
Factors Affecting Roadkills of Semi-Aquatic Mammals





Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
3. Results
3.1. Species Composition of the Wild Mammal Roadkills
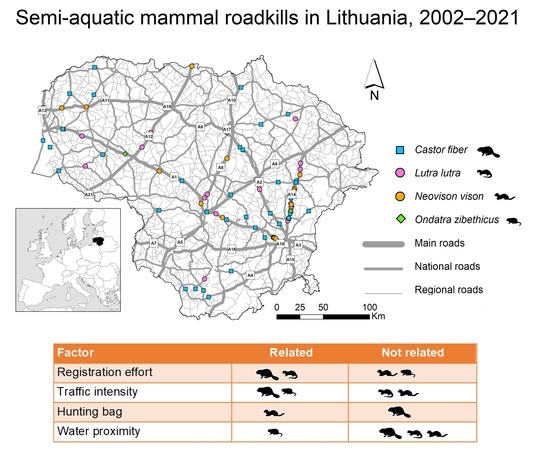
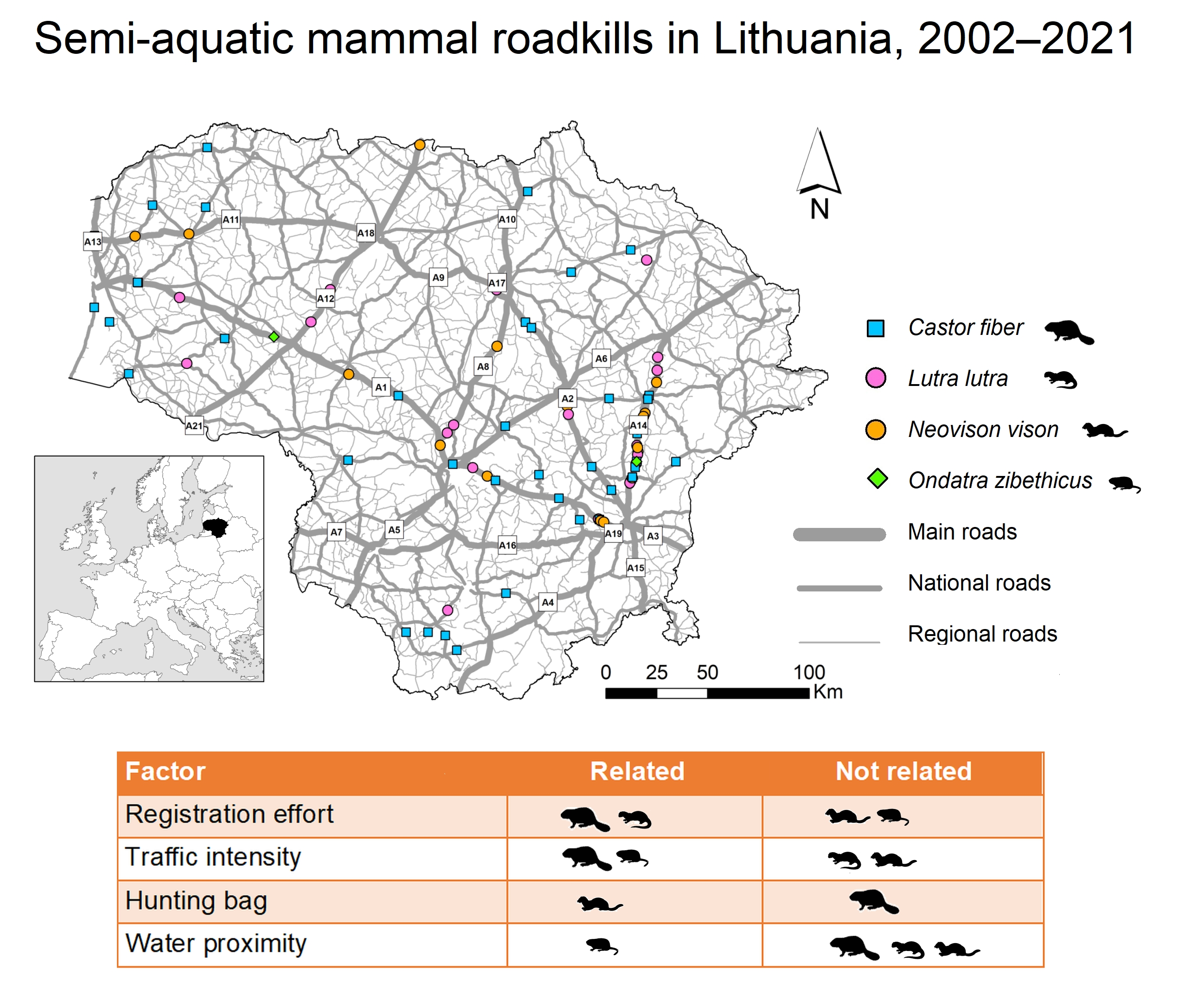
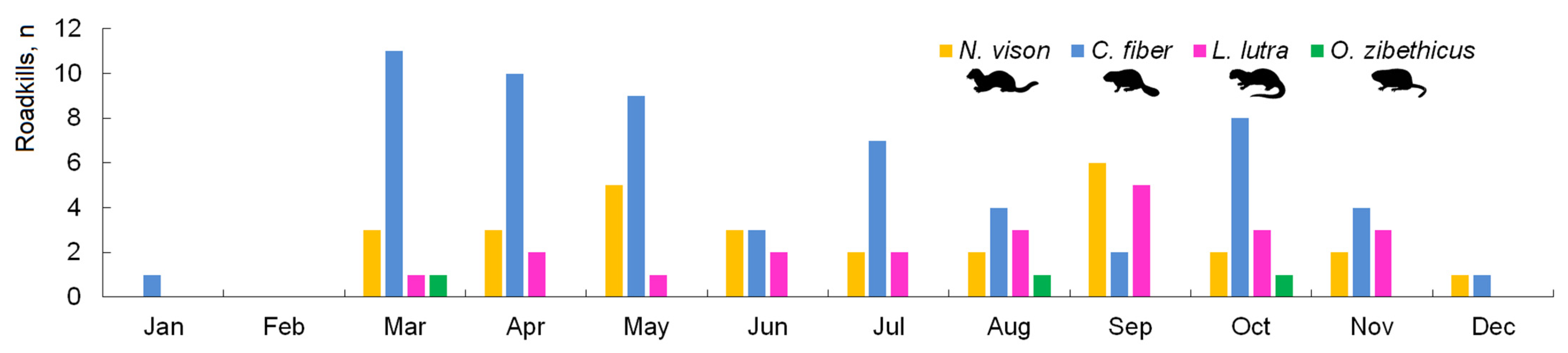
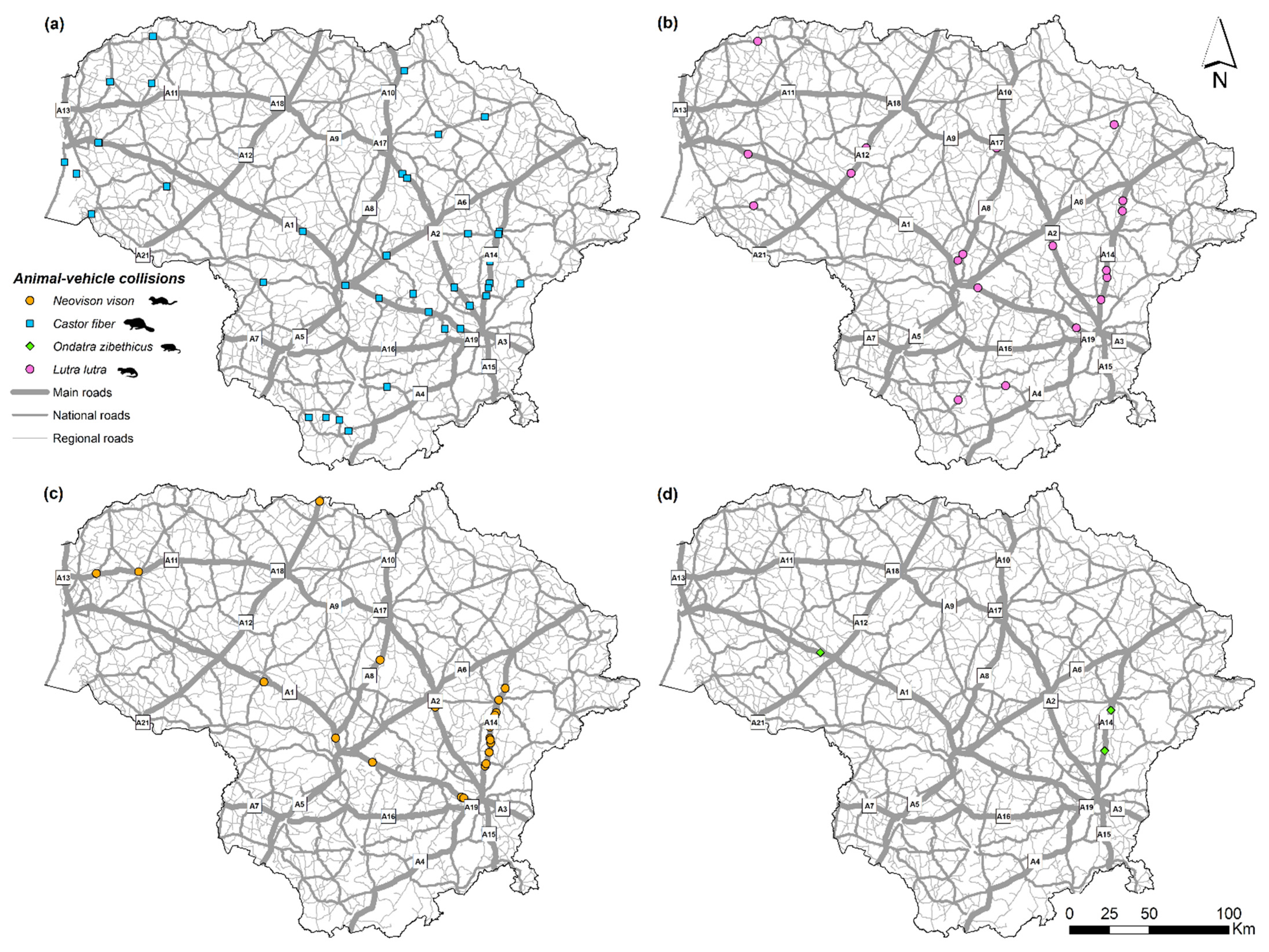
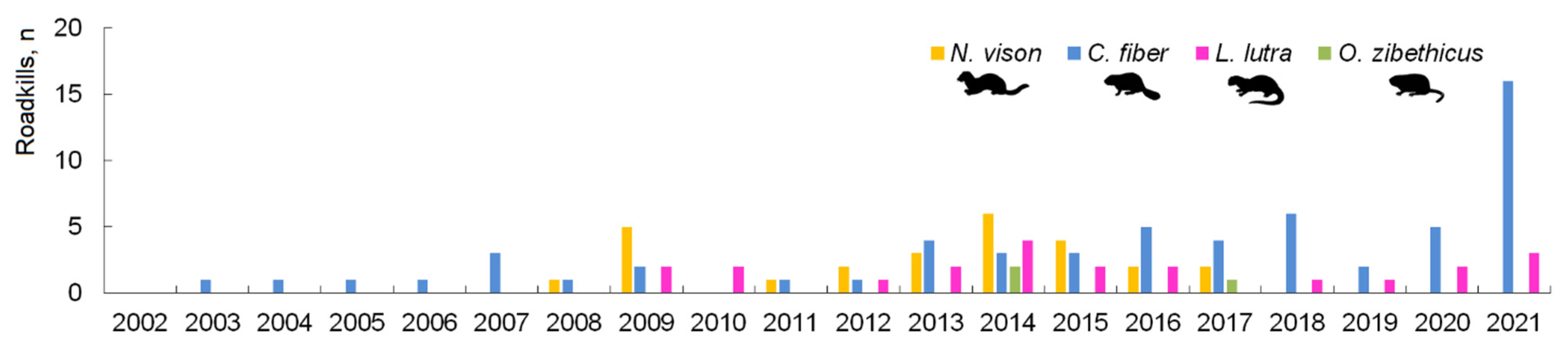
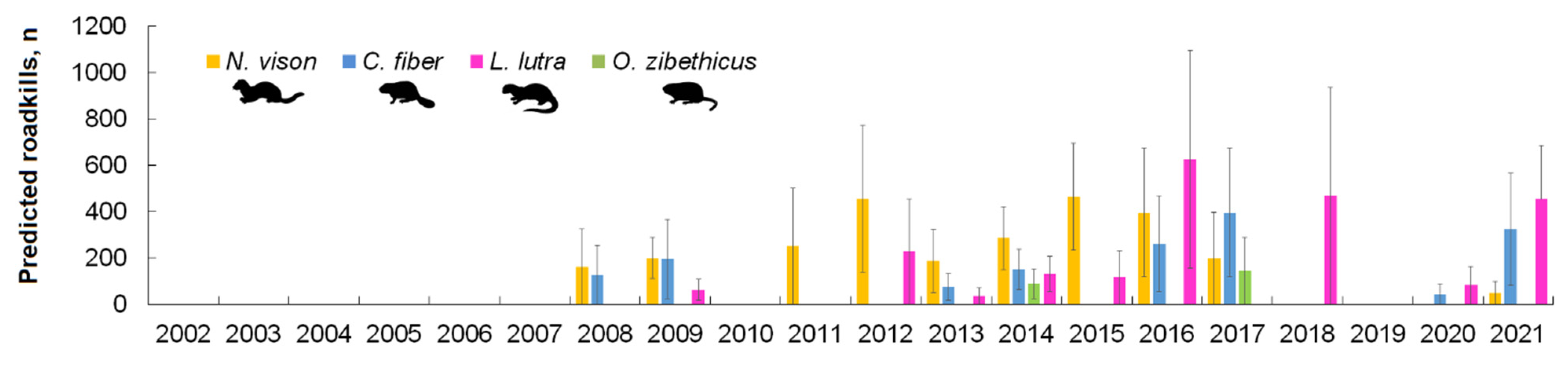
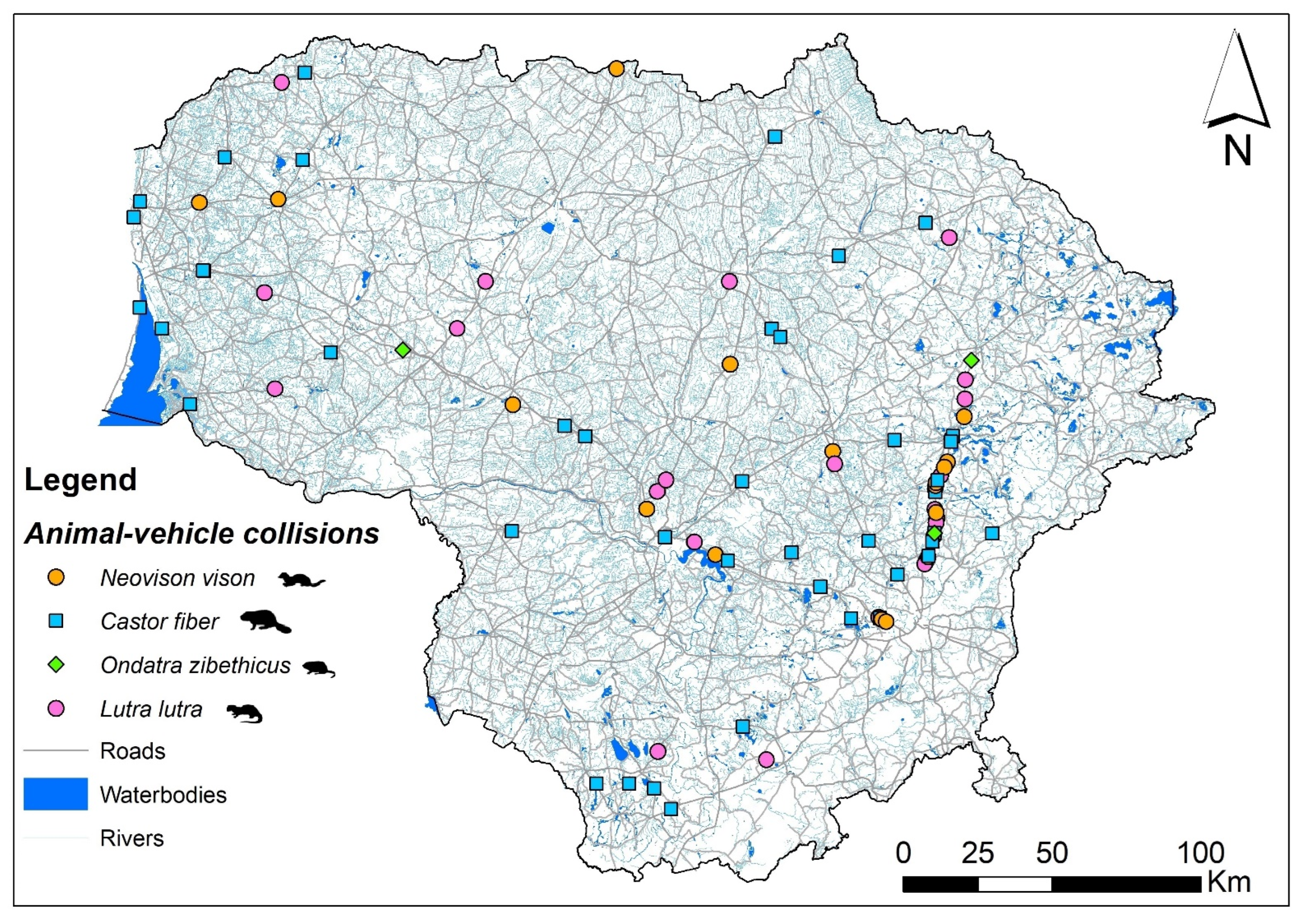
3.2. Spatiotemporal Distribution and Index of Semiaquatic Mammal Roadkill
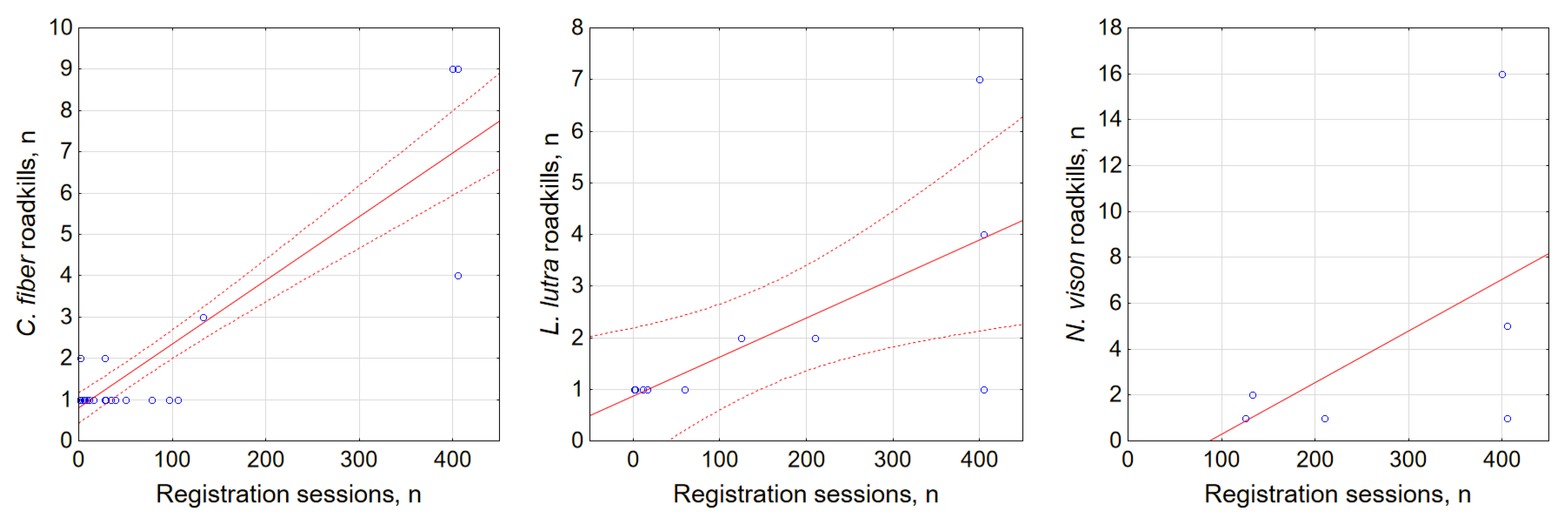
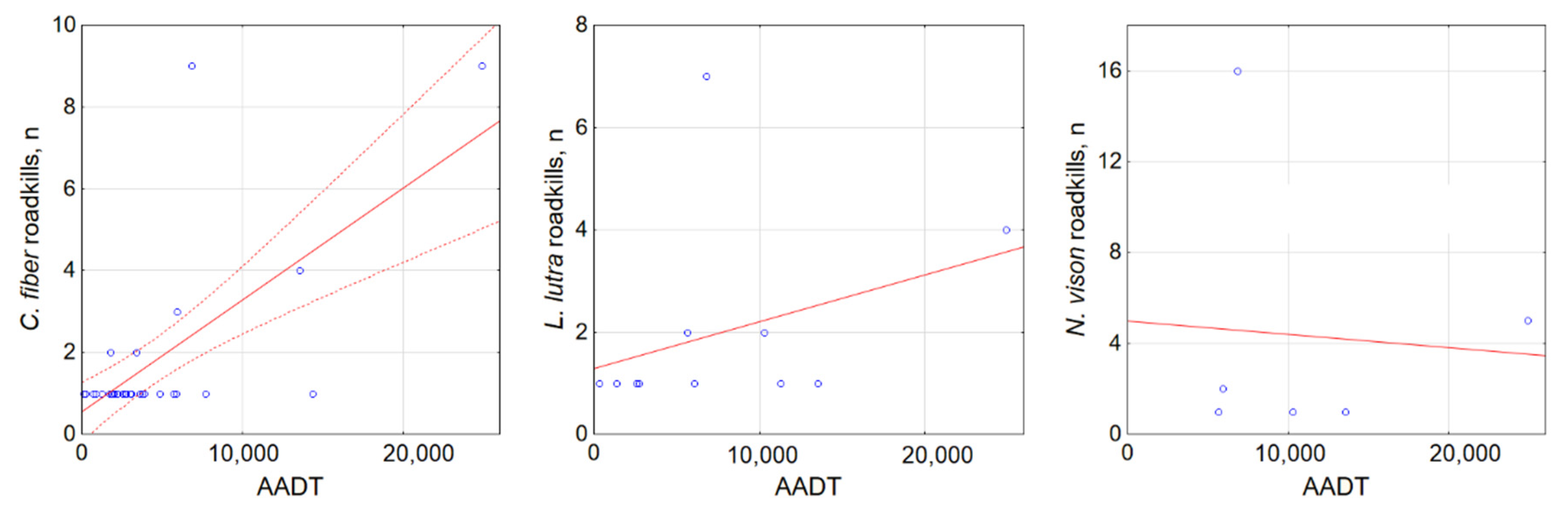
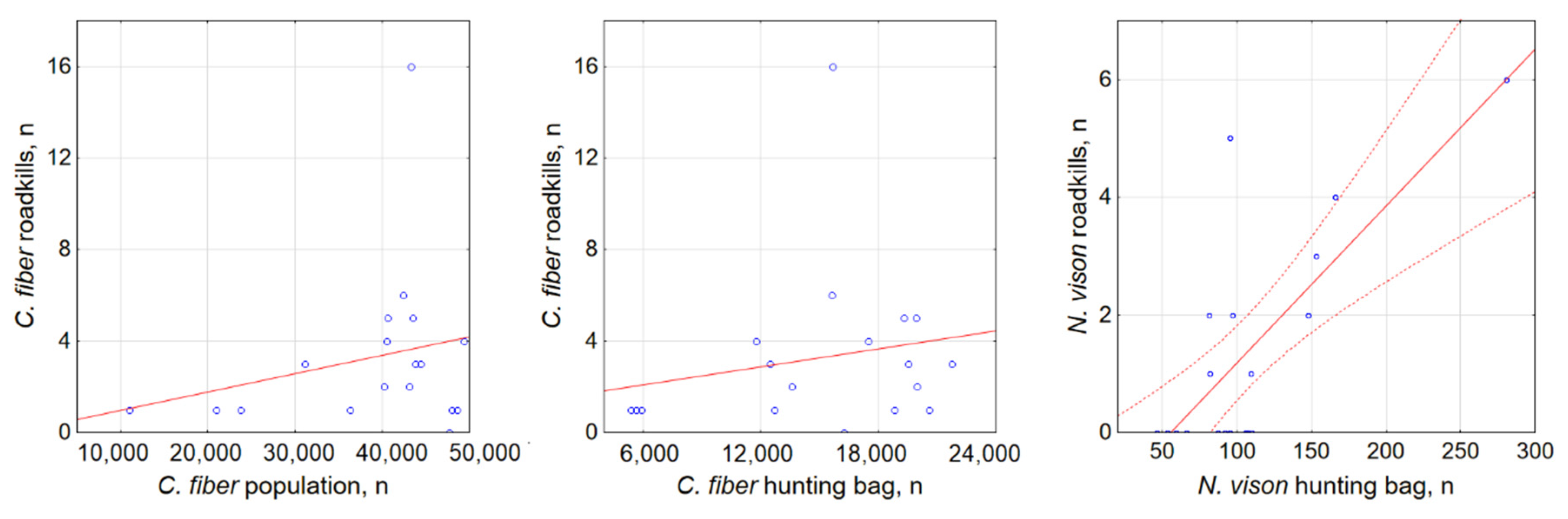
3.3. Factors Affecting Semi-Aquatic Mammal Roadkills
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Miljutin, A. Substrate utilization and feeding strategies of mammals: Description and classification. Est. J. Ecol. 2009, 58, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Hood, G.A. Semi-Aquatic Mammals: Ecology and Biology; JHU Press: Baltimore, MD, USA, 2020. [Google Scholar]
- Ulevičius, A.; Balčiauskas, L. Spatial relations among semi-aquatic mammals on the riverside. Acta Zool. Litu. 1999, 9, 42–48. [Google Scholar] [CrossRef]
- Hood, G.A. Accessing and mobilizing “new” data to evaluate emerging environmental impacts on semi-aquatic mammals. Land 2020, 9, 345. [Google Scholar] [CrossRef]
- Jones, C.G.; Lawton, J.H.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Nummi, P.; Liao, W.; Huet, O.; Scarpulla, E.; Sundell, J. The beaver facilitates species richness and abundance of terrestrial and semi-aquatic mammals. Glob. Ecol. Cons. 2019, 20, e00701. [Google Scholar] [CrossRef]
- Mortensen, R.M.; Reinhardt, S.; Hjønnevåg, M.E.; Wilson, R.P.; Rosell, F. Aquatic habitat use in a semi-aquatic mammal: The Eurasian beaver. Anim. Biotelem. 2021, 9, 1–19. [Google Scholar] [CrossRef]
- Hoffer, D.M.; Nielsen, C.; Rutter, A.; Anile, S. Occupancy of Semi-Aquatic Mammals in an Urban Landscape; Research Square preprint; Southern Illinois University: Carbondale, IL, USA, 2021. [Google Scholar] [CrossRef]
- Ulevičius, A.; Kisielytė, N.; Jasiulionis, M. Habitat use and selectivity by beavers (Castor fiber) in anthropogenic landscape. Ekologija 2011, 57, 47–54. [Google Scholar] [CrossRef] [Green Version]
- Balčiauskas, L.; Trakimas, G.; Juškaitis, R.; Ulevičius, A.; Balčiauskienė, L. Atlas of Lithuanian Mammals, Amphibians and Reptiles, 2nd ed.; Akstis: Vilnius, Lithuania, 1999. [Google Scholar]
- Atlas of Lithuanian Mammals. Available online: https://gamtostyrimai.lt/en/users/viewGroup/id.57/pageId.23 (accessed on 10 February 2022).
- Baltrūnaitė, L.; Balčiauskas, L.; Matulaitis, R.; Stirkė, V. Otter distribution in Lithuania in 2008 and changes in the last decade. Est. J. Ecol. 2009, 58, 94–102. [Google Scholar] [CrossRef] [Green Version]
- Ulevičius, A.; Balčiauskas, L. Changes in the territory use by semi-aquatic mammals in some of rivers in Lithuania. Acta Zool. Litu. 2002, 12, 151–158. [Google Scholar] [CrossRef]
- Arbačiauskas, K.; Augutis, D.; Balčiauskas, L.; Bastytė-Cseh, D. Red Data Book of Lithuania. Animals, Plants, Fungi; Rašomavičius, V., Ed.; Lietuvos Respublikos Aplinkos Ministerija: Vilnius, Lietuva, 2021; pp. 1–684. [Google Scholar]
- Krukowicz, T.; Firląg, K.; Chrobot, P. Spatiotemporal Analysis of Road Crashes with Animals in Poland. Sustainability 2022, 14, 1253. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Stratford, J.; Balčiauskienė, L.; Kučas, A. Importance of professional roadkill data in assessing diversity of mammal roadkills. Transp. Res. Part D Transp. Environ. 2020, 87, 102493. [Google Scholar] [CrossRef]
- González-Gallina, A.; Benítez-Badillo, G.; Rojas-Soto, O.R.; Hidalgo-Mihart, M.G. The small, the forgotten and the dead: Highway impact on vertebrates and its implications for mitigation strategies. Biodivers. Conserv. 2013, 22, 325–342. [Google Scholar] [CrossRef]
- Ortega, A.; Weiler, A. Diversity of vertebrates affected by roadkill and areas of greatest impact in Ruta III General Elizardo Aquino, Paraguay. Reportes Científicos de la FACEN 2018, 9, 31–41. [Google Scholar] [CrossRef]
- Abra, F.D.; da Costa Canena, A.; Garbino, G.S.T.; Medici, E.P. Use of unfenced highway underpasses by lowland tapirs and other medium and large mammals in central-western Brazil. Perspect. Ecol. Conserv. 2020, 18, 247–256. [Google Scholar] [CrossRef]
- Ferreguetti, A.C.; Graciano, J.M.; Luppi, A.P.; Pereira-Ribeiro, J.; Rocha, C.F.D.; Bergallo, H.G. Roadkill of medium to large mammals along a Brazilian road (BR-262) in Southeastern Brazil: Spatial distribution and seasonal variation. Stud. Neotrop. Fauna Environ. 2020, 55, 216–225. [Google Scholar] [CrossRef]
- Prist, P.R.; Garbino, G.S.; Abra, F.D.; Pagotto, T.; Giacon, O.O. Use of highway culverts by the water opossum (Chironectes minimus) in southeastern Brazil. Biota Neotrop. 2020, 20, e20201058. [Google Scholar] [CrossRef]
- Canova, L.; Balestrieri, A. Long-term monitoring by roadkill counts of mammal populations living in intensively cultivated landscapes. Biodivers. Conserv. 2019, 28, 97–113. [Google Scholar] [CrossRef]
- Schwartz, A.L.; Shilling, F.M.; Perkins, S.E. The value of monitoring wildlife roadkill. Eur. J. Wildl. Res. 2020, 66, 18. [Google Scholar] [CrossRef] [Green Version]
- Kučas, A.; Balčiauskas, L. Impact of Road Fencing on Ungulate-Vehicle Collisions and Hotspot Patterns. Land 2021, 10, 338. [Google Scholar] [CrossRef]
- Rodríguez-Morales, B.; Diaz-Varela, E.; Marey-Perez, M. Spatiotemporal Analysis of Vehicle Collisions Involving Wild Boar and Roe Deer in NW Spain. Accid. Anal. Prev. 2013, 60, 121–133. [Google Scholar] [CrossRef]
- D’Amico, M.; Roman, J.; de los Reyes, L.; Revilla, E. Vertebrate Road-Kill Patterns in Mediterranean Habitats: Who, When and Where. Biol. Conserv. 2015, 191, 234–242. [Google Scholar] [CrossRef]
- Bíl, M.; Kubeček, J.; Sedoník, J.; Andrášik, R. Srazenazver.Cz: A System for Evidence of Animal-Vehicle Collisions along Transportation Networks. Biol. Conserv. 2017, 213, 167–174. [Google Scholar] [CrossRef]
- Sullivan, J.M. Trends and Characteristics of Animal-Vehicle Collisions in the United States. J. Saf. Res. 2011, 42, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Ascensão, F.; Yogui, D.; Alves, M.; Medici, E.P.; Desbiez, A. Predicting Spatiotemporal Patterns of Road Mortality for Medium-Large Mammals. J. Environ. Manag. 2019, 248, 109320. [Google Scholar] [CrossRef] [PubMed]
- Pagany, R. Wildlife-Vehicle Collisions—Influencing Factors, Data Collection and Research Methods. Biol. Conserv. 2020, 251, 108758. [Google Scholar] [CrossRef]
- Kušta, T.; Keken, Z.; Ježek, M.; Holá, M.; Šmíd, P. The Effect of Traffic Intensity and Animal Activity on Probability of Ungulate-Vehicle Collisions in the Czech Republic. Saf. Sci. 2017, 91, 105–113. [Google Scholar] [CrossRef]
- Bramati, P.S.; Heinert, L.F.; Narloch, L.B.; Hostetter, J.; Finkielman, J.D. Animal-Related Motorcycle Collisions in North Dakota. Wilderness Environ. Med. 2012, 23, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Collinson, W.J.; Marneweck, C.; Davies-Mostert, H.T. Protecting the protected: Reducing wildlife roadkill in protected areas. Anim. Conserv. 2019, 22, 396–403. [Google Scholar] [CrossRef]
- Grilo, C.; Borda-de-Água, L.; Beja, P.; Goolsby, E.; Soanes, K.; le Roux, A.; Koroleva, E.; Ferreira, F.Z.; Gagné, S.A.; Wang, Y.; et al. Conservation threats from roadkill in the global road network. Glob. Ecol. Biogeogr. 2021, 30, 2200–2210. [Google Scholar] [CrossRef]
- Clevenger, A.; Chruszcz, B.; Gunson, K. Spatial Patterns and Factors Influencing Small Vertebrate Fauna Road-Kill Aggregations. Biol. Conserv. 2003, 109, 15–26. [Google Scholar] [CrossRef]
- Keken, Z.; Sedoník, J.; Kušta, T.; Andrášik, R.; Bíl, M. Roadside Vegetation Influences Clustering of Ungulate Vehicle Collisions. Transp. Res. Part D Transp. Environ. 2019, 73, 381–390. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L. Roadkill-Data-Based Identification and Ranking of Mammal Habitats. Land 2021, 10, 477. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Stratford, J.; Balčiauskienė, L.; Kučas, A. Roadkills as a Method to Monitor Raccoon Dog Populations. Animals 2021, 11, 3147. [Google Scholar] [CrossRef]
- Colino-Rabanal, V.J.; Langen, T.A.; Peris, S.J.; Lizana, M. Ungulate: Vehicle Collision Rates Are Associated with the Phase of the Moon. Biodivers. Conserv. 2018, 27, 681–694. [Google Scholar] [CrossRef]
- Kučas, A.; Balčiauskas, L. Temporal Patterns of Ungulate-Vehicle Collisions in Lithuania. J. Environ. Manag. 2020, 273, 111172. [Google Scholar] [CrossRef]
- VĮ Lietuvos Automobilių Kelių Direkcija. Lietuvos Keliai. Available online: https://lakd.lrv.lt/lt/veiklos-sritys/lietuvos-keliai (accessed on 16 March 2021).
- VĮ Lietuvos Automobilių Kelių Direkcija. Eismo Apskaita Valstybinės Reikšmės Keliuose 2020 m. Available online: https://lakd.lrv.lt/uploads/lakd/documents/files/Eismo_intensyvumas/eismo_apskaita_valstybines_reiksmes_keliuose_2020_m.pdf (accessed on 16 March 2021).
- Jakimavičius, M. Analysis and Assessment of Lithuanian Road Accidents by AHP Method. Balt. J. Road Bridge Eng. 2018, 13, 238–260. [Google Scholar] [CrossRef]
- Brzeziński, M.; Romanowski, J.; Żmihorski, M.; Karpowicz, K. Muskrat (Ondatra zibethicus) decline after the expansion of American mink (Neovison vison) in Poland. Eur. J. Wildl. Res. 2010, 56, 341–348. [Google Scholar] [CrossRef]
- Lietuvos Respublikos Aplinkos Ministerija. Sumedžioti Žvėrys ir Paukščiai. Available online: https://am.lrv.lt/lt/veiklos-sritys-1/gamtos-apsauga/medziokle/ (accessed on 15 March 2022).
- Balčiauskas, L.; Ulevičius, A. Semi-aquatic mammal environment correlates in South Lithuanian river valleys. Ekologija 1995, 2, 37–44. [Google Scholar]
- Ulevicius, A.; Balciauskas, L. Ecology of semi-aquatic mammals in some landscapes of Lithuania: Abundance, community structure and distribution. Ekologija 1995, 2, 77–83. [Google Scholar]
- Balčiauskas, L.; Balčiauskienė, L.; Nature Research Centre, Vilnius, Lithuania. Personal Communication, 2022.
- Balčiauskas, L.; Nature Research Centre, Vilnius, Lithuania. Personal Communication, 2022.
- European Environmental Agency. CORINE Land Cover—Copernicus Land Monitoring Service [WWW Document]. 2018. Available online: https://land.copernicus.eu/pan-european/corine-land-cover/clc2018?tab=metadata (accessed on 15 March 2020).
- Brown, L.D.; Cat, T.T.; DasGupta, A. Interval Estimation for a proportion. Stat. Sci. 2001, 16, 101–133. [Google Scholar] [CrossRef]
- Dean, A.G.; Sullivan, K.M.; Soe, M.M. OpenEpi: Open Source Epidemiologic Statistics for Public Health. Available online: http://OpenEpi.com (accessed on 19 January 2021).
- G-Test Calculator. Available online: https://elem.com/~btilly/effective-ab-testing/g-test-calculator.html (accessed on 16 February 2021).
- Philcox, C.K.; Grogan, A.L.; Macdonald, D.W. Patterns of otter Lutra lutra road mortality in Britain. J. Appl. Ecol. 1999, 36, 748–762. [Google Scholar] [CrossRef]
- Chanin, P. Otter road casualties. Hystrix Ital. J. Mammal. 2006, 17, 79–90. [Google Scholar]
- Jancke, S.; Giere, P. Patterns of otter Lutra lutra road mortality in a landscape abundant in lakes. Eur. J. Wildl. Res. 2011, 57, 373–381. [Google Scholar] [CrossRef]
- Hauer, S.; Ansorge, H.; Zinke, O. Mortality patterns of otters (Lutra lutra) from eastern Germany. J. Zool. 2002, 256, 361–368. [Google Scholar] [CrossRef]
- Poledník, L.; Poledníková, K.; Větrovcová, J.; Hlaváč, V.; Beran, V. Causes of deaths of Lutra lutra in the Czech Republic (Carnivora: Mustelidae). Lynx Series Nova 2011, 42, 145–157. [Google Scholar]
- Cserkész, T.; Ottlecz, B.; Cserkész-Nagy, Á.; Farkas, J. Interchange as the main factor determining wildlife–vehicle collision hotspots on the fenced highways: Spatial analysis and applications. Eur. J. Wildl. Res. 2013, 59, 587–597. [Google Scholar] [CrossRef]
- Madsen, A.B. Otter Lutra lutra mortality in relation to traffic, and experience with newly established fauna passages at existing road bridges. Lutra 1996, 39, 76–90. [Google Scholar]
- Guter, A.; Dolev, A.; Saltz, D.; Kronfeld-Schor, N. Temporal and spatial influences on road mortality in otters: Conservation implications. Isr. J. Ecol. Evol. 2005, 51, 199–207. [Google Scholar] [CrossRef]
- Rendall, A.R.; Webb, V.; Sutherland, D.R.; White, J.G.; Renwick, L.; Cooke, R. Where wildlife and traffic collide: Roadkill rates change through time in a wildlife-tourism hotspot. Glob. Ecol. Conserv. 2021, 27, e01530. [Google Scholar] [CrossRef]
- Pokorny, B.; Cerri, J.; Bužan, E. Wildlife roadkill and COVID-19: A biologically significant, but heterogeneous, reduction. J. Appl. Ecol. 2022, 59, 1291–1301. [Google Scholar] [CrossRef]
- Barthelmess, E.L.; Brooks, M.S. The influence of body-size and diet on road-kill trends in mammals. Biodivers. Conserv. 2010, 19, 1611–1629. [Google Scholar] [CrossRef]
- Kustusch, K.; Wuczynski, A.; Smiertelnosc Zwierzat na Drogach w Polsce w Roku 2016. Raport Roczny “Ogólnopolskiego Rejestru Smiertelnosci Zwierzat na Drogach”. Available online: https://zwierzetanadrodze.pl/files/Zwierzeta_na_Drodze_-_Raport_2016.pdf (accessed on 3 November 2021).
- Orlowski, G.; Nowak, L. Factors influencing mammal roadkills in the agricultural landscape of south-western Poland. Pol. J. Ecol. 2006, 54, 283–294. [Google Scholar]
- Gryz, J.; Krauze, D. Mortality of vertebrates on a road crossing the Biebrza Valley (NE Poland). Eur. J. Wildl. Res. 2008, 54, 709–714. [Google Scholar] [CrossRef]
- Pintur, K.; Slijepčević, V.; Popović, N.; Andrijašević, D. Dynamics of wildlife-vehicle collisions on roads of karlovac county, Croatia. J. Cent. Eur. Agric. 2012, 13, 340–349. [Google Scholar] [CrossRef] [Green Version]
- Šprem, N.; Duduković, D.; Keros, T.; Konjević, D. Wildlife-vehicle collisions in Croatia—A hazard for humans and animals. Coll. Antropol. 2013, 37, 531–535. [Google Scholar] [PubMed]
- Guinard, E.; Billon, C.; Bretaud, F.; Sordello, R.; Chevallier, L.; Witté, I. Comparative study of animal carcass survey methods on roads. Transport. Res. D 2022. submitted. [Google Scholar] [CrossRef]
- Morelle, K.; Lehaire, F.; Lejeune, P. Spatio-temporal patterns of wildlife-vehicle collisions in a region with a high-density road network. Nat. Conserv. 2013, 5, 53–73. [Google Scholar] [CrossRef] [Green Version]
- Haigh, A. Annual patterns of mammalian mortality on Irish roads. Hystrix Ital. J. Mammal. 2012, 23, 57–65. [Google Scholar] [CrossRef]
- Barthelmess, E.L. Spatial distribution of road-kills and factors influencing road mortality for mammals in Northern New York State. Biodivers. Conserv. 2014, 23, 2491–2514. [Google Scholar] [CrossRef]
- Winton, B.R.; Takekawa, J.Y. Transportation Impacts to Wildlife on State Route 37 in Northern San Pablo Bay, California. Trans. West. Sect. Wildl. Soc. 2001, 37, 55–60. [Google Scholar]
- Körbel, O. Hindering Otter Lutra lutra Road Kills Part 2. IUCN Otter Spec. Group Bull. 1995, 11, 32–36. [Google Scholar]
- Quiles Tundidor, P.; Ascensão, F.; D’Amico, M.; Revilla, E.; Barrientos, R. Are road-kills representative of wildlife community obtained from atlas data? Hystrix Ital. J. Mammal. 2021, 32, 89–94. [Google Scholar] [CrossRef]
- Brieger, F.; Kämmerle, J.L.; Hagen, R.; Suchant, R. Behavioural reactions to oncoming vehicles as a crucial aspect of wildlife-vehicle collision risk in three common wildlife species. Accid. Anal. Prev. 2022, 168, 106564. [Google Scholar] [CrossRef] [PubMed]
- Barela, I.; Burger, L.M.; Taylor, J.; Evans, K.O.; Ogawa, R.; McClintic, L.; Wang, G. Relationships between survival and habitat suitability of semi-aquatic mammals. Ecol. Evol. 2020, 10, 4867–4875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zschille, J.; Stier, N.; Roth, M.; Berger, U. Dynamics in space use of American mink (Neovison vison) in a fishpond area in Northern Germany. Eur. J. Wildl. Res. 2012, 58, 955–968. [Google Scholar] [CrossRef]
- Galanaki, A.; Kominos, T. The distribution of American mink (Neovison vison) in Greece. Mammalia 2022, 86, 57–65. [Google Scholar] [CrossRef]
- Mori, E.; Bini, A.; Viviano, A.; Bartolommei, P.; Mazza, G. Distribution of introduced American mink in the Northern Apennine area (Central Italy). Mammalia 2022. [Google Scholar] [CrossRef]
- Brzeziński, M.; Ignatiuk, P.; Żmihorski, M.; Zalewski, A. An invasive predator affects habitat use by native prey: American mink and water vole co-existence in riparian habitats. J. Zool. 2018, 304, 109–116. [Google Scholar] [CrossRef]
- Ahlers, A.A.; Cotner, L.A.; Wolff, P.J.; Mitchell, M.A.; Heske, E.J.; Schooley, R.L. Summer precipitation predicts spatial distributions of semiaquatic mammals. PLoS ONE 2015, 10, e0135036. [Google Scholar] [CrossRef] [Green Version]
- Ward, E.M.; Wysong, K.; Gorelick, S.M. Drying landscape and interannual herbivory-driven habitat degradation control semiaquatic mammal population dynamics. Ecohydrology 2020, 13, e2169. [Google Scholar] [CrossRef]
- Holland, A.M.; Schauber, E.M.; Nielsen, C.K.; Hellgren, E.C. River otter and mink occupancy dynamics in riparian systems. J. Wildl. Manag. 2019, 83, 1552–1564. [Google Scholar] [CrossRef] [Green Version]
- Skyriene, G.; Paulauskas, A. Distribution of invasive muskrats (Ondatra zibethicus) and impact on ecosystem. Ekologija 2012, 58, 357–367. [Google Scholar] [CrossRef] [Green Version]
- Skyrienė, G.; Paulauskas, A.; Ulevičius, A.; Samas, A. Decline of invasive muskrat in Lithuania. Весmник Oxomoведения 2014, 11, 291–294. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Beaver (C. fiber) | American Mink (N. vison) | Muskrat (O. zibethicus) | |||
|---|---|---|---|---|---|---|
| Population | Hunting Bag | Population | Hunting Bag | Population | Hunting Bag | |
| 2003 | 11,012 | 5630 | n/a | 66 | n/a | 75 |
| 2004 | 20,926 | 5890 | n/a | 92 | n/a | 31 |
| 2005 | n/a | 4713 | n/a | 87 | n/a | 21 |
| 2006 | 23,778 | 5359 | n/a | 95 | n/a | 105 |
| 2007 | 31,160 | 12,473 | n/a | 110 | n/a | 79 |
| 2008 | 36,375 | 12,702 | n/a | 109 | n/a | 60 |
| 2009 | 40,222 | 13,593 | n/a | 95 | n/a | 72 |
| 2010 | 47,702 | 16,231 | n/a | 107 | n/a | 30 |
| 2011 | 48,046 | 18,821 | n/a | 82 | n/a | 72 |
| 2012 | 48,604 | 20,591 | n/a | 148 | n/a | 231 |
| 2013 | 49,446 | 11,778 | n/a | 153 | n/a | 148 |
| 2014 | 44,416 | 21,749 | n/a | 281 | n/a | 329 |
| 2015 | 43,802 | 19,544 | n/a | 166 | n/a | 99 |
| 2016 | 40,618 | 19,293 | n/a | 97 | n/a | 76 |
| 2017 | 40,506 | 17,503 | n/a | 81 | n/a | 73 |
| 2018 | 42,396 | 15,637 | n/a | 59 | n/a | 60 |
| 2019 | 43,148 | 19,977 | n/a | 106 | n/a | 42 |
| 2020 | 43,551 | 19,907 | n/a | 46 | n/a | 39 |
| 2021 | 43,355 | 15,651 | n/a | 53 | n/a | 23 |
| Species | N |
|---|---|
| Roe deer (Capreolus capreolus) | 25,583 |
| Moose (Alces alces) | 2457 |
| Wild boar (Sus scrofa) | 2027 |
| Raccoon dog (Nyctereutes procyonoides) nn | 1659 |
| Red fox (Vulpes vulpes) | 1175 |
| Eastern European hedgehog (Erinaceus roumanicus) | 1158 |
| European hare (Lepus europaeus) | 739 |
| Red deer (Cervus elaphus) | 633 |
| Marten (Martes sp.) | 413 |
| Badger (Meles meles) | 203 |
| European polecat (Mustela putorius) | 184 |
| Pine marten (Martes martes) | 70 |
| Beaver (Castor fiber) | 60 |
| Stone marten (Martes foina) | 35 |
| Red squirrel (Sciurus vulgaris) | 40 |
| Fallow deer (Dama dama) nn | 45 |
| American mink (Neovison vison)nn | 26 |
| Grey wolf (Canis lupus) | 25 |
| Eurasian otter (Lutra lutra) | 22 |
| European mole (Talpa europaea) | 20 |
| Muskrat (Ondatra zibethicus)nn | 3 |
| Unknown | 3313 |
| Other species | 46 |
| Total, N | 39,936 |
| C. fiber | L. lutra | N. vison | O. zibethicus | |
|---|---|---|---|---|
| CLC water | 2823 ± 371 0–11,310 | 2563 ± 619 159–10,450 | 2343 ± 389 338–9704 | 2376 ± 1085 229–3719 |
| CLC wetland | 7067 ± 782 296–21,824 | 6503 ± 1173 267–20,608 | 5434 ± 1056 719–27,233 | 7372 ± 4695 159–16,185 |
| GPRK river | 303 ± 101 6–4256 | 284 ± 63 13–1059 | 324 ± 64 13–1388 | 90 ± 10 73–106 |
| GPRK lake | 1093 ± 156 13–4116 | 1260 ± 203 179–2961 | 1108 ± 193 114–4100 | 901 ± 547 229–1984 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiauskas, L.; Stratford, J.; Kučas, A.; Balčiauskienė, L. Factors Affecting Roadkills of Semi-Aquatic Mammals. Biology 2022, 11, 748. https://doi.org/10.3390/biology11050748
Balčiauskas L, Stratford J, Kučas A, Balčiauskienė L. Factors Affecting Roadkills of Semi-Aquatic Mammals. Biology. 2022; 11(5):748. https://doi.org/10.3390/biology11050748
Chicago/Turabian StyleBalčiauskas, Linas, Jos Stratford, Andrius Kučas, and Laima Balčiauskienė. 2022. "Factors Affecting Roadkills of Semi-Aquatic Mammals" Biology 11, no. 5: 748. https://doi.org/10.3390/biology11050748
APA StyleBalčiauskas, L., Stratford, J., Kučas, A., & Balčiauskienė, L. (2022). Factors Affecting Roadkills of Semi-Aquatic Mammals. Biology, 11(5), 748. https://doi.org/10.3390/biology11050748