, 27, 262–273. [Google Scholar] [CrossRef]
- Scoccianti, V.; Crinelli, R.; Tirillini, B.; Mancinelli, V.; Speranza, A. Uptake and toxicity of Cr (III) in celery seedlings. Chemosphere 2006, 64, 1695–1703. [Google Scholar] [CrossRef]
- Stambulska, U.Y.; Baylijak, M.M.; Lushchak, V.I. Chromium(VI) Toxicity in Legume Plants: Modulation Effects of Rhizobial Symbiosis. BioMed Res. Int. 2018, 2018, 8031213. [Google Scholar] [CrossRef]
- Hu, X.; Wei, X.; Ling, J.; Chen, J. Cobalt: An Essential Micronutrient for Plant Growth? Front. Plant Sci. 2021, 12, 768523. [Google Scholar] [CrossRef]
- Mahey, S.; Kumar, R.; Sharma, M.; Kumar, V.; Bhardwaj, R. A critical review on toxicity of cobalt and its bioremediation strategies. SN Appl. Sci. 2020, 2, 1279. [Google Scholar] [CrossRef]
- de Oliveira Jucoski, G.; Cambraia, J.; Ribeiro, C.; de Oliveira, J.A.; de Paula, S.O.; Oliva, M.A. Impact of iron toxicity on oxidative metabolism in young Eugenia uniflora L. plants. Acta Physiol. Plant 2013, 35, 1645–1657. [Google Scholar] [CrossRef]
- Chatterjee, C.; Gopal, R.; Dube, B.K. Impact of iron stress on biomass, yield, metabolism and quality of potato (Solanum tuberosum L.). Sci. Hortic. 2006, 108, 1–6. [Google Scholar] [CrossRef]
- Kleiber, T.; Graje, K.M. Tomato reaction on excessive manganese nutrition. Bulg. J. Agric. Sci. 2015, 21, 118–125. [Google Scholar]
- Kovácik, J.; Babula, P.; Hedbavny, J.; Švec, P. Manganese-induced oxidative stress in two ontogenetic stages of chamomile and amelioration by nitric oxide. Plant Sci. 2014, 215, 1–10. [Google Scholar] [CrossRef]
- McLaughlin, M.J.; Smolders, E.; Degryse, F.; Rietra, R. Uptake of Metals from Soil into Vegetables. In Dealing with Contaminated Sites; Swartjes, F., Ed.; Springer: Dordrecht, The Netherlands, 2011; pp. 325–367. [Google Scholar]
- Cobbett, C.; Goldsbrought, P. Phytochelatins and Metallothioneins: Roles in Heavy Metal Detoxification and Homeostasis. Annu. Rev. Plant Biol. 2002, 53, 159–182. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Dalvi, A.A.; Bhalerao, S.A. Response of Plants towards Heavy Metal Toxicity: An overview of Avoidance, Tolerance and Uptake Mechanism. Ann. Plant Sci. 2013, 2, 362–368. [Google Scholar]
- Gupta, D.K.; Vandenhove, H.; Inouhe, M. Role of Phytochelatins in Heavy Metal Stress and Detoxification Mechanisms in Plants. In Heavy Metal Stress in Plants; Gupta, D., Corpas, F., Palma, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 73–94. [Google Scholar]
- Yaashikaa, P.R.; Senthil Kumar, P.; Jeevanantham, S.; Saravanan, R. A review on bioremediation approach for heavy metal detoxification and accumulation in plants. Environ. Pollut. 2022, 301, 119035. [Google Scholar] [CrossRef]
- Sharma, P.; Pandey, K.A.; Udayan, A.; Kumar, S. Role of microbial community and metal-binding proteins in phytoremediation of heavy metals from industrial wastewater. Bioresour. Technol. 2021, 326, 124750. [Google Scholar] [CrossRef]
- Polak, N.; Read, D.S.; Jurkschat, K.; Matzke, M.; Kelly, F.J.; Spurgeon, D.J.; Stürzenbaum, S.R. Metalloproteins and phytochelatin synthase may confer protection against zinc oxide nanoparticle induced toxicity in Caenorhabditis elegans. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2014, 160, 75–85. [Google Scholar] [CrossRef] [Green Version]
- Huang, G.Y.; Wang, Y.S.; Ying, G.G. Cadmium-inducible BgMT2, a type 2 metallothionein gene from mangrove species (Bruguiera gymnorrhiza), its encoding protein shows metal-binding ability. J. Exp. Mar. Biol. Ecol. 2011, 405, 128–132. [Google Scholar] [CrossRef]
- Rodríguez-Llorente, I.D.; Perez-Palacios, P.; Doukkali, B.; Caviedes, M.A.; Pajuelo, E. Expression of the seed-specific metallothionein mt4a in plant vegetative tissues increases Cu and Zn tolerance. Plant Sci. 2010, 178, 327–332. [Google Scholar] [CrossRef]
- Peer, W.A.; Baxter, I.R.; Richards, E.L.; Freeman, J.L.; Murphy, A.S. Phytoremediation and hyperaccumulator plants. In Molecular Biology of Metal Homeostasis and Detoxification; Tamas, M.J., Martinoia, E., Eds.; Springer: Berlin, Germany, 2005; Volume 1, pp. 299–340. [Google Scholar]
- Thakur, S.; Singh, L.; Wahid, Z.A.; Siddiqui, M.F.; Atnaw, S.M.; Din, M.F.M. Plant-driven removal of heavy metals from soil: Uptake, translocation, tolerance mechanism, challenges, and future perspectives. Environ. Monit. Assess. 2016, 188, 206. [Google Scholar] [CrossRef]
- Williams, L.E.; Pittman, J.K.; Hall, J.L. Emerging mechanisms for heavy metal transport in plants. Biochim. Biophys. Acta Bioenerg BBA 2000, 1465, 104–126. [Google Scholar] [CrossRef]
- Singh, S.; Parihar, P.; Singh, R.; Singh, V.P.; Prasad, S.M. Heavy Metal Tolerance in Plants: Role of Transcriptomics, Proteomics, Metabolomics, and Ionomics. Front. Plant Sci. 2016, 6, 1143. [Google Scholar] [CrossRef] [Green Version]
- Yan, A.; Wang, Y.; Tan, S.N.; Mohd Yusof, L.M.; Ghosh, S.; Chen, Z. Phytoremediation: A Promising Approach for Revegetation of Heavy Metal-Polluted Land. Front. Plant Sci. 2020, 11, 359. [Google Scholar] [CrossRef]
- Guerinot, M.L. The ZIP family of metal transporters. Biochim. Biophys. Acta 2000, 1465, 190–198. [Google Scholar] [CrossRef] [Green Version]
- Ajeesh Krishna, T.P.; Maharajan, T.; Roch, G.V.; Savarimuthu, I.; Ceasar, S.A. Structure, Function, Regulation and Phylogenetic Relationship of ZIP Family Transporters of Plants. Front. Plant Sci. 2020, 11, 662. [Google Scholar] [CrossRef]
- Takahashi, R.; Bashir, K.; Ishimaru, Y.; Nishizawa, N.K.; Nakanishi, H. The role of heavy-metal ATPases, HMAs, in zinc and cadmium transport in rice. Plant Signal. Behav. 2012, 7, 1605–1607. [Google Scholar] [CrossRef]
- Nevo, Y.; Nelson, N. The NRAMP family of metal-ion transporters. Biochim. Biophys. Acta Mol. Cell Res. 2006, 1763, 609–620. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Li, G.; Yang, J.; Zhao, X.; Sun, Z.; Hou, H. Role of Nramp transporter genes of Spirodela polyrhiza in cadmium accumulation. Ecotoxicol. Environ. Saf. 2021, 227, 112907. [Google Scholar] [CrossRef]
- Gustin, J.L.; Zanis, M.J.; Salt, D.E. Structure and evolution of the plant cation diffusion facilitator family of ion transporters. BMC Evol. Biol. 2011, 11, 76. [Google Scholar] [CrossRef] [Green Version]
- Ricachenevsky, F.K.; Menguer, P.K.; Sperotto, R.A.; Williams, L.E.; Fett, J.P. Roles of plant metal tolerance proteins (MTP) in metal storage and potential use in biofortification strategies. Front. Plant Sci. 2013, 4, 144. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.-U.; Song, W.-Y.; Hong, D.; Ko, D.; Yamaoka, Y.; Jang, S.; Yim, S.; Lee, E.; Khare, D.; Kim, K.; et al. Plant ABC Transporters enable many unique aspects of a terrestrial plant’s lifestyle. Mol. Plant 2016, 9, 338–355. [Google Scholar] [CrossRef] [Green Version]
- Pajuelo, E.; Rodríguez-Llorente, I.D.; Lafuente, A.; Caviedes, M.Á. Legume–Rhizobium Symbioses as a Tool for Bioremediation of Heavy Metal Polluted Soils. In Biomanagement of Metal-Contaminated Soils. Environmental Pollution; Khan, M., Zaidi, A., Goel, R., Musarrat, J., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 20, pp. 95–123. [Google Scholar]
- Rai, K.K.; Pandey, N.; Meena, R.P.; Rai, S.P. Biotechnological strategies for enhancing heavy metal tolerance in neglected and underutilized legume crops: A comprehensive review. Ecotoxicol. Environ. Saf. 2021, 208, 111750. [Google Scholar] [CrossRef]
- Nedjimi, B. Phytoremediation: A sustainable environmental technology for heavy metals decontamination. SN Appl. Sci. 2021, 3, 286. [Google Scholar] [CrossRef]
- Mohamad, R.; Maynaud, G.; Le Quéré, A.; Vidal, C.; Klonowska, A.; Yashiro, E.; Cleyet-Marel, J.-C.; Brunel, B. Ancient Heavy Metal Contamination in Soils as a Driver of Tolerant Anthyllis vulneraria Rhizobial Communities. Appl. Environ. Microbiol. 2017, 83, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sujkowska-Rybkowska, M.; Ważny, R. Metal resistant rhizobia and ultrastructure of Anthyllis vulneraria nodules from zinc and lead contaminated tailing in Poland. Int. J. Phytoremediation 2018, 20, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Gratão, P.L.; Vara Prasad, M.N.; Cardoso, P.F.; Lea, P.J.; Azevedo, R.A. Phytoremediation: Green technology for the clean up of toxic metals in the environment. Braz. J. Plant Physiol. 2005, 17, 53–65. [Google Scholar] [CrossRef]
- Lago-Vila, M.; Arenas-Lago, D.; Rodríguez-Seijo, A.; Andrade, M.L.; Vega, F.A. Ability of Cytisus scoparius for phytoremediation of soils from a Pb/Zn mine: Assessment of metal bioavailability and bioaccumulation. J. Environ. Manag. 2019, 235, 152–160. [Google Scholar] [CrossRef]
- Guo, J.; Chi, J. Effect of Cd-tolerant plant growth-promoting rhizobium on plant growth and Cd uptake by Lolium multiflorum Lam. and Glycine max (L.) Merr. in Cd-contaminated soil. Plant Soil 2014, 375, 205–214. [Google Scholar] [CrossRef]
- Reichman, S.M. The Potential use of the legume-rhizobium symbiosis for the remediation of arsenic contaminated sites. Soil Biol. Biochem. 2007, 39, 2587–2593. [Google Scholar] [CrossRef]
- Younis, M. Responses of Lablab purpureus–Rhizobium symbiosis to heavy metals in pot and field experiments. World J. Agric. Res. 2007, 3, 111–122. [Google Scholar]
- Brunet, J.; Repellin, A.; Varrault, G.; Terryna, N.; Zuily-Fodila, Y. Lead accumulation in the roots of grass pea (Lathyrus sativus L.): A novel plant for phytoremediation systems? Comptes Rendus Biol. 2008, 331, 859–864. [Google Scholar] [CrossRef]
- Wani, P.; Khan, M.; Zaidi, A. Impact of zinc-tolerant plant growth-promoting rhizobacteria on lentil grown in zinc-amended soil. Agron. Sustain. Dev. 2008, 28, 449–455. [Google Scholar] [CrossRef]
- Ijilmassi, B.; Filali-Maltouf, A.; Boulahyaoui, H.; Kricha, A.; Boubekri, K.; Udupa, S.; Kumar, S.; Amri, A. Assessment of genetic diversity and symbiotic efficiency of selected rhizobia strains nodulating lentil (Lens culinaris Medik.). Plants 2020, 10, 15. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Rusaczonek, A.; Kochańska-Jeziorska, A. Exploring apoplast reorganization in the nodules of Lotus corniculatus L. growing on old Zn-Pb calamine wastes. J. Plant Physiol. 2022, 268, 153561. [Google Scholar] [CrossRef] [PubMed]
- Vázquez, S.; Agha, R.; Granado, A.; Sarro, M.J.; Esteban, E.; Peñalosa, J.M.; Carpena, R.O. Use of White Lupin Plant for Phytostabilization of Cd and As Pollute Acid Soil. Water Air Soil Pollut. 2006, 177, 349–365. [Google Scholar] [CrossRef]
- Fumagalli, P.; Comolli, R.; Ferre, C.; Ghiani, A.; Gentili, R.; Citterio, S. The rotation of white lupin (Lupinus albus L.) with metal-accumulating plant crops: A strategy to increase the benefits of soil phytoremediation. J. Environ. Manag. 2014, 145, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Dary, M.; Chamber-Pérez, M.A.; Palomares, A.J.; Pajuelo, E. “In situ” phytostabilisation of heavy metal polluted soils using Lupinus luteus inoculated with metal resistant plant-growth promoting rhizobacteria. J. Hazard. Mater. 2010, 177, 323–330. [Google Scholar] [CrossRef]
- Kong, Z.; Glick, B.R.; Duan, J.; Ding, S.; Tian, J.; McConkey, B.J.; Wei, G. Effects of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-overproducing Sinorhizobium meliloti on plant growth and copper tolerance of Medicago lupulina. Plant Soil 2015, 391, 383–394. [Google Scholar] [CrossRef]
- Ghnaya, T.; Mnassri, M.; Ghabriche, R.; Wali, M.; Poschenrieder, C.; Lutts, S.; Abdelly, C. Nodulation by Sinorhizobium meliloti originated from a mining soil alleviates Cd toxicity and increases Cd-phytoextraction in Medicago sativa L. Front. Plant Sci. 2015, 6, 863. [Google Scholar] [CrossRef] [Green Version]
- Nonnoi, F.; Chinnaswamy, A.; García de la Torre, V.S.; Coba de la Peña, T.; Lucas, M.M.; Pueyo, J.J. Metal tolerance of rhizobial strains isolated from nodules of herbaceous legumes (Medicago spp. and Trifolium spp.) growing in mercury-contaminated soils. Appl. Soil Ecol. 2012, 61, 49–59. [Google Scholar] [CrossRef] [Green Version]
- Shen, G.; Ju, W.; Liu, Y.; Guo, X.; Zhao, W.; Fang, L. Impact of Urea Addition and Rhizobium Inoculation on Plant Resistance in Metal Contaminated Soil. Int. J. Environ. Res. Public Health 2019, 16, 1955. [Google Scholar] [CrossRef] [Green Version]
- Wani, P.A.; Khan, M.S.; Zaidi, A. Effect of metal-tolerant plant growth-promoting Rhizobium on the performance of pea grown in metal-amended soil. Arch. Environ. Contam. Toxicol. 2008, 55, 33–42. [Google Scholar] [CrossRef]
- Wani, P.A.; Khan, M.S.; Zaidi, A. Effects of heavy metal toxicity on growth, symbiosis, seed yield and metal uptake in pea grown in metal amended soil. Bull. Environ. Contam. Toxicol. 2008, 81, 152–158. [Google Scholar] [CrossRef]
- Muro-Gonzáles, D.A.; Mussali-Galante, P.; Valencia-Cuevas, L.; Flores-Trujillo, K.; Tovar-Sachez, E. Morphological, physiological, and genotoxic effects of heavy metal bioaccumulation in Prosopis laevigata reveal its potential for phytoremediation. Environ. Sci. Pollut. Res. 2020, 27, 40187–40204. [Google Scholar] [CrossRef] [PubMed]
- Băbău, A.M.C.; Micle, V.; Damian, G.E.; Sur, I.M. Sustainable Ecological Restoration of Sterile Dumps Using Robinia pseudoacacia. Sustainability 2021, 13, 14021. [Google Scholar] [CrossRef]
- Dadea, C.; Russo, A.; Tagliavini, M.; Mimmo, T.; Zerbe, S. Tree Species as Tools for Biomonitoring and Phytoremediation in Urban Environments: A Review with Special Regard to Heavy Metals. Arboric. Urban For. 2017, 43, 155–167. [Google Scholar] [CrossRef]
- Yang, B.; Shu, W.; Ye, Z.; Lan, C.; Wong, M. Growth and metal accumulation in vetiver and two Sesbania species on lead/zinc mine tailings. Chemosphere 2003, 52, 1593–1600. [Google Scholar] [CrossRef] [Green Version]
- Saadani, O.; Jebara, S.H.; Fatnassi, I.C.; Chiboub, M.; Mannai, K.; Zarrad, I.; Jebara, M. Effect of Vicia faba L. var. minor and Sulla coronaria (L.) Medik associated with plant growth-promoting bacteria on lettuce cropping system and heavy metal phytoremediation under field conditions. Environ. Sci. Pollut. Res. 2019, 26, 8125–8135. [Google Scholar] [CrossRef]
- Ibañez, S.; Medina, M.; Agostini, E. Vicia: A green bridge to clean up polluted environments. Appl. Microbiol. Biotechnol. 2020, 104, 13–21. [Google Scholar] [CrossRef]
- Gospodarek, J.; Nadgórska–Socha, A. Chemical composition of broad beans (Vicia faba L.) and development parameters of black bean aphid (Aphis fabae Scop.) under conditions of soil contamination with oil derivatives. J. Elem. 2016, 21, 1359–1376. [Google Scholar] [CrossRef]
- Kanwal, A.; Ali, S.; Farhan, M. Heavy metal phytoextraction potential of indigenous tree species of family fabaceae. Int. J. Phytoremediation 2019, 21, 251–258. [Google Scholar] [CrossRef]
- Wong, M.H. Ecological restoration of mine degraded soils, with emphasis on metal contaminated soils. Chemosphere 2003, 50, 775–780. [Google Scholar] [CrossRef]
- Hao, X.; Taghavi, S.; Xie, P.; Orbach, M.J.; Alwathanini, H.A.; Rensing, C.; Wei, G. Phytoremediation of Heavy and Transition Metals Aided by Legume-Rhizobia Symbiosis. Int. J. Phytoremediation 2014, 16, 179–202. [Google Scholar] [CrossRef]
- Renella, G.; Chaudri, A.; Brookes, P. Fresh additions of heavy metals do not model long-term effects on microbial biomass and activity. Soil Biol. Biochem. 2002, 34, 121–124. [Google Scholar] [CrossRef]
- Smolders, E.; McGrath, S.P.; Lombi, E.; Karman, C.C.; Bernhard, R.; Cools, D.; Van den Brande, K.; van Os, B.; Walrave, N. Comparison of toxicity of zinc for soil microbial processes between laboratory-contamined and polluted field soils. Environ. Toxicol. Chem. 2003, 22, 2592–2598. [Google Scholar] [CrossRef] [PubMed]
- Smolders, E.; Buekers, J.; Oliver, I.; McLaughlin, M.J. Soil properties affecting toxicity of zinc to soil microbial properties in laboratory-spiked and field-contaminated soils. Environ. Toxicol. Chem. 2004, 23, 2633–2640. [Google Scholar] [CrossRef] [PubMed]
- Sujkowska-Rybkowska, M.; Kasowska, D.; Gediga, K.; Banasiewicz, J.; Stępkowski, T. Lotus corniculatus-rhizobia symbiosis under Ni, Co and Cr stress on ultramafic soil. Plant Soil 2020, 451, 459–484. [Google Scholar] [CrossRef]
- Martíınez-Hidalgo, P.; Hirsch, A.M. The nodule microbiome: N2–fixing rhizobia do not live alone. Phytobiomes 2017, 1, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Rodrígez, H.; Fraga, R. Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 1999, 17, 319–339. [Google Scholar] [CrossRef]
- Ma, Y.; Prasad, M.N.; Rajkumar, M.; Freitas, H. Plant growth promoting rhizobacteria and endophytes accelerate phytoremediation of metalliferous soils. Biotechnol. Adv. 2011, 29, 248–258. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metals polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.H.; Koumoutsi, A.; Scholz, R.; Eisenreich, A.; Schneider, K.; Heinemeyer, I.; Morgenstern, B.; Voss, B.; Hess, W.R.; Reva, O.; et al. Comparative analysis of the complete genome sequence of the plant growth promoting bacterium Bacillus amyloliquefaciens FZB42. Nat. Biotechnol. 2007, 25, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Giller, K.; McGrath, S.; Hirsch, P. Absence of nitrogen fixation in clover grown on soil subject to long-term contamination with heavy metals is due to survival of only ineffective Rhizobium. Soil Biol. Biochem. 1989, 21, 841–848. [Google Scholar] [CrossRef]
- Brown, S.; Svendsen, A.; Henry, C. Restoration of high zinc and lead tailings with municipal biosolids and lime: A field study. J. Environ. Qual. 2009, 38, 2189–2197. [Google Scholar] [CrossRef]
- O’Day, P.A.; Vlassopoulos, D. Mineral-based amendments for remediation. Elements 2010, 6, 375–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adams, M.A.; Simon, J.; Pfautsch, S. Woody legumes: A (re)view from the south. Tree Physiol. 2010, 30, 1072–1082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajnaga, E.; Jach, M.E. Bradyrhizobia associated with Laburnum anagyroides, an exotic legume grown in Poland. Symbiosis 2020, 80, 245–255. [Google Scholar] [CrossRef] [Green Version]
- Gyaneshwar, P.; Hirsch, A.M.; Moulin, L.; Chen, W.-M.; Elliott, G.N.; Bontemps, C.; Estrada-de los Santos, P.; Gross, E.; Dos Reis, F.B.; Sprent, J.I.; et al. Legume-nodulating betaproteobacteria: Diversity, host range, and future prospects. Mol. Plant Microbe. Interact. 2011, 24, 1276–1288. [Google Scholar] [CrossRef] [Green Version]
- Bontemps, C.; Elliott, G.N.; Simon, M.F.; Dos Reis júnior, F.B.; Gross, E.; Lawton, R.C.; Neto, N.E.; de Fátima Loureiro, M.; De Faria, S.M.; Sprent, J.I.; et al. Burkholderia species are ancient symbionts of legumes. Mol. Ecol. 2010, 19, 44–52. [Google Scholar] [CrossRef]
- De Meyer, S.E.; Briscoe, L.; Martínez-Hidalgo, P.; Agapakis, C.M.; Estrada de-los Santos, P.; Seshadri, R.; Reeve, W.; Weinstock, G.; O’Hara, G.; Howieson, J.G.; et al. Symbiotic Burkholderia species show diverse arrangements of nif/fix and nod genes, and lack typical high affinity cytochrome cbb3 oxidase genes. Mol. Plant Microbe. Interact. 2016, 29, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Moulin, L.; Munive, A.; Dreyfus, B.; Boivin-Masson, C. Nodulation of legumes by members of the b-subclass of Proteobacteria. Lett. Nat. 2001, 411, 948–950. [Google Scholar] [CrossRef]
- Kalita, M.; Małek, W.; Coutinho, T.A. Putative novel Bradyrhizobium and Phyllobacterium species isolated from root nodules of Chamaecytisus ruthenicus. Syst. Appl. Microbiol. 2020, 43, 126056. [Google Scholar] [CrossRef]
- De Meyer, S.E.; De Beuf, K.; Vekeman, B. A large diversity of nonrhizobial endophytes found in legume root nodules in Flanders (Belgium). Soil Biol. Biochem. 2015, 83, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Radutoiu, S.; Madsen, L.H.; Madsen, E.B.; Felle, H.H.; Umehara, Y.; Gronlund, M.; Sato, S.; Nakamura, Y.; Tabata, S.; Sandal, N.; et al. Plant recognition of symbiotic bacteria requires two LysM receptor-like kinases. Nature 2003, 425, 585–592. [Google Scholar] [CrossRef]
- Rodríguez-Caballero, G.; Caravaca, F.; Fernández-González, A.J.; Alguacil, M.M.; Fernández-López, M.; Roldán, A. Arbuscular mycorrhizal fungi inoculation mediated changes in rhizosphere bacterial community structure while promoting revegetation in a semiarid ecosystem. Sci. Total. Environ. 2017, 584, 838–848. [Google Scholar] [CrossRef]
- Nzoué, A.; Domergue, O.; Moulin, L.; Avarre, J.-C.; de Lajudie, P. Tropical Legume Nodulating Bacteria. In Molecular Biology of Tropical Plants; Franche, C., Ed.; Research Signpost: Kerala, India, 2006; Volume 1, pp. 1–37. [Google Scholar]
- de Lajudie, P.; Willems, A.; Pot, B.; Dewettinck, D.; Maestrojuan, G.; Neyra, M.; Collins, M.D.; Dreyfus, B.; Kersters, K.; Gillis, M. Polyphasic taxonomy of rhizobia: Emendation of the genus Sinorhizobium and description of Sinorhizobium meliloti comb. nov., Sinorhizobium saheli sp. nov., and Sinorhizobium teranga sp. nov. Int. J. Syst. Bacteriol. 1994, 44, 715–733. [Google Scholar] [CrossRef]
- Nour, S.M.; Fernandez, M.P.; Normand, P.; Cleyet-Marel, J.C. Rhizobium ciceri sp. nov., consisting of strains that nodulate chickpea (Cicer arietinum L.). Int. J. Syst. Bacteriol. 1994, 44, 511–522. [Google Scholar] [CrossRef] [Green Version]
- Xu, L.M.; Ge, C.; Cui, Z.; Li, J.; Fan, H. Bradyrhizobium liaoningense sp. nov., isolated from the root nodules of soybeans. Int. J. Syst. Bacteriol. 1995, 45, 706–711. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.L.; Sun, J.G.; Li, Y.; Wang, E.T.; Chen, W.X. Numerical taxonomy and DNA relatedness of tropical rhizobia isolated from Hainan Province, China. Int. J. Syst. Bacteriol. 1994, 44, 151–158. [Google Scholar] [CrossRef]
- Chen, W.X.; Wang, E.T.; Wang, S.Y.; Ly, Y.B.; Chen, X.Q.; Li, Y. Characteristics of Rhizobium tianshanense sp. nov., a moderately and slowly growing root nodule bacterium. Int. J. Syst. Bacteriol. 1995, 45, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Parker, M.A. Relationships of bradyrhizobia from the legumes Apios americana and Desmodium glutinosum. Appl. Environ. Microbiol. 1999, 65, 4914–4920. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, J.T.; Eardly, B.D.; van Berkum, P.; Ronson, C.W. Four unnamed species of nonsymbiotic rhizobia isolated from the rhizosphere of Lotus corniculatus. Appl. Environ. Microbiol. 1996, 62, 2818–2825. [Google Scholar] [CrossRef] [Green Version]
- Vanlnsberghe, D.; Maas, K.R.; Cardenas, E.; Strachan, C.R.; Hallam, S.J.; Mohn, W.W. Non-symbiotic Bradyrhizobium ecotypes dominate north American forest soils. ISME J. 2015, 9, 2435–2441. [Google Scholar] [CrossRef] [Green Version]
- Downie, J.A.; Young, J.P. Genome sequencing. The ABC of symbiosis. Nature 2001, 412, 597–598. [Google Scholar] [CrossRef]
- Hawkins, J.P.; Oresnik, I.J. The rhizobium-legume symbiosis: Co-opting successful stress management. Front. Plant Sci. 2022, 12, 796045. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Downie, J.A. Coordinating nodule morphogenesis with rhizobial infection in legumes. Annu. Rev. Plant Biol. 2008, 59, 519–546. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Murray, J.D.; Poole, P.S.; Downie, J.A. The rules of engagement in the legume-rhizobial symbiosis. Annu. Rev. Genet. 2011, 45, 119–144. [Google Scholar] [CrossRef]
- Barnett, M.J.; Fisher, R.F. Global gene expression in the rhizobial-legume symbiosis. Symbiosis 2006, 42, 1–24. [Google Scholar]
- Geddes, B.A.; Oresnik, I.J. The mechanism of symbiotic nitrogen fixation. In The Mechanistic Benefits of Microbial Symbionts. Advances in Environmental Microbiology; Hurst, C.J., Ed.; International Publishing Springer: Cham, Switzerland, 2016; Volume 2, pp. 69–97. [Google Scholar]
- Madsen, E.B.; Madsen, L.H.; Radutoiu, S.; Olbryt, M.; Rakwalska, M.; Szczyglowski, K.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; et al. A receptor kinase gene of the LysM type is involved in legume perception of rhizobial signals. Nature 2003, 425, 637–640. [Google Scholar] [CrossRef]
- Hungria, M.; Menna, P.; Delamuta, J.R. Bradyrhizobium, the ancestor of all rhizobia: Phylogeny of housekeeping and nitrogen-fixation genes. In Biological Nitrogen Fixation; de Bruijn, F.J., Ed.; Wiley: Hoboken, NJ, USA, 2015; Volume 1, pp. 191–202. [Google Scholar]
- Parker, M.A. The spread of Bradyrhizobium lineages across host legume clades: From Abarema to Zygia. Microb. Ecol. 2015, 69, 630–640. [Google Scholar] [CrossRef]
- Jones, K.M.; Kobayashi, H.; Davies, B.W.; Taga, M.E.; Walker, G.C. How rhizobial symbionts invade plants: The Sinorhizobium-Medicago model. Nat. Rev. Microbiol. 2007, 5, 619–633. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, D.W.; Wais, R.; Long, S.R. Calcium spiking in plant root hairs responding to rhizobium nodulation signals. Cell 1996, 85, 673–681. [Google Scholar] [CrossRef] [Green Version]
- Shaw, S.L.; Long, S.R. Nod factor inhibition of reactive oxygen efflux in a host legume. Plant Physiol. 2003, 132, 2196–2204. [Google Scholar] [CrossRef] [Green Version]
- Fournier, J.; Timmers, A.C.J.; Sieberer, B.J.; Jauneau, A.; Chabaud, M.; Barker, D.G. Mechanism of infection thread elongation in root hairs of Medicago truncatula and dynamic interplay with associated rhizobial colonization. Plant Physiol. 2008, 148, 1985–1995. [Google Scholar] [CrossRef] [Green Version]
- Del Cerro, P.; Megías, M.; López-Baena, F.J.; Gil-Serrano, A.; Pérez-Montaño, F.; Ollero, F.J. Osmotic stress activates nif and fix genes and induces the rhizobium tropici CIAT 899 nod factor production via NodD2 by up-regulation of the nodA2 operon and the nodA3 gene. PLoS ONE 2019, 14, e0213298. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Montaño, F.; del Cerro, P.; Jiménez-Guerrero, I.; López-Baena, F.J.; Cubo, M.T.; Hungria, M.; Megías, M.; Ollero, F.J. RNA-seq analysis of the rhizobium tropici CIAT 899 transcriptome shows similarities in the activation patterns of symbiotic genes in the presence of apigenin and salt. BMC Genom. 2016, 17, 198. [Google Scholar] [CrossRef]
- Miller-Williams, M.; Loewen, P.C.; Oresnik, I.J. Isolation of salt-sensitive mutants of Sinorhizobium meliloti strain Rm1021. Microbiology 2006, 152, 2049–2059. [Google Scholar] [CrossRef] [Green Version]
- Dylan, T.; Nagpal, P.; Helinski, D.R.; Ditta, G.S. Symbiotic pseudorevertants of Rhizobium meliloti ndv mutants. J. Bacteriol. 1990, 172, 1409–1417. [Google Scholar] [CrossRef] [Green Version]
- Nagpal, P.; Khanuja, S.P.; Stanfield, S.W. Suppression of the ndv mutant phenotype of Rhizobium meliloti by cloned exo genes. Mol. Microbiol. 1992, 6, 479–488. [Google Scholar] [CrossRef]
- Miller, K.J.; Wood, J.M. Osmoadatptation by rhizosphere bacteria. Annu. Rev. Microbiol. 1996, 6, e23307. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.T.; Allaith, A.A.; Smith, G.M. Mechanism of osmotically regulated N-acetylglutaminylglutamine amide production in rhizobium meliloti. Plant Soil 1994, 161, 103–108. [Google Scholar] [CrossRef]
- Gober, J.W.; Kashket, E.R. K+ regulates bacteroid-associated functions of Bradyrhizobium. Proc. Natl. Acad. Sci. USA 1987, 84, 4650–4654. [Google Scholar] [CrossRef] [Green Version]
- Hunt, S.; King, B.J.; Canvin, D.T.; Layzell, D.B. Steady and nonsteady state gas exchange characteristics of soybean nodules in relation to the oxygen diffusion barrier. Plant Physiol. 1987, 84, 164–172. [Google Scholar] [CrossRef] [Green Version]
- Dixon, R.; Kahn, D. Genetic regulation of biological nitrogen fixation. Nat. Rev. Microbiol. 2004, 2, 621–631. [Google Scholar] [CrossRef] [PubMed]
- Zamorano-Sánchez, D.; Girard, L. FNR-like proteins in rhizobia: Past and future. In Biology Nitrogen Fixation; Brujin, F.J., Ed.; John Wiley & Sons: Seattle, WA, USA, 2015; Volume 1, pp. 155–166. [Google Scholar]
- Reyes-González, A.; Talbi, C.; Rodríguez, S.; Rivera, P.; Zamorano-Sánchez, D.; Girard, L. Expanding the regulatory network that controls nitrogen fixation in Sinorhizobium meliloti: Elucidating the role of the two-component system hFixL-FxkR. Microbiology 2016, 162, 979–988. [Google Scholar] [CrossRef] [PubMed]
- Rutten, P.J.; Steel, H.; Hood, G.A.; Ramachandran, V.K.; McMurtry, L.; Geddes, B.; Papachristodoulou, A.; Philip, S.; Poole, P.S. Multiple sensors provide spatiotemporal oxygen regulation of gene expression in a rhizobium-legume symbiosis. PLoS Genet. 2021, 17, e1009099. [Google Scholar] [CrossRef] [PubMed]
- Boscari, A.; Meilhoc, E.; Castella, C.; Bruand, C.; Puppo, A.; Brouquisse, R. Which role for nitric oxide in symbiotic N2-fixing nodules: Toxic by-product or useful signaling/metabolic intermediate? Front. Plant Sci. 2013, 4, 384. [Google Scholar] [CrossRef] [Green Version]
- Puppo, A.; Pauly, N.; Boscari, A.; Mandon, K.; Brouquisse, R. Hydrogen peroxide and nitric oxide: Key regulators of the legume-rhizobium and mycorrhizal symbioses. Antioxid. Redox Signal. 2013, 18, 2202–2219. [Google Scholar] [CrossRef]
- Lohar, D.P.; Haridas, S.; Gantt, J.S.; VandenBosch, K.A. A transient decrease in reactive oxygen species in roots leads to root hair deformation in the legume-rhizobia symbiosis. New Phytol. 2007, 173, 39–49. [Google Scholar] [CrossRef]
- Jamet, A.; Sigaud, S.; Van de Sype, G.; Puppo, A.; Herouart, D. Expression of the bacterial catalase genes during Sinorhizobium meliloti- Medicago sativa symbiosis and their crucial role during the infection process. Mol. Plant-Microbe Interact. 2003, 16, 217–225. [Google Scholar] [CrossRef] [Green Version]
- Pauly, N.; Pucciariello, C.; Mandon, K.; Innocenti, G.; Jamet, A.; Baudouin, E.; Hérouart, D.; Frendo, P.; Puppo, A. Reactive oxygen and nitrogen species and glutathione: Key players in the legume-rhizobium symbiosis. J. Exp. Bot. 2006, 57, 1769–1776. [Google Scholar] [CrossRef] [Green Version]
- Donati, A.J.; Jeon, J.M.; Sangurdekar, D.; So, J.S.; Chang, W.S. Genome-wide transcriptional and physiological responses of Bradyrhizobium japonicum to paraquat-mediated oxidative stress. Appl. Environ. Microbiol. 2011, 77, 3633–3643. [Google Scholar] [CrossRef] [Green Version]
- Fonseca-García, C.; Nava, N.; Lara, M.; Quinto, C. An NADPH oxidase regulates carbon metabolism and the cell cycle during root nodule symbiosis in common bean (Phaseolus vulgaris). BMC Plant Biol. 2021, 21, 274–290. [Google Scholar] [CrossRef]
- Lehman, A.P.; Long, S.R. Exopolysaccharides from Sinorhizobium meliloti can protect against H2O2-dependent damage. J. Bacteriol. 2013, 195, 5362–5369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faget, M.; Blossfeld, S.; von Gillhaussen, P.; Schurr, U.; Temperton, V.M. Disentangling who is who during rhizosphere acidification in root interactions: Combining fluorescence with optode techniques. Front. Plant Sci. 2013, 4, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pierre, O.; Engler, G.; Hopkins, J.; Brau, F.; Boncompagni, E.; Hérouart, D. Peribacteroid space acidification: A marker of mature bacteroid functioning in Medicago truncatula nodules. Plant Cell Environ. 2013, 36, 2059–2070. [Google Scholar] [PubMed]
- Geddes, B.A.; González, J.E.; Oresnik, I.J. Exopolysaccharide production in response to medium acidification is correlated with an increase in competition for nodule occupancy. Mol. Plant-Microbe Interact. 2014, 27, 1307–1317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrero-Castro, J.; Lozano, L.; Sohlenkamp, C. Dissecting the acid stress response of rhizobium tropici CIAT 899. Front. Microbiol. 2018, 9, 846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Richardson, A.E.; Rolfe, B.G. Studies of the physiological and genetic basis of acid tolerance in rhizobium leguminosarum biovar trifolii. Appl. Environ. Microbiol. 1993, 59, 1798–1804. [Google Scholar] [CrossRef] [Green Version]
- Fenner, B.J.; Tiwari, R.P.; Reeve, W.G.; Dilworth, M.J.; Glenn, A.R. Sinorhizobium medicae genes whose regulation involves the ActS and/or ActR signal transduction proteins. FEMS Microbiol. Lett. 2004, 236, 21–31. [Google Scholar] [CrossRef]
- Draghi, W.O.; Del Papa, M.F.; Hellweg, C.; Watt, S.A.; Watt, T.F.; Barsch, A.; Lozano, M.J.; Lagares, A., Jr.; Salas, M.E.; López, J.L.; et al. A consolidated analysis of the physiologic and molecular responses induced under acid stress in the legume-symbiont model-soil bacterium Sinorhizobium meliloti. Sci. Rep. 2016, 6, 29278. [Google Scholar] [CrossRef] [Green Version]
- Riccillo, P.M.; Muglia, C.I.; De Bruijn, F.J.; Roe, A.J.; Booth, I.R.; Aguilar, O.M. Glutathione is involved in environmental stress responses in rhizobium tropici, including acid tolerance. J. Bacteriol. 2000, 182, 1748–1753. [Google Scholar] [CrossRef] [Green Version]
- Muglia, C.I.; Grasso, D.H.; Aguilar, O.M. Rhizobium tropici response to acidity involves activation of glutathione synthesis. Microbiology 2007, 153, 1286–1296. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, S.D.; Munns, D.N. The correlation between extracellular polysaccharide production and acid tolerance in rhizobium. Soil Sci. Soc. Am. J. 1984, 48, 1273–1276. [Google Scholar] [CrossRef] [Green Version]
- Hawkins, J.P.; Geddes, B.A.; Oresnik, I.J. Succinoglycan directly contributes to pH tolerance in Sinorhizboium meliloti Rm1021. Mol. Plant-Microbe Interact. 2017, 30, 1009–1019. [Google Scholar] [CrossRef] [Green Version]
- Heavner, M.E.; Qiu, W.G.; Cheng, H. Phylogenetic co-occurrence of ExoR, ExoS, and ChvI, components of the RSI bacterial invasion switch, suggests a key adaptive mechanism regulating the transition between free-living and host-invading phases in Rhizobiales. PLoS ONE 2015, 10, e0135655. [Google Scholar] [CrossRef] [PubMed]
- Maillet, F.; Fournier, J.; Mendis, H.C.; Tadege, M.; Wen, J.; Ratet, P.; Mysore, K.S.; Gough, C.; Jones, K.M. Sinorhizobium meliloti succinylated high-molecular-weight succinoglycan and the Medicago truncatula LysM receptor-like kinase MtLYK10 participate independently in symbiotic infection. Plant J. 2020, 102, 311–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendis, H.C.; Madzima, T.F.; Queiroux, C.; Jones, K.M. Function of succinoglycan polysaccharide in Sinorhizobium meliloti host plant invasion depends on succinylation, not molecular weight. MBio 2016, 7, e00606-16. [Google Scholar] [CrossRef] [Green Version]
- Alunni, B.; Gourion, B. Terminal bacteroid differentiation in the legume-rhizobium symbiosis: Nodule-specific cysteine-rich peptides and beyond. New Phytol. 2016, 211, 411–417. [Google Scholar] [CrossRef] [Green Version]
- Haag, A.F.; Baloban, M.; Sani, M.; Kerscher, B.; Pierre, O.; Farkas, A.; Longhi, R.; Boncompagni, E.; Hérouart, D.; Dall’angelo, S.; et al. Protection of Sinorhizobium against host cysteine-rich antimicrobial peptides is critical for symbiosis. PLoS Biol. 2011, 9, e1001169. [Google Scholar] [CrossRef]
- Maroti, G.G.; Kereszt, A.; Kondorosi, E.; Mergaert, P. Natural roles of antimicrobial peptides in microbes, plants and animals. Res. Microbiol. 2011, 162, 363–374. [Google Scholar] [CrossRef]
- Guefrachi, I.; Pierre, O.; Timchenko, T.; Alunni, B.; Barrière, Q.; Czernic, P.; Villaécija-Aguilar, J.A.; Verly, C.; Bourge, M.; Fardoux, J.; et al. Bradyrhizobium BclA is a peptide transporter required for bacterial differentiation in Symbiosis with Aeschynomene legumes. Mol. Plant-Microbe Interact. 2015, 28, 1155–1166. [Google Scholar] [CrossRef] [Green Version]
- Khalid, A.; Tahir, S.; Arshad, M.; Zahir, Z.A. Relative efficiency of rhizobacteria for auxin biosynthesis in rhizosphere and non-rhizosphere soils. Soil Res. 2005, 42, 921–926. [Google Scholar] [CrossRef]
- Hao, X.; Xie, P.; Johnstone, L.; Miller, S.J.; Rensing, C.; Wei, G. Genome sequence and mutational analysis of plant growth promoting bacterium Agrobacterium tumefaciens CCNWGS0286 isolated from a zinc-lead mine tailing. Appl. Environ. Microbiol. 2012, 78, 5384–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianco, C.; Defez, R. Improvement of phosphate solubilization and Medicago plant yield by an indole-3-acetic acid-overproducing strain of Sinorhizobium meliloti. Appl. Environ. Microbiol. 2010, 76, 4626–4632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theunis, M.; Kobayashi, H.; Broughton, W.J.; Prinsen, E. Flavonoids, NodD1, NodD2, and nodbox NB15 modulate expression of the y4wEFG locus that is required for indole-3-acetic acid synthesis in Rhizobium sp. strain NGR234. Mol. Plant-Microbe Interact. 2004, 17, 1153–1161. [Google Scholar] [CrossRef] [Green Version]
- Barazani, O.; Friedman, J. Is IAA the major root growth factor secreted from plant-growth mediating bacteria? J. Chem. Ecol. 1999, 25, 2397–2406. [Google Scholar] [CrossRef]
- Dazzo, F.; Biswas, J.; Ladha, J. Rhizobia inoculation improves nutrient uptake and growth of lowland rice. Soil Sci. Soc. Am. J. 2000, 64, 1644–1650. [Google Scholar]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [Green Version]
- Glick, B.R. Modulation of plant ethylene levels by the bacterial enzyme ACC deaminase. FEMS Microbiol. Lett. 2005, 251, 1–7. [Google Scholar] [CrossRef]
- Ghosh, S.; Basu, P.S. Production and metabolism of indole acetic acid in roots and root nodules of Phaseolus mungo. Microbiol. Res. 2006, 161, 362–366. [Google Scholar] [CrossRef]
- Penmetsa, R.V.; Frugoli, J.A.; Smith, L.S.; Long, S.R.; Cook, D.R. Dual genetic pathways controlling nodule number in Medicago truncatula. Plant Physiol. 2003, 131, 998–1008. [Google Scholar] [CrossRef] [Green Version]
- Camerini, S.; Senatore, B.; Lonardo, E.; Imperlini, E.; Bianco, C.; Moschetti, G.; Rotino, G.L.; Campion, B.; Defez, R. Introduction of a novel pathway for IAA biosynthesis to rhizobia alters vetch root nodule development. Arch. Microbiol. 2008, 190, 67–77. [Google Scholar] [CrossRef]
- Prinsen, E.; Chauvaux, N.; Schmidt, J.; John, M.; Wieneke, U.; De Greef, J.; Schell, J.; Van Onckelen, H. Stimulation of indole-3-acetic acid production in Rhizobium by flavonoids. FEBS Lett. 1991, 282, 53–55. [Google Scholar] [CrossRef] [Green Version]
- Purchase, D.; Miles, R.; Young, T. Cadmium uptake and nitrogen fixing ability in heavy-metal resistant laboratory and field strains of Rhizobium leguminosarum biovar trifolii. FEMS Microb. Ecol. 1997, 22, 85–93. [Google Scholar] [CrossRef]
- Singh, S.; Kayastha, A.M.; Asthana, R.K.; Srivastava, P.K.; Singh, S. Response of Rhizobium leguminosarum to nickel stress. World J. Microb. Biot. 2001, 17, 667–672. [Google Scholar] [CrossRef]
- Lima, A.I.G.; Corticeiro, S.C.; de Almeida Paula Figueira, E.M. Glutathione-mediated cadmium sequestration in Rhizobium leguminosarum. Enzym. Microb. Technol. 2006, 39, 763–769. [Google Scholar] [CrossRef]
- Lakzian, A.; Murphy, P.; Giller, K.E. Transfer and loss of naturally-occurring plasmids among isolates of Rhizobium leguminosarum bv. viciae in heavy metal contaminated soils. Soil Biol. Biochem. 2007, 39, 1066–1077. [Google Scholar] [CrossRef]
- Luo, S.L.; Chen, L.; Chen, J.L.; Xiao, X.; Xu, T.Y.; Wan, Y.; Rao, C.; Liu, C.B.; Liu, Y.T.; Lai, C.; et al. Analysis and characterization of cultivable heavy metal-resistant bacterial endophytes isolated from Cd-hyperaccumulator Solanum nigrum L. and their potential use for phytoremediation. Chemosphere 2011, 85, 1130–1138. [Google Scholar] [CrossRef]
- Martinez, R.J.; Wang, Y.; Raimondo, M.A.; Coombs, J.M.; Barkay, T.; Sobecky, P.A. Horizontal gene transfer of PIB-Type ATPases among bacteria isolated from radionuclide-and metal-contaminated subsurface soils. Appl. Environ. Microbiol. 2006, 72, 3111–3118. [Google Scholar] [CrossRef] [Green Version]
- Abdelkrim, S.; Jebara, S.H.; Jebara, M. Antioxidant systems responses and the compatible solutes as contributing factors to lead accumulation and tolerance in Lathyrus sativus inoculated by plant growth promoting rhizobacteria. Ecotoxicol. Environ. Saf. 2018, 166, 427–436. [Google Scholar] [CrossRef]
- Chiboub, M.; Jebara, S.H.; Saadani, O.; Fatnassi, I.C.; Abdelkrim, S.; Abid, G.; Jebara, M.; Harzalli Jebara, S. Characterization of efficient plant-growth-promoting bacteria isolated form Sulla coronaria resistant to cadmium and to other heavy metals. Comptes Rendus Biol. 2016, 339, 391–398. [Google Scholar] [CrossRef]
- Del Rio, M.; Font, R.; Almela, C.; Velez, D.; Montoro, R.; De Haro Bailon, A. Heavy metals and arsenic uptake by wild vegetation in the Guadiamar river area after the toxic spill of the Aznalcollar mine. J. Biotechnol. 2002, 98, 125–137. [Google Scholar] [CrossRef]
- Bai, T.; Liu, Y.-Y.; Muhammad, I.; Yang, X.; Yin, X.-J.; Bai, L.; Wang, Y.-J. Mixed nitrogen form addition facilitates the growth adaptation of legume plant to heavy metal contamination in degraded mining areas. Glob. Ecol. Conserv. 2020, 24, e01387. [Google Scholar] [CrossRef]
- Sujkowska-Rybkowska, M.; Muszyńska, E.; Labudda, M. Structural adaptation and physiological mechanisms in the leaves of Anthyllis vulneraria L. from metallicolous and non-metallicolous populations. Plants 2020, 9, 662. [Google Scholar] [CrossRef]
- Chaney, R.L.; Siebielec, G.; Li, Y.M.; Kerschner, B.A. Response of four turfgrass cultivars to limestone and biosolids-compost amendment of a zinc and cadmium contaminated soil at Palmerton, Pennsylvania. J. Environ. Qual. 2000, 29, 1440–1447. [Google Scholar]
- Chaney, R.L.; Broadhurst, C.L.; Centofanti, T. Phytoremediation of Soil Trace Elements. In Trace Elements in Soils; Hooda, P., Ed.; Blackwell Publishers: Oxford, UK, 2010; Volume 1, pp. 311–352. [Google Scholar]
- Carrasco, J.A.; Armario, P.; Pajuelo, E.; Burgos, A.; Caviedes, M.A.; López, R.; Chamber, M.A.; Palomares, J.A. Isolation and characterisation of symbiotically effective Rhizobium resistant to arsenic and heavy metals after the toxic spill at the Aznalcóllar pyrite mine. Soil Biol. Biochem. 2005, 37, 1131–1140. [Google Scholar] [CrossRef]
- Vara Prasad, M.N.; de Oliveira Freitas, H.M. Metal hyperaccumulation in plants: Biodiversity prospecting for phytoremediation technology. Electron. J. Biotechn. 2003, 6, 285–321. [Google Scholar] [CrossRef]
- Bolan, N.; Kunhikrishnan, A.; Thangarajan, R.; Kumpiene, J.; Park, J.; Makino, T.; Kirkham, M.B.; Scheckel, K. Remediation of heavy metal(loid)s contaminated soils-to mobilize or to immobilize? J. Hazard. Mater. 2014, 266, 141–166. [Google Scholar] [CrossRef]
- Pramanik, K.; Mitra, S.; Sarkar, A.; Maiti, T.K. Alleviation of phytotoxic effects of cadmium on rice seedlings by cadmium resistant PGPR strain Enterobacter aerogenes MCC 3092. J. Hazard. Mater. 2018, 351, 317–329. [Google Scholar] [CrossRef]
- de Sousa Leite, T.S.; Monteiro, F.A. Nitrogen form regulates cadmium uptake and accumulation in Tanzania guinea grass used for phytoextraction. Chemosphere 2019, 236, 124324. [Google Scholar] [CrossRef]
- Leite, T.S.; Monteiro, F.A. Partial replacement of nitrate by ammonium increases photosynthesis and reduces oxidative stress in Tanzania guinea grass exposed to cadmium. Ecotoxicol. Environ. Saf. 2019, 174, 592–600. [Google Scholar] [CrossRef]
- Checcucci, A.; Bazzicalupo, M.; Mengoni, A. Exploiting nitrogen-fixing rhizobial symbionts genetic resources for improving phytoremediation of contaminated soils. In Enhancing Cleanup of Environmental Pollutants; Springer: Heidelberg, Germany, 2017; Volume 1, pp. 275–288. [Google Scholar]
- Hao, X.L.; Xie, P.; Zhu, Y.G.; Taghavi, S.; Wei, G.H.; Rensing, C. Copper tolerance mechanisms of Mesorhizobium amorphae and its role in aiding phytostabilization by Robinia pseudoacacia in copper contaminated soil. Environ. Sci. Technol. 2015, 49, 2328–2340. [Google Scholar] [CrossRef]
- Sánchez-Navarro, V.; Zornoza, R.; Faz, A.; Fernandez, J.A. Comparing legumes for use in multiple cropping to enhance soil organic carbon, soil fertility, aggregates stability and vegetables yields under semiarid conditions. Sci. Hortic. 2019, 246, 835–841. [Google Scholar] [CrossRef]
- Zhang, X.; Lou, X.; Zhang, H.; Ren, W.; Tang, M. Effects of sodium sulfide application on the growth of Robinia pseudoacacia, heavy metal immobilization, and soil microbial activity in PbeZn polluted soil. Ecotoxicol. Environ. Saf. 2020, 197, 110563. [Google Scholar] [CrossRef]
- de-Bashan, L.E.; Hernandez, J.P.; Bashan, Y. The potential contribution of plant growth-promoting bacteria to reduce environmental degradation–A comprehensive evaluation. Appl. Soil Ecol. 2011, 61, 171–189. [Google Scholar] [CrossRef]
- Teng, Y.; Wang, X.; Li, L.; Li, Z.; Luo, Y. Rhizobia and their bio-partners as novel drivers for functional remediation in contaminated soils. Front. Plant Sci. 2015, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Fritioff, Å.; Kautsky, L.; Greger, M. Influence of temperature and salinity on heavy metal uptake by submersed plants. Environ. Pollut. 2005, 133, 265–274. [Google Scholar] [CrossRef]
- Pastor, J.; Hernandez, A.J.; Prieto, N.; Fernandez-Pascual, M. Accumulating behaviour of Lupinus albus L. growing in a normal and a decalcified calcic luvisol polluted with Zn. J. Plant Physiol. 2003, 160, 1457–1465. [Google Scholar] [CrossRef] [Green Version]
- Vazquez, S.; Moreno, E.; Carpena, R.O. Bioavailability of metals and as from acidified multicontaminated soils: Use of white lupin to validate several extraction methods. Environ. Geochem. Health 2008, 30, 193–198. [Google Scholar] [CrossRef]
- Freeman, J.L.; Zhang, L.H.; Marcus, M.A.; Fakra, S.; McGrath, S.P.; Pilon-Smits, E.A. Spatial imaging, speciation, and quantification of selenium in the hyperaccumulator plants Astragalus bisulcatus and Stanleya pinnata. Plant Physiol. 2006, 142, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Xu, S.; Zhang, D.; Fu, Z.; Zhang, H.; Zhu, H. Phytoremediation of Secondary Salinity in Greenhouse Soil with Astragalus sinicus, Spinacea oleracea and Lolium perenne. Agriculture 2022, 12, 212. [Google Scholar] [CrossRef]
- Liu, Z.; Tran, K.Q. A review on disposal and utilization of phytoremediation plants containing heavy metals. Ecotoxicol. Environ. Saf. 2021, 226, 112821. [Google Scholar] [CrossRef]
- Ju, W.L.; Liu, L.; Fang, L.C.; Cui, Y.X.; Duan, C.J.; Wu, H. Impact of co-inoculation with plant-growth-promoting rhizobacteria and rhizobium on the biochemical responses of alfalfa-soil system in copper contaminated soil. Ecotoxicol. Environ. Saf. 2019, 167, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.J.; Ma, Z.Q.; Sun, L.L.; Han, M.S.; Lu, J.J.; Li, Z.X.; Mohamad, O.A.; Wei, G.H. Extracellular polymeric substances from copper-tolerance Sinorhizobium meliloti immobilize Cu2+. J. Hazard. Mater. 2013, 261, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Etesami, H. Bacterial mediated alleviation of heavy metal stress and decreased accumulation of metals in plant tissues: Mechanisms and future prospects. Ecotoxicol. Environ. Saf. 2018, 147, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Hua, Y.P.; Chen, M.X.; Zhang, J.H.; Guan, C.Y.; Zhang, Z.H. Mechanism enhancing Arabidopsis resistance to cadmium: The role of NRT1.5 and proton pump. Front. Plant Sci. 2018, 9, 1892. [Google Scholar] [CrossRef] [Green Version]
- Xie, P.; Hao, X.L.; Herzberg, M.; Luo, Y.T.; Nies, D.H.; Wei, G.H. Genomic analyses of metal resistance genes in three plant growth promoting bacteria of legume plants in Northwest mine tailings, China. J. Environ. Sci. 2015, 27, 179–187. [Google Scholar] [CrossRef]
- Rajkumar, M.; Ae, N.; Prasad, M.N.V.; Freitas, H. Potential of siderophore-producing bacteria for improving heavy metal phytoextraction. Trends Biotechnol. 2010, 28, 142–149. [Google Scholar] [CrossRef]
- Saadani, O.; Fatnassi, I.C.; Chiboub, M.; Abdelkrim, S.; Barhoumi, F.; Jebara, M.; Jebara, S.H. In situ phytostabilisation capacity of three legumes and their associated Plant Growth Promoting Bacteria (PGPBs) in mine tailings of northern Tunisia. Ecotoxicol. Environ. Saf. 2016, 130, 263–269. [Google Scholar] [CrossRef]
- Duan, C.J.; Fang, L.C.; Yang, C.L.; Chen, W.B.; Cui, Y.X.; Li, S.Q. Reveal the response of enzyme activities to heavy metals through in situ zymography. Ecotoxicol. Environ. Saf. 2018, 156, 106–115. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, E.Y.; Hyun, S.; Kim, J.G. Metal availability in heavy metal-contaminated open burning and open detonation soil: Assessment using soil enzymes, earthworms, and chemical extractions. J. Hazard. Mater. 2009, 170, 382–388. [Google Scholar] [CrossRef]
- Shenker, M.; Fan, T.W.M.; Crowley, D.E. Phytosiderophores influence on cadmium mobilization and uptake by wheat and barley plants. J. Environ. Qual. 2001, 30, 2091–2098. [Google Scholar] [CrossRef]
- Rahman, T.; Seraj, M.F. Available approaches of remediation and stabilization of metal contamination in soil: A review. Am. J. Plant Sci. 2018, 9, 2033–2052. [Google Scholar] [CrossRef] [Green Version]
- Fan, M.; Xiao, X.; Guo, Y.; Zhang, J.; Wang, E.; Chen, W.; Lin, Y.; Wei, G. Enhanced phytoremediation of Robinia pseudoacatia in heavy metal-contamined soils with rhizobia and the associated bacterial community structure and function. Chemosphere 2018, 197, 729–740. [Google Scholar] [CrossRef]
- Backer, R.; Rokem, J.S.; Ilangumaran, G.; Lamont, J.; Praslickova, D.; Ricci, E.; Subramanian, S.; Smith, D.L. Plant growth-promoting rhizobacteria: Context, mechanisms of action, and roadmap to commercialization of biostimulants for sustainable agriculture. Front. Plant Sci. 2018, 9, 1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abhilash, P.C.; Powell, J.R.; Singh, H.B.; Singh, B.K. Plant-microbe interactions: Novel applications for exploitation in multipurpose remediation technologies. Trends Biotechnol. 2012, 30, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Kotrba, P.; Najmanova, J.; Macek, T.; Ruml, T.; Mackova, M. Genetically modified plants in phytoremediation of heavy metal and metalloid soil and sediment pollution. Biotechnol. Adv. 2009, 27, 799–810. [Google Scholar] [CrossRef]
- Tussipkan, D.; Manabayeva, S.A. Alfalfa (Medicago Sativa L.): Genotypic Diversity and Transgenic Alfalfa for Phytoremediation. Front. Environ. Sci. 2022, 10, 87. [Google Scholar] [CrossRef]
- Hussein, H.S.; Ruiz, O.N.; Terry, N.; Daniell, H. Phytoremediation of mercury and organomercurials in chloroplast transgenic plants: Enhanced root uptake, translocation to shoots, and volatilization. Environ. Sci. Technol. 2007, 41, 8439–8446. [Google Scholar] [CrossRef] [Green Version]
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.-J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [Green Version]
- Koźmińska, A.; Wiszniewska, A.; Hanus-Fajerska, E.; Muszyńska, E. Recent strategies of increasing metal tolerance and phytoremediation potential using genetic transformation of plants. Plant Biotechnol. Rep. 2018, 12, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Venegas-Rioseco, J.; Ginocchio, R.; Ortiz-Calderón, C. Increase in Phytoextraction Potential by Genome Editing and Transformation: A Review. Plants 2021, 11, 86. [Google Scholar] [CrossRef]
- Shim, D.; Kim, S.; Choi, Y.I.; Song, W.Y.; Park, J.; Youk, E.S.; Jeong, S.C.; Martinoia, E.; Noh, E.W.; Lee, Y. Transgenic poplar trees expressing yeast cadmium factor 1 exhibit the characteristics necessary for the phytoremediation of mine tailing soil. Chemosphere 2013, 90, 1478–1486. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Han, J.; Xue, W.; Wang, J.; Wang, B.; Liu, L.; Zou, J. Overexpression of SmZIP plays important roles in Cd accumulation and translocation, subcellular distribution, and chemical forms in transgenic tobacco under Cd stress. Ecotoxicol. Environ. Saf. 2021, 214, 112097. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, M.S.U.; Min, S.R.; Jeong, W.J.; Sultana, S.; Choi, K.S.; Lee, Y.; Liu, J.R. Overexpression of AtATM3 in Brassica juncea confers enhanced heavy metal tolerance and accumulation. Plant Cell Tissue Organ Cult. 2011, 107, 69–77. [Google Scholar] [CrossRef]
- Wang, C.; Chen, X.; Yao, Q.; Long, D.; Fan, X.; Kang, H.; Zeng, J.; Sha, L.; Zhang, H.; Zhou, Y.; et al. Overexpression of TtNRAMP6 enhances the accumulation of Cd in Arabidopsis. Gene 2019, 696, 225–232. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, M.; Tian, S.; Lu, L.; Shohag, M.J.; Yang, X. Metallothionein 2 (SaMT2) from Sedum alfredii Hance confers increased Cd tolerance and accumulation in yeast and tobacco. PLoS ONE 2014, 9, e102750. [Google Scholar] [CrossRef]
- Zhao, C.; Xu, J.; Li, Q.; Li, S.; Wang, P.; Xiang, F. Cloning and characterization of a Phragmites australis phytochelatin synthase (PaPCS) and achieving Cd tolerance in tall fescue. PLoS ONE 2014, 9, e103771. [Google Scholar] [CrossRef]
- Zhu, S.; Shi, W.; Jie, Y. Overexpression of BnPCS1, a Novel Phytochelatin Synthase Gene from Ramie (Boehmeria nivea), Enhanced Cd Tolerance, Accumulation, and Translocation in Arabidopsis thaliana. Front. Plant Sci. 2021, 12, 639189. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.; Pareek, A.; Singla-Pareek, S.L. Plant Metallothioneins: Classification, Distribution, Function, and Regulation. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Oxford, UK, 2016; pp. 239–261. [Google Scholar]
- Grill, E.; Löffler, S.; Winnacker, E.L.; Zenk, M.H. Phytochelatins, the heavy-metal-binding peptides of plants, are synthesized from glutathione by a specific gamma-glutamylcysteine dipeptidyl transpeptidase (phytochelatin synthase). Proc. Natl. Acad. Sci. USA 1989, 86, 6838–6842. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Trivedi, P.K. Glutathione S-Transferases: Role in Combating Abiotic Stresses Including Arsenic Detoxification in Plants. Front. Plant Sci. 2018, 9, 751. [Google Scholar] [CrossRef] [Green Version]
- Kumar, V.; AlMomin, S.; Al-Shatti, A.; Al-Aqeel, H.; Al-Salameen, F.; Shajan, A.B.; Nair, S.M. Enhancement of heavy metal tolerance and accumulation efficiency by expressing Arabidopsis ATP sulfurylase gene in alfalfa. Int. J. Phytoremediation 2019, 1, 1112–1121. [Google Scholar] [CrossRef]
- Xu, W.; Shi, W.; Liu, F.; Ueda, A.; Takabe, T. Enhanced zinc and cadmium tolerance and accumulation in transgenic Arabidopsis plants constitutively overexpressing a barley gene (HvAPX1) that encodes a peroxisomal ascorbate peroxidase. Botany 2008, 86, 567–575. [Google Scholar] [CrossRef]
- Faè, M.; Balestrazzi, A.; Confalonieri, M.; Donà, M.; Macovei, A.; Valassi, A.; Giraffa, G.; Carbonera, D. Copper-mediated genotoxic stress is attenuated by the overexpression of the DNA repair gene MtTdp2α (tyrosyl-DNA phosphodiesterase 2) in Medicago truncatula plants. Plant Cell Rep. 2014, 33, 1071–1080. [Google Scholar] [CrossRef] [PubMed]
- Charfeddine, M.; Charfeddine, S.; Bouaziz, D.; Messaoud, R.B.; Bouzid, R.G. The effect of cadmium on transgenic potato (Solanum tuberosum) plants overexpressing the StDREB transcription factors. Plant Cell Tissue Organ Cult. 2017, 128, 521–541. [Google Scholar] [CrossRef]
- Ahammed, G.J.; Li, C.X.; Li, X.; Liu, A.; Chen, S.; Zhou, J. Overexpression of tomato RING E3 ubiquitin ligase gene SlRING1 confers cadmium tolerance by attenuating cadmium accumulation and oxidative stress. Physiol. Plant 2021, 173, 449–459. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Dai, X.; Xu, W.; Ma, M. Overexpressing GSH1 and AsPCS1 simultaneously increases the tolerance and accumulation of cadmium and arsenic in Arabidopsis thaliana. Chemosphere 2008, 72, 1020–1026. [Google Scholar] [CrossRef]
- LeDuc, D.L.; Tarun, A.S.; Montes-Bayon, M.; Meija, J.; Malit, M.F.; Wu, C.P.; AbdelSamie, M.; Chiang, C.Y.; Tagmount, A.; deSouza, M.; et al. Overexpression of selenocysteine methyltransferase in Arabidopsis and Indian mustard increases selenium tolerance and accumulation. Plant Physiol. 2004, 135, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.; Li, M.; Sinharoy, S.; Verdier, J. A Snapshot of Functional Genetic Studies in Medicago truncatula. Front. Plant Sci. 2016, 7, 1175. [Google Scholar] [CrossRef] [Green Version]
- Boisson-Dernier, A.; Chabaud, M.; Garcia, F.; Bécard, G.; Rosenberg, C.; Barker, D.G. Agrobacterium rhizogenes-transformed roots of Medicago truncatula for the study of nitrogen-fixing and endomycorrhizal symbiotic associations. Mol. Plant-Microbe Interact. 2001, 14, 695–700. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Fu, C.; Wang, Z.Y. A Unified Agrobacterium-Mediated Transformation Protocol for Alfalfa (Medicago sativa L.) and Medicago truncatula. Methods Mol. Biol. 2019, 1864, 153–163. [Google Scholar]
- Pérez-Palacios, P.; Romero-Aguilar, A.; Delgadillo, J.; Doukkali, B.; Caviedes, M.A.; Rodríguez-Llorente, I.D.; Pajuelo, E. Double genetically modified symbiotic system for improved Cu phytostabilization in legume roots. Environ. Sci. Pollut. Res. Int. 2017, 24, 14910–14923. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J. Transgenic alfalfa plants co-expressing glutathione S-transferase (GST) and human CYP2E1 show enhanced resistance to mixed contaminates of heavy metals and organic pollutants. J. Hazard. Mater. 2011, 189, 357–362. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, J.; Zhou, Y.; Gong, T.; Wang, J.; Ge, Y. Enhanced phytoremediation of mixed heavy metal (mercury)-organic pollutants (trichloroethylene) with transgenic alfalfa co-expressing glutathione S-transferase and human P450 2E1. J. Hazard. Mater. 2013, 260, 1100–1107. [Google Scholar] [CrossRef]
- Basu, S.; Rabara, R.C.; Negi, S.; Shukla, P. Engineering PGPMOs through Gene Editing and Systems Biology: A Solution for Phytoremediation? Trends Biotechnol. 2018, 36, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Sriprang, R.; Murooka, Y. Accumulation and Detoxification of Metals by Plants and Microbes. In Environmental Bioremediation Technologies; Singh, S.N., Tripathi, R.D., Eds.; Springer: Berlin/Heidelberg, Germany, 2007; pp. 77–99. [Google Scholar]
- Sriprang, R.; Hayashi, M.; Yamashita, M.; Ono, H.; Saeki, K.; Murooka, Y. A novel bioremediation system for heavy metals using the symbiosis between leguminous plant and genetically engineered rhizobia. J. Biotechnol. 2002, 99, 279–293. [Google Scholar] [CrossRef]
- Sriprang, R.; Hayashi, M.; Ono, H.; Takagi, M.; Hirata, K.; Murooka, Y. Enhanced accumulation of Cd2+ by a Mesorhizobium sp. transformed with a gene from Arabidopsis thaliana coding for phytochelatin synthase. Appl. Environ. Microbiol. 2003, 69, 1791–1796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ike, A.; Sriprang, R.; Ono, H.; Murooka, Y.; Yamashita, M. Bioremediation of cadmium contaminated soil using symbiosis between leguminous plant and recombinant rhizobia with the MTL4 and the PCS genes. Chemosphere 2007, 66, 1670–1676. [Google Scholar] [CrossRef] [PubMed]
- Tsyganov, V.E.; Tsyganova, A.V.; Gorshkov, A.P.; Seliverstova, E.V.; Kim, V.E.; Chizhevskaya, E.P.; Belimov, A.A.; Serova, T.A.; Ivanova, K.A.; Kulaeva, O.A.; et al. Efficacy of a Plant-Microbe System: Pisum sativum (L.) cadmium-tolerant mutant and Rhizobium leguminosarum strains, expressing pea metallothionein Genes PsMT1 and PsMT2, for cadmium phytoremediation. Front. Microbiol. 2020, 11, 15. [Google Scholar] [CrossRef]
- Delgadillo, J.; Lafuente, A.; Doukkali, B.; Redondo-Gómez, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Improving legume nodulation and Cu rhizostabilization using a genetically modified rhizobia. Environ. Technol. 2015, 36, 1237–1245. [Google Scholar] [CrossRef]
- Pajuelo, E.; Carrasco, J.A.; Romero, L.C.; Chamber, M.A.; Gotor, C. Evaluation of the metal phytoextraction potential of crop legumes. Regulation of the expression of O-acetylserine (thiol)lyase under metal stress. Plant Biol. 2007, 9, 672–681. [Google Scholar] [CrossRef]
- Zhang, J.; Xu, Y.; Cao, T.; Chen, J.; Rosen, B.P.; Zhao, F.J. Arsenic methylation by a genetically engineered Rhizobium-legume symbiont. Plant Soil 2017, 416, 259–269. [Google Scholar] [CrossRef]
- Espinoza, C.; Schlechter, R.; Herrera, D.; Torres, E.; Serrano, A.; Medina, C.; Arce-Johnson, P. Cisgenesis and intragenesis: New tools for improving crops. Biol. Res. 2013, 46, 323–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Checcucci, A.; diCenzo, G.C.; Ghini, V.; Bazzicalupo, M.; Beker, A.; Decorosi, F.; Döhlemann, J.; Fagorzi, C.; Finan, T.M.; Fondi, M.; et al. Creation and multi-omics charac-terization of a genomically hybrid strain in the nitrogen-fixing symbiotic bacterium Sinorhizobium meliloti. bioRxiv 2018, 296483. [Google Scholar] [CrossRef]
- Landeta, C.; Dávalos, A.; Cevallos, M.Á.; Geiger, O.; Brom, S.; Romero, D. Plasmids with a chromosome-like role in rhizobia. J. Bacteriol. 2011, 193, 1317–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- diCenzo, G.C.; Benedict, A.B.; Fondi, M.; Walker, G.C.; Finan, T.M.; Mengoni, A.; Griffitts, J.S. Robustness encoded across essential and accessory replicons of the ecologically versatile bacterium Sinorhizobium meliloti. PLoS Genet. 2018, 14, e1007357. [Google Scholar] [CrossRef] [Green Version]
- Drewniak, L.; Dziewit, L.; Ciężkowska, M.; Gawor, J.; Gromadka, R.; Skłodowska, A. Structural and functional genomics of plasmid pSinA of Sinorhizobium sp. M14 encoding genes for the arsenite oxidation and arsenic resistance. J. Biotechnol. 2013, 164, 479–488. [Google Scholar] [CrossRef] [Green Version]
- El-Sappah, A.H.; Yan, K.; Huang, Q.; Islam, M.M.; Li, Q.; Wang, Y.; Khan, M.S.; Zhao, X.; Mir, R.R.; Li, J.; et al. Comprehensive Mechanism of Gene Silencing and Its Role in Plant Growth and Development. Front. Plant Sci. 2021, 12, 705249. [Google Scholar] [CrossRef]
- Dhankher, O.P.; Rosen, B.P.; McKinney, E.C.; Meagher, R.B. Hyperaccumulation of arsenic in the shoots of Arabidopsis silenced for arsenate reductase (ACR2). Proc. Natl. Acad. Sci. USA 2006, 103, 5413–5418. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Bashir, K.; Senoura, T.; Sugimoto, K.; Ono, K.; Suzui, N.; Kawachi, N.; Ishii, S.; et al. From Laboratory to Field: OsNRAMP5-Knockdown Rice Is a Promising Candidate for Cd Phytoremediation in Paddy Fields. PLoS ONE 2014, 9, e98816. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Huang, S.Q.; Yang, Z.M. Bioinformatic identification and expression analysis of new microRNAs from Medicago truncatula. Biochem. Biophys. Res. Commun. 2008, 374, 538–542. [Google Scholar] [CrossRef]
- Bao, A.; Burritt, D.J.; Chen, H.; Zhou, X.; Cao, D.; Tran, L.P. The CRISPR/Cas9 system and its applications in crop genome editing. Crit. Rev. Biotechnol. 2019, 39, 321–336. [Google Scholar] [CrossRef]
- Tang, L.; Mao, B.; Li, Y.; Lv, Q.; Zhang, L.; Chen, C.; He, H.; Wang, W.; Zeng, X.; Shao, Y.; et al. Knockout of OsNramp5 using the CRISPR/Cas9 system produces low Cd-accumulating indica rice without compromising yield. Sci. Rep. 2017, 7, 14438. [Google Scholar] [CrossRef] [PubMed] [Green Version]

| Heavy Metals | The Toxic Effects of Heavy Metals on Plants | References |
|---|---|---|
| Cd | Reduction in biomass and root length; inhibition of seed germination; growth reduction; wilting; chlorosis, and cell damage | [56,57,78,79,80,81] |
| Cu | Inhibition of root, shoot, and leaf development; quantity reduction in leaves per plant; decreased antioxidant activities; shoot length reduction; decreased total chlorophyll content; reduction in chlorophyl biosynthesis; decreased enzyme activities; decreased plant growth and yield; leaf chlorosis; generation of oxidative stress and ROS | [56,57,77,79,82,83,84,85,86] |
| Zn | Decreased total chlorophyll content; reduction in transpiration rate, inhibition of transport of microelements; limitation of root and shoot growth; reduction in photosynthetic and respiratory rate; enhancement of generation of reactive oxygen species; chlorosis in the younger leaves; reduction in germination | [56,57,79,87,88,89,90] |
| As | Inhibition of growth and low crop production; reduction in leaf quantities; chlorosis; leaf senescence necrosis; defoliation; reduction in leaf area and dry matter production; reduction in shoot and root growth; restricted stomatal conductance and nutrient uptake; chlorophyll degradation; limited biomass and yield production; overproduction of reactive oxygen species (ROS) leading to carbohydrate, protein, and DNA damage. | [57,91,92,93,94] |
| Ni | Reduction in chlorophyll content; decreased levels of sugar, starch, and protein nitrogen; decrease in shoot yield; chlorosis; inhibition of root growth; inhibition of growth, induction of chlorosis, necrosis, and wilting; generation of ROS | [57,79,95,96,97] |
| Pb | DNA damage; decrease in chlorophyll content; decrease in protein content; stunted foliage; reduction in photosynthesis; impaired nutrient uptake; decrease in seed germination, root elongation, decreased biomass; inhibition of chlorophyll biosynthesis; inhibition of mineral nutrition and enzymatic reactions, induction of ROS production | [42,56,57,98,99,100] |
| Cr | Inhibition of root, steam, and leaf growth; inhibition of chlorophyll biosynthesis; induction of oxidative stress; inhibition of photosynthesis; inhibition of seed germination and seedling development; reduction in root and shoot biomass, quality of flowers, and crop yield | [57,77,101,102,103] |
| Co | Inhibition of plant growth; chlorosis in young leaves; reduction in biomass; inhibition of greening | [57,76,104,105] |
| Fe | Reduction in root and shoot growth; hindered growth, reduction in chlorophyll content in older leaves, decreased sugars, starch, and protein nitrogen contents | [106,107] |
| Mn | Reduction in biomass production; adversely affects nutrient uptake; hindered seedling growth; induction of oxidative stress | [108,109] |
| Symbiotic Species | Heavy Metal | References | |
|---|---|---|---|
| Legume | Rhizobium | ||
| Anthyllis vulneraria L. | Mesorhizobium sp. | Zn, Cd, Pb | [136,137] |
| Astragalus thaliana L. | - | Cd | [138] |
| Cytisus scoparius (L.) Link | - | Pb, Zn | [139] |
| Glycine max (L.) Merr | - | Cd | [140] |
| G. max (L.) Merr | Bradyrhizobium japonicum | As | [141] |
| Lablab purpurens | Rhizobium sp. | Cd, Cu, Zn | [142] |
| Lathyrus sativus L. | - | Pb | [143] |
| Lens culinaris Medik. | Rhizobium leguminosarum | Zn, Cu, Cd, Pb | [144,145] |
| Lotus corniculatus L. | Bradyrhizobium liaoningense | Zn, Pb | [146] |
| Lupinus albus L. | - | Cd, As, Cu, Pb, Zn | [147,148] |
| Lupinus luteus L. | Bradyrhizobium sp. | Cd, Cu, Pb, Zn | [149] |
| Lupinus sp. | Bradyrhizobium sp. | Cd, Cu, Pb, Zn | [133] |
| Medicago lupulina L | Sinorhizobium meliloti | Cu | [150] |
| Medicago sativa L. | Rhizobium leguminosarum bv. trifolii | Cr, Cu, Zn, Hg | [151,152] |
| M. sativa L. | Sinorhizobium meliloti | Cu | [153] |
| Pisum sativum L. | Rhizobium sp. | Cd, Cu, Zn | [154,155] |
| Prosopis laevigata (Willd.) M.C. Johnst. | - | Cu, Pb, Zn | [156] |
| Robinia pseudoacacia L. | - | Pb, Cu, Zn | [38,157,158] |
| Sesbania rostrata Bremek. & Oberm. | - | Cu, Pb, Zn | [159] |
| Sesbania sesban L. | - | Cu, Pb, Zn | [159] |
| Vicia faba L. | - | Pb, Zn, As, Cd, Cu | [160,161] |
| V. faba L. | Rhizobium leguminosarum | Cd, Cu, PB | [162] |
| Vicia sativa L. | - | Cd, Pb, Zn | [161] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jach, M.E.; Sajnaga, E.; Ziaja, M. Utilization of Legume-Nodule Bacterial Symbiosis in Phytoremediation of Heavy Metal-Contaminated Soils. Biology 2022, 11, 676. https://doi.org/10.3390/biology11050676
Jach ME, Sajnaga E, Ziaja M. Utilization of Legume-Nodule Bacterial Symbiosis in Phytoremediation of Heavy Metal-Contaminated Soils. Biology. 2022; 11(5):676. https://doi.org/10.3390/biology11050676
Chicago/Turabian StyleJach, Monika Elżbieta, Ewa Sajnaga, and Maria Ziaja. 2022. "Utilization of Legume-Nodule Bacterial Symbiosis in Phytoremediation of Heavy Metal-Contaminated Soils" Biology 11, no. 5: 676. https://doi.org/10.3390/biology11050676
APA StyleJach, M. E., Sajnaga, E., & Ziaja, M. (2022). Utilization of Legume-Nodule Bacterial Symbiosis in Phytoremediation of Heavy Metal-Contaminated Soils. Biology, 11(5), 676. https://doi.org/10.3390/biology11050676