
Effects of Bacillus subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site and Experimental Design
2.2. Sample Collection and Count of Culturable Microorganism
2.3. Determination of Edaphic Properties
2.4. DNA Isolation and PCR Amplification
2.5. Real-Time (q)PCR
2.6. Illumina MiSeq Sequencing
2.7. Processing of Sequencing Data
2.8. Statistical Analyses
3. Results
3.1. Changes in Poplar Tree DBH and Plant Height
3.2. Changes in Edaphic Properties from March 2018 to October 2020
3.3. Differences in Microbequantities in Rhizosphere Soils of Poplars
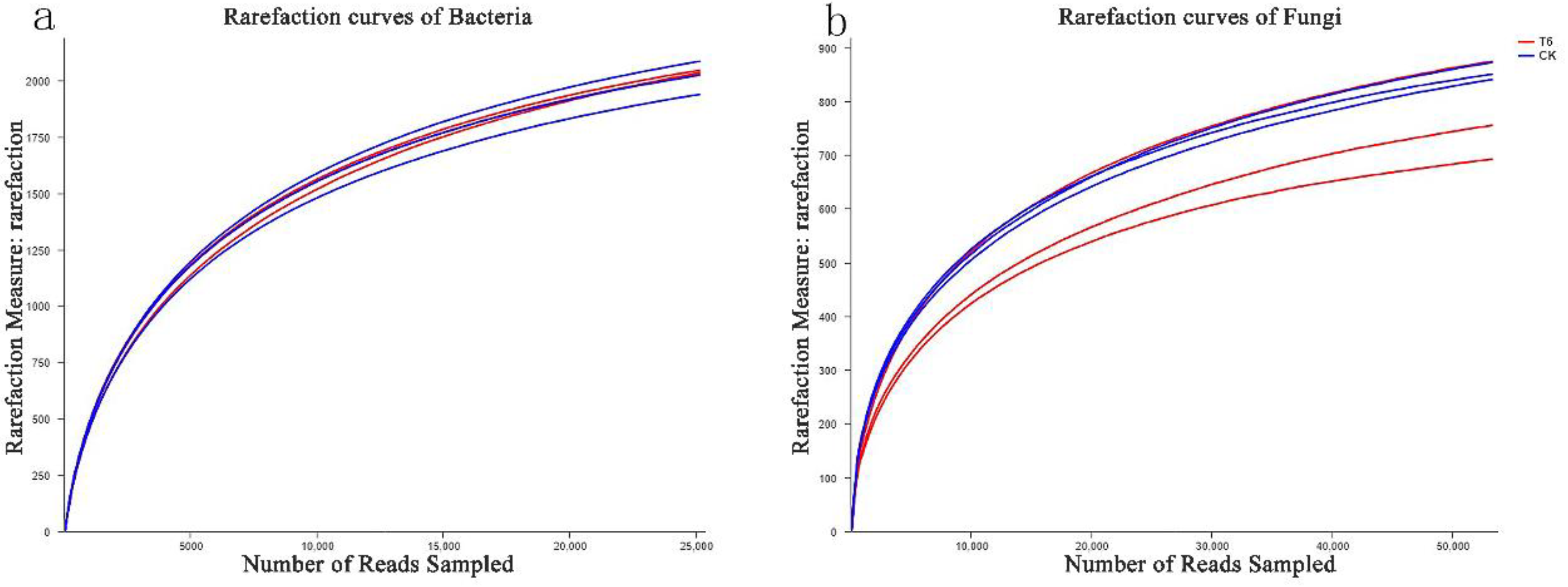
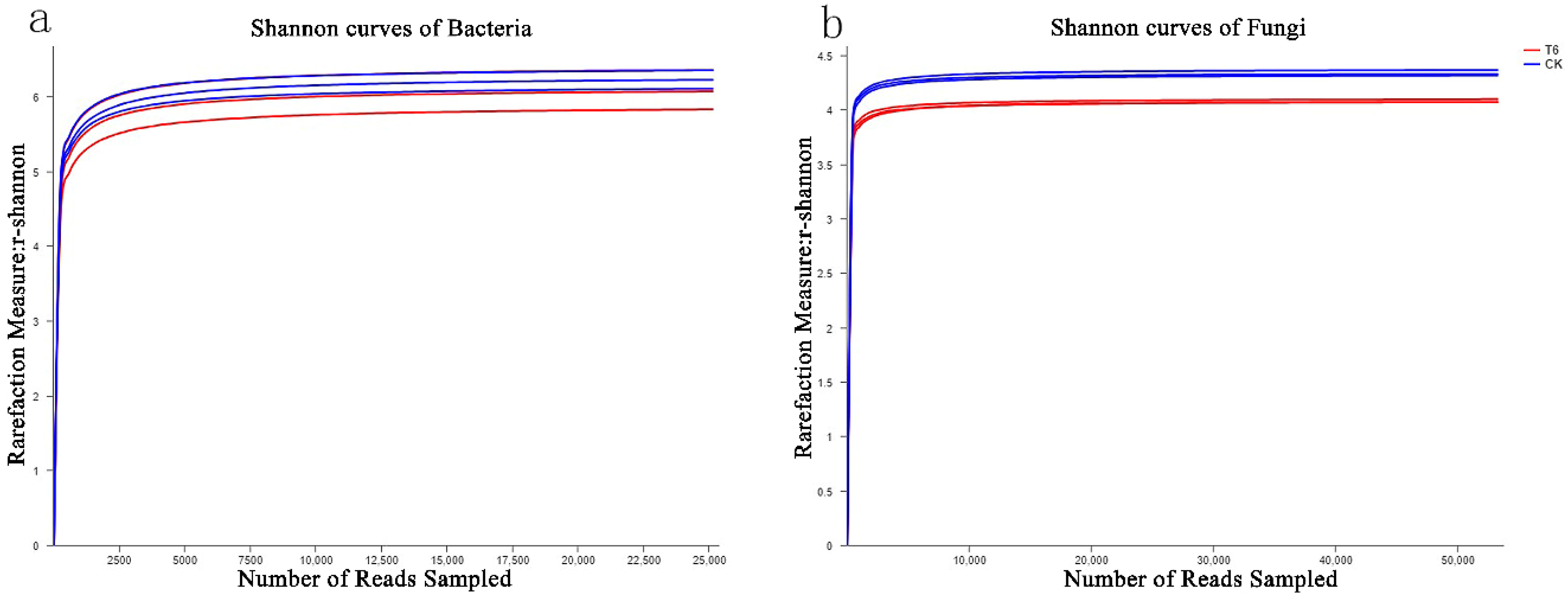
3.4. Estimators of the Diversity along with Species Richness of Microbial Communities
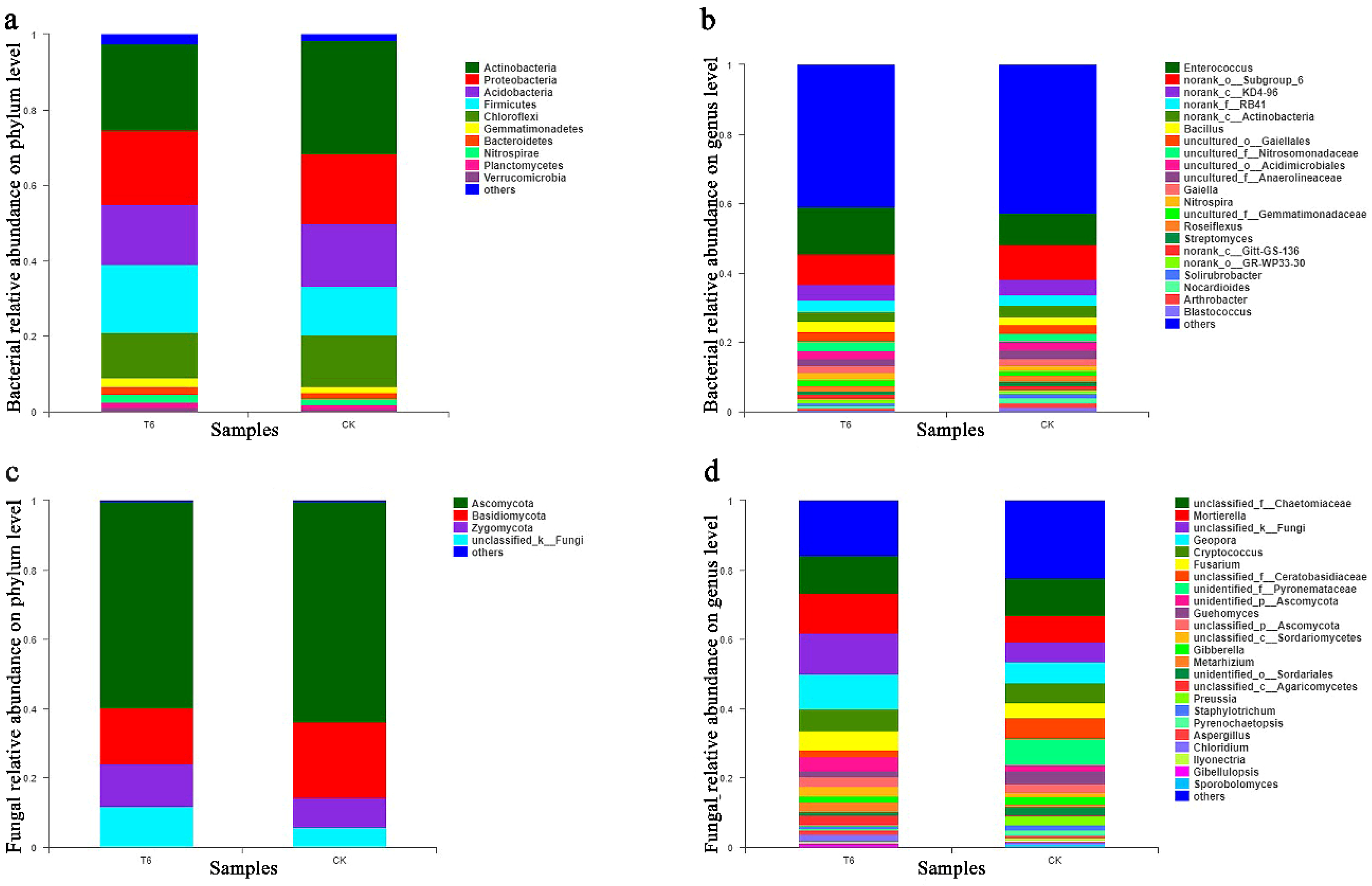
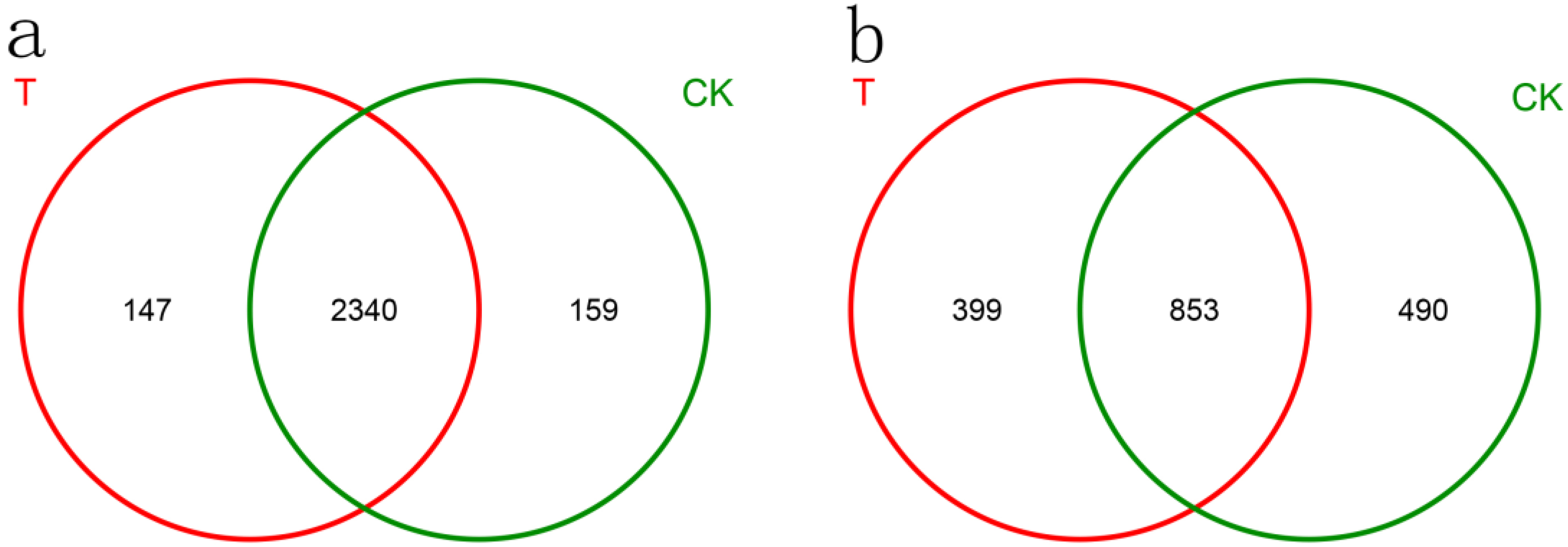
3.5. Effects of T6-1 on Community Composition and Structure
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilske, B.; Lu, N.; Wei, L.; Chen, S.; Zha, T.; Liu, C.; Xu, W.; Noormets, A.; Huang, J.; Wei, Y.; et al. Poplar plantation has the potential to alter the water balance in semiarid Inner Mongolia. J. Environ. Manag. 2009, 90, 2762–2770. [Google Scholar] [CrossRef]
- Xi, B.; Clothier, B.; Coleman, M.; Duan, J.; Hu, W.; Li, D.; Di, N.; Liu, Y.; Fu, J.; Li, J.; et al. Irrigation management in poplar (Populus spp.) plantations: A review. For. Ecol. Manag. 2021, 494, 119330. [Google Scholar] [CrossRef]
- Wang, Y.; Bai, G.; Shao, G.; Cao, Y. An analysis of potential investment returns and their determinants of poplar plantations in state-owned forest enterprises of China. New For. 2014, 45, 251–264. [Google Scholar] [CrossRef]
- Cheng, S.; Xu, Z.; Su, Y.; Zhen, L. Spatial and temporal flows of China’s forest resources: Development of a framework for evaluating resource efficiency. Ecol. Econ. 2010, 69, 1405–1415. [Google Scholar] [CrossRef]
- Xia, J.; Zhang, S.; Li, T.; Liu, X.; Zhang, R.; Zhang, G. Effect of Continuous Cropping Generations on Each Component Biomass of Poplar Seedlings during Different Growth Periods. Sci. World J. 2014, 2014, 618421. [Google Scholar] [CrossRef] [Green Version]
- Liu, F.; Jiang, Y.; Liu, Y.; Wang, H. Effects of continuous cropping on the growth of I-107 (Populus × Euramercana) poplar cutting stock. Acta Agric. Univ. Jiangxiensis 2004, 26, 681–685. [Google Scholar]
- Zhang, Z.L.; Wang, W.Q. Progress on formation mechanism and control measurements of continuous cropping obstacles in plants. J. Biol. 2010, 27, 69–72. [Google Scholar]
- Pang, Z.; Dong, F.; Liu, Q.; Lin, W.; Yuan, Z. Soil Metagenomics Reveals Effects of Continuous Sugarcane Cropping on the Structure and Functional Pathway of Rhizospheric Microbial Community. Front. Microbiol. 2021, 12, 627569. [Google Scholar] [CrossRef]
- Bresolin, J.D.; Bustamante, M.M.C.; Krüger, R.H.; Silva, M.R.S.S.; Perez, K.S. Structure and composition of bacterial and fungal community in soil under soybean monoculture in the Brazilian Cerrado. Braz. J. Microbiol. [Publ. Braz. Soc. Microbiol.] 2010, 41, 391. [Google Scholar] [CrossRef] [Green Version]
- Gu, Y.; Wang, Y.; Wang, P.; Wang, C.; Ma, J.; Yang, X.; Ma, D.; Li, M. Study on the Diversity of Fungal and Bacterial Communities in Continuous Cropping Fields of Chinese Chives (Allium tuberosum). BioMed Res. Int. 2020, 2020, 1–14. [Google Scholar] [CrossRef]
- He, W.; Guo, L.W.; Fan, P.H.; Guo, B.; Fu, Y.P.; Wei, Y.H. Allelopathy and the rhizospere bacterial community structure of Oxytropis ochrocephala. Acta Prataculturae Sin. 2015, 24, 21–29. [Google Scholar]
- Huang, Y.; Xiao, X.; Huang, H.; Jing, J.; Zhao, H.; Wang, L.; Long, X.-E. Contrasting beneficial and pathogenic microbial communities across consecutive cropping fields of greenhouse strawberry. Appl. Microbiol. Biotechnol. 2018, 102, 5717–5729. [Google Scholar] [CrossRef]
- Xiong, W.; Zhao, Q.; Zhao, J.; Xun, W.; Li, R.; Zhang, R.; Wu, H.; Shen, Q. Different Continuous Cropping Spans Significantly Affect Microbial Community Membership and Structure in a Vanilla-Grown Soil as Revealed by Deep Pyrosequencing. Microb. Ecol. 2014, 70, 209–218. [Google Scholar] [CrossRef]
- Lu, T.; Ke, M.; Lavoie, M.; Jin, Y.; Fan, X.; Zhang, Z.; Fu, Z.; Sun, L.; Gillings, M.; Peñuelas, J.; et al. Rhizosphere microorganisms can influence the timing of plant flowering. Microbiome 2018, 6, 231. [Google Scholar] [CrossRef]
- Yuan, J.; Zhao, J.; Wen, T.; Zhao, M.; Li, R.; Goossens, P.; Huang, Q.; Bai, Y.; Vivanco, J.M.; Kowalchuk, G.A.; et al. Root exudates drive the soil-borne legacy of aboveground pathogen infection. Microbiome 2018, 6, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.; Cui, Y.; Li, H.; Kuang, A.; Li, X.; Wei, Y.; Ji, X. Rhizospheric soil and root endogenous fungal diversity and composition in response to continuous Panax notoginseng cropping practices. Microbiol. Res. 2017, 194, 10–19. [Google Scholar] [CrossRef]
- Olanrewaju, O.S.; Ayilara, M.S.; Ayangbenro, A.S.; Babalola, O.O. Genome Mining of Three Plant Growth-PromotingBacillusSpecies from Maize Rhizosphere. Appl. Biochem. Biotechnol. 2021, 193, 3949–3969. [Google Scholar] [CrossRef]
- Qiao, J.; Yu, X.; Liang, X.; Liu, Y.; Borriss, R.; Liu, Y. Addition of plant-growth-promoting Bacillus subtilis PTS-394 on tomato rhizosphere has no durable impact on composition of root microbiome. BMC Microbiol. 2017, 17, 131. [Google Scholar] [CrossRef]
- Solanki, M.K.; Kumar, S.; Pandey, A.K.; Srivastava, S.; Singh, R.K.; Kashyap, P.L.; Srivastava, A.K.; Arora, D.K. Diversity and antagonistic potential of Bacillus spp. associated to the rhizosphere of tomato for the management of Rhizoctonia solani. Biocontrol Sci. Technol. 2012, 22, 203–217. [Google Scholar] [CrossRef]
- Qiu, M.; Zhang, R.; Xue, C.; Zhang, S.; Li, S.; Zhang, N.; Shen, Q. Application of bio-organic fertilizer can control Fusarium wilt of cucumber plants by regulating microbial community of rhizosphere soil. Biol. Fertil. Soils 2012, 48, 807–816. [Google Scholar] [CrossRef]
- Kröber, M.; Ewibberg, D.; Grosch, R.; Eikmeyer, F.G.; Verwaaijen, B.; Chowdhury, S.P.; Hartmann, A.E.; Pühler, A.; Schlüter, A. Effect of the strain Bacillus amyloliquefaciens FZB42 on the microbial community in the rhizosphere of lettuce under field conditions analyzed by whole metagenome sequencing. Front. Microbiol. 2014, 5, 252. [Google Scholar]
- Cai, F.; Pang, G.; Li, R.-X.; Li, R.; Gu, X.-L.; Shen, Q.-R.; Chen, W. Bioorganic fertilizer maintains a more stable soil microbiome than chemical fertilizer for monocropping. Biol. Fertil. Soils 2017, 53, 861–872. [Google Scholar] [CrossRef]
- Chowdhury, S.P.; Dietel, K.; Rändler, M.; Schmid, M.; Junge, H.; Borriss, R.; Hartmann, A.; Grosch, R. Effects of Bacillus amyloliquefaciens FZB42 on lettuce growth and health under pathogen pressure and its impact on the rhizosphere bacterial community. PLoS ONE 2013, 8, e68818. [Google Scholar] [CrossRef] [Green Version]
- Smalla, K.; Cresswell, N.; Mendonca‐Hagler, L.C.; Wolters, A.; Elsas, J.V. Rapid DNA extraction protocol from soil for polymerase chain reaction-mediated amplification. J. Appl. Microbiol. 1993, 74, 78–85. [Google Scholar] [CrossRef]
- Bonito, G.; Hameed, K.; Ventura, R.; Krishnan, J.; Schadt, C.; Vilgalys, R. Isolating a functionally relevant guild of fungi from the root microbiome of Populus. Fungal Ecol. 2016, 22, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Walkley, A.; Black, I.A. An examination of the Degtjareff method for determining soil organic matter and a proposed modification to the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Olsen, S.R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate. Misc. Pap. Inst. Agric. Res. Samaru Pap. 1954, 939, 18–19. [Google Scholar]
- Peng, W.; Bo, C.; Hua, Z. High throughput sequencing analysis of bacterial communities in soils of a typical Poyang Lake wetland. Acta Ecol. Sin. 2017, 37, 1650–1658. [Google Scholar]
- Xin, L.I.; Wang, Y.H.; Lin, J.Y.; Liu, L.S.; Huang, Z.X. Analysis of Microbial Diversity in the Fermented Grains of Maotai-flavor Liquor Using High-throughput Sequencing. J. Fujian Norm. Univ. 2017, 11, 51–59. [Google Scholar]
- Hao, Y.T.; Wu, S.G.; Xiong, F.; Tran, N.T.; Jakovlić, I.; Zou, H.; Li, W.X.; Wang, G.T. Succession and Fermentation Products of Grass Carp (Ctenopharyngodon idellus) Hindgut Microbiota in Response to an Extreme Dietary Shift. Front. Microbiol. 2017, 8, 1585. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.; Qiu, T.; Sun, Y.; Wang, X. The abundance and diversity of antibiotic resistance genes in the atmospheric environment of composting plants. Environ. Int. 2018, 116, 229. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amato, K.R.; Yeoman, C.J.; Kent, A.; Righini, N.; Carbonero, F.; Estrada, A.; Gaskins, H.R.; Stumpf, R.M.; Yildirim, S.; Torralba, M.; et al. Habitat degradation impacts black howler monkey (Alouatta pigra) gastrointestinal microbiomes. ISME J. 2013, 7, 1344–1353. [Google Scholar] [CrossRef]
- Abarenkov, K.; Nilsson, R.H.; Larsson, K.-H.; Alexander, I.J.; Eberhardt, U.; Erland, S.; Høiland, K.; Kjøller, R.; Larsson, E.; Pennanen, T.; et al. The UNITE database for molecular identification of fungi—Recent updates and future perspectives. New Phytol. 2010, 186, 281. [Google Scholar] [CrossRef]
- Shi, G.; Sun, H.; Calderón‐Urrea, A.; Li, M.; Yang, H.; Wang, W.; Su, G. Bacterial communities as indicators for soil health under continuous cropping system. Land Degrad. Dev. 2021, 32, 2393–2408. [Google Scholar] [CrossRef]
- Manici, L.M.; Kelderer, M.; Franke-Whittle, I.; Rühmer, T.; Baab, G.; Nicoletti, F.; Caputo, F.; Topp, A.; Insam, H.; Naef, A. Relationship between root-endophytic microbial communities and replant disease in specialized apple growing areas in Europe. Appl. Soil Ecol. 2013, 72, 207–214. [Google Scholar] [CrossRef]
- Li, X.; Lewis, E.E.; Liu, Q.; Li, H.; Bai, C.; Wang, Y. Effects of long-term continuous cropping on soil nematode community and soil condition associated with replant problem in strawberry habitat. Sci. Rep. 2016, 6, 30466. [Google Scholar] [CrossRef] [Green Version]
- Mena‐Violante, H.G.; Olalde-Portugal, V. Alteration of tomato fruit quality by root inoculation with plant growth-promoting rhizobacteria (PGPR): Bacillus subtilis BEB-13bs. Sci. Hortic. 2007, 113, 103–106. [Google Scholar] [CrossRef]
- Feng-Zhi, W.U.; Wang, X.Z. Effect of Monocropping and Rotation on Soil Microbial Community Diversity and Cucumber Yield, Quality Under Protected Cultivation. Sci. Agric. Sin. 2007, 761, 555–561. [Google Scholar]
- Paudel, B.R.; Carpenter-Boggs, L.; Higgins, S. Influence of brassicaceous soil amendments on potentially beneficial and pathogenic soil microorganisms and seedling growth in Douglas-fir nurseries. Appl. Soil Ecol. 2016, 105, 91–100. [Google Scholar] [CrossRef] [Green Version]
- Patkowska, E. Biostimulants Managed Fungal Phytopathogens and Enhanced Activity of Beneficial Microorganisms in Rhizosphere of Scorzonera (Scorzonera hispanica L.). Agriculture 2021, 11, 347. [Google Scholar] [CrossRef]
- Janvier, C.; Villeneuve, F.; Alabouvette, C.; Edel-Hermann, V.; Mateille, T.; Steinberg, C. Soil health through soil disease suppression: Which strategy from descriptors to indicators? Soil Biol. Biochem. 2007, 39, 1–23. [Google Scholar] [CrossRef]
- Jayasinghe, B.; Parkinson, D. Actinomycetes as antagonists of litter decomposer fungi. Appl. Soil Ecol. 2008, 38, 109–118. [Google Scholar] [CrossRef]
- Muangham, S.; Pathom‐Aree, W.; Duangmal, K. Melanogenic actinomycetes from rhizosphere soil-antagonistic activity against Xanthomonas oryzae and plant-growth-promoting traits. Can. J. Microbiol. 2015, 61, 164. [Google Scholar] [CrossRef] [PubMed]
- Soltanzadeh, M.; Soltani Nejad, M.; Shahidi Bonjar, G.H. Application of Soil‐borne Actinomycetes for Biological Control against Fusarium Wilt of Chickpea (Cicer arietinum) caused by Fusarium solani fsp pisi. J. Phytopathol. 2016, 164, 967–978. [Google Scholar] [CrossRef]
- Lee, S.H.; Ka, J.O.; Cho, J.C. Members of the phylum Acidobacteria are dominant and metabolically active in rhizosphere soil. FEMS Microbiol. Lett. 2010, 285, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Giguere, A.T.; Eichorst, S.A.; Meier, D.V.; Herbold, C.W.; Richter, A.; Greening, C.; Woebken, D. Acidobacteria are active and abundant members of diverse atmospheric H2 -oxidizing communities detected in temperate soils. ISME J. 2020, 15, 363–367. [Google Scholar] [CrossRef]
- Sun, H.; Terhonen, E.; Koskinen, K.; Paulin, L.; Kasanen, R.; Asiegbu, F.O. Bacterial diversity and community structure along different peat soils in boreal forest. Appl. Soil Ecol. 2014, 74, 37–45. [Google Scholar] [CrossRef]
- Qiu, Z.; Shi, C.; Zhao, M.; Wang, K.; Zhang, M.; Wang, T.; Shi, F. Improving Effects of Afforestation with Different Forest Types on Soil Nutrients and Bacterial Community in Barren Hills of North China. Sustainability 2022, 14, 1202. [Google Scholar] [CrossRef]
- Szczech, M.; Shoda, M. The Effect of Mode of Application of Bacillus subtilis RB14-C on its Efficacy as a Biocontrol Agent Against Rhizoctonia solani. J. Phytopathol. 2010, 154, 370–377. [Google Scholar] [CrossRef]
- Zhou, X.; Wu, F. Dynamics of the diversity of fungal and Fusarium communities during continuous cropping of cucumber in the greenhouse. FEMS Microbiol. Ecol. 2012, 80, 469–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, L.; Cui, J.; Jie, W.; Cai, B. Analysis of the community compositions of rhizosphere fungi in soybeans continuous cropping fields. Microbiol Res. 2015, 180, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Khaledi, N.; Taheri, P.; Tarighi, S. Antifungal activity of various essential oils against Rhizoctonia solani and Macrophomina phaseolina as major bean pathogens. J. Appl. Microbiol. 2015, 118, 704–717. [Google Scholar] [CrossRef] [PubMed]
- Yugander, A.; Ladhalakshmi, D.; Prakasham, V.; Mangrauthia, S.K.; Prasad, M.S.; Krishnaveni, D.; Madhav, M.S.; Sundaram, R.M.; Laha, G.S. Pathogenic and Genetic Variation among the Isolates of Rhizoctonia solani (AG 1-IA), the Rice Sheath Blight Pathogen. J. Phytopathol. 2015, 163, 465–474. [Google Scholar] [CrossRef]
- Scherwinski, K.; Grosch, R.; Berg, G. Effect of bacterial antagonists on lettuce: Active biocontrol of Rhizoctonia solani and negligible, short-term effects on nontarget microorganisms. FEMS Microbiol. Ecol. 2010, 64, 106–116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asaka, O.; Shoda, M. Biocontrol of Rhizoctonia solani Damping-Off of Tomato with Bacillus subtilis RB14. Appl. Environ. Microbiol. 1996, 62, 4081–4085. [Google Scholar] [CrossRef] [Green Version]
- Donn, S.; Almario, J.; Muller, D.; Moënne-Loccoz, Y.; Gupta, V.V.; Kirkegaard, J.A.; Richardson, A.E. Rhizosphere microbial communities associated with Rhizoctonia damage at the field and disease patch scale. Appl. Soil Ecol. 2014, 78, 37–47. [Google Scholar] [CrossRef]
- Li, L.; Ma, J.; Ibekwe, A.M.; Wang, Q.; Yang, C.-H. Cucumber Rhizosphere Microbial Community Response to Biocontrol Agent Bacillus subtilis B068150. Agriculture 2015, 6, 2–18. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Ma, J.; Ibekwe, A.M.; Wang, Q.; Yang, C.-H. Influence of Bacillus subtilis B068150 on cucumber rhizosphere microbial composition as a plant protective agent. Plant Soil 2018, 429, 519–531. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | DBH (mm) in 2018/10 | DBH (mm) in 2020/10 | Plant Height (m) in 2018/10 | Plant Height (m) in 2020/10 |
|---|---|---|---|---|
| T6-1 | 38.54 ± 3.88 a | 124.16 ± 9.06 a | 4.15 ± 0.36 a | 11.34 ± 0.62 a |
| CK | 35.23 ± 4.12 b | 111.13 ± 8.84 b | 3.99 ± 0.30 a | 10.73 ± 0.48 b |
| OC (g/kg) | AP (mg/kg) | AK (mg/kg) | TN (mg/kg) | pH | ||
|---|---|---|---|---|---|---|
| Mar-13 | T6-1 | 6.12 ± 0.08 c | 35.91 ± 4.24 b | 132.06 ± 2.38 c | 758.98 ± 9.26 b | 7.19 ± 0.02 b |
| CK | 6.05 ± 0.07 c | 35.63 ± 2.33 b | 132.28 ± 1.44 c | 765.82 ± 7.43 b | 7.23 ± 0.02 b | |
| Oct-15 | T6-1 | 7.12 ± 0.03 a | 46.72 ± 3.02 a | 231.79 ± 4.23 a | 1092.49 ± 44.79 a | 7.39 ± 0.02 a |
| CK | 6.27 ± 0.04 b | 35.38 ± 2.77 b | 139.20 ± 2.03 b | 791.58 ± 3.77 b | 7.07 ± 0.04 c |
| Treatment | Culturable Microbial Contents after the Two Treatments | Total Microbial Contents after the Two Treatments | ||
|---|---|---|---|---|
| Bacterial × 107 (cfu/g Soil) | Fungal × 106 (cfu/g Soil) | Bacterial × 107 (Copies/μL) | Fungal × 104 (Copies/μL) | |
| T6-1 | 9.71 ± 0.12 a | 5.00 ± 0.25 b | 1.17 ± 0.11 a | 2.19 ± 0.01 b |
| CK | 0.96 ± 0.17 b | 9.25 ± 0.49 a | 0.71 ± 0.02 b | 2.95 ± 0.10 a |
| Sample | Cutoff | OTUs | ACE | Chao | Shannon | Simpson | Coverage | |
|---|---|---|---|---|---|---|---|---|
| Bacterial | T6-1 | 0.03 | 2161 ± 44.84 a | 2362.75 ± 68.25 a | 2362.00 ± 102.20 a | 6.06 ± 0.23 a | 0.023 ± 0.013 a | 0.980169 |
| CK | 0.03 | 2106 ± 106.71 a | 2384.41 ± 82.6 a | 2402.16 ± 78.39 a | 6.20 ± 0.12 a | 0.012 ± 0.004 b | 0.980821 | |
| Fungal | T6-1 | 0.03 | 829.33 ± 96.45 a | 923.56 ± 90.50 b | 919.39 ± 93.32 b | 4.08 ± 0.02 b | 0.045 ± 0.007 a | 0.9967853 |
| CK | 0.03 | 901 ± 7.55 a | 1032.86 ± 57.48 a | 1022.94 ± 93.32 a | 4.34 ± 0.02 a | 0.040 ± 0.005 a | 0.996280 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sui, J.; Yu, Q.; Yang, K.; Yang, J.; Li, C.; Liu, X. Effects of Bacillus subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar. Biology 2022, 11, 791. https://doi.org/10.3390/biology11050791
Sui J, Yu Q, Yang K, Yang J, Li C, Liu X. Effects of Bacillus subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar. Biology. 2022; 11(5):791. https://doi.org/10.3390/biology11050791
Chicago/Turabian StyleSui, Junkang, Qianqian Yu, Kai Yang, Jiayi Yang, Chenyu Li, and Xunli Liu. 2022. "Effects of Bacillus subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar" Biology 11, no. 5: 791. https://doi.org/10.3390/biology11050791
APA StyleSui, J., Yu, Q., Yang, K., Yang, J., Li, C., & Liu, X. (2022). Effects of Bacillus subtilis T6-1 on the Rhizosphere Microbial Community Structure of Continuous Cropping Poplar. Biology, 11(5), 791. https://doi.org/10.3390/biology11050791