
Receptor Activity Modifying Protein RAMP Sub-Isoforms and Their Functional Differentiation, Which Regulates Functional Diversity of Adrenomedullin

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
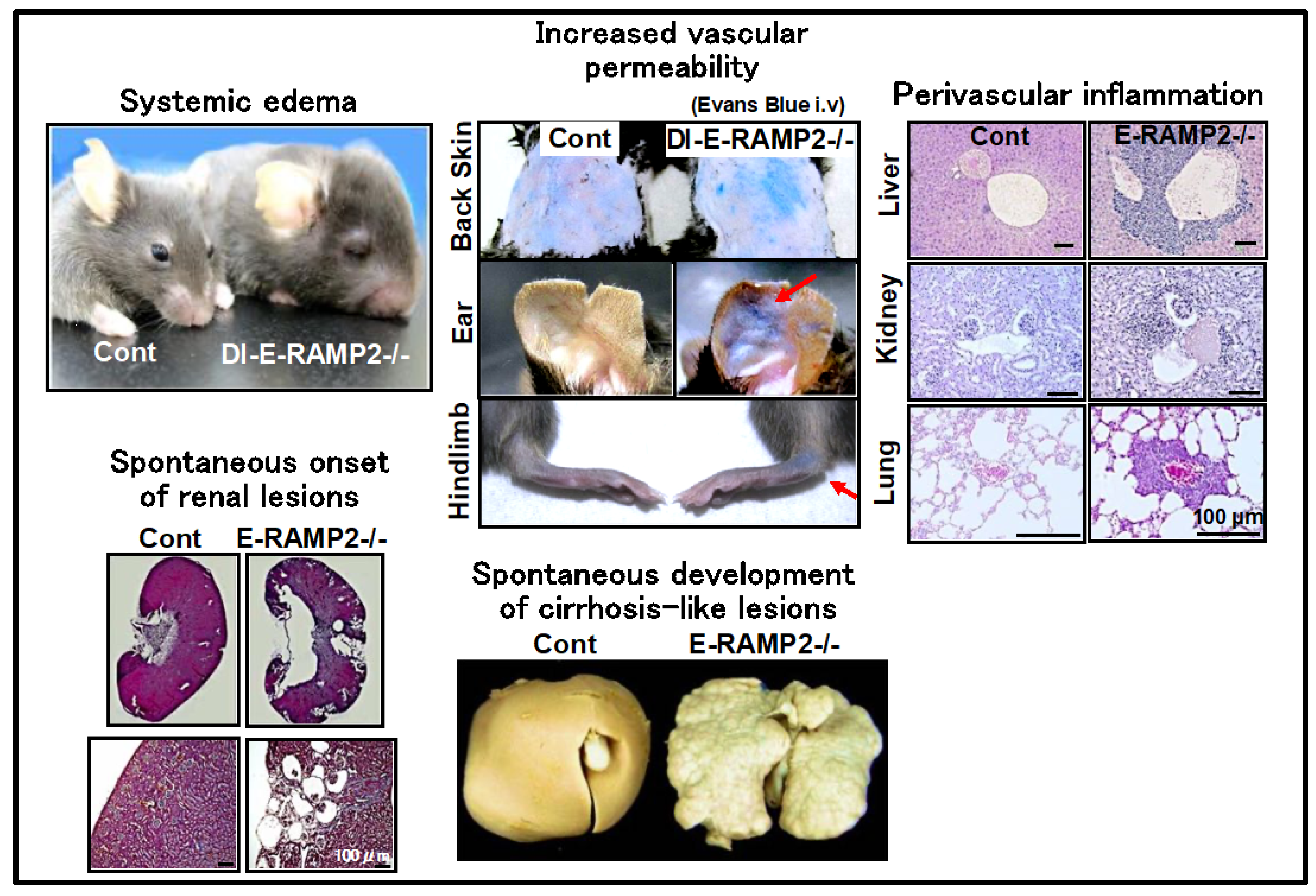
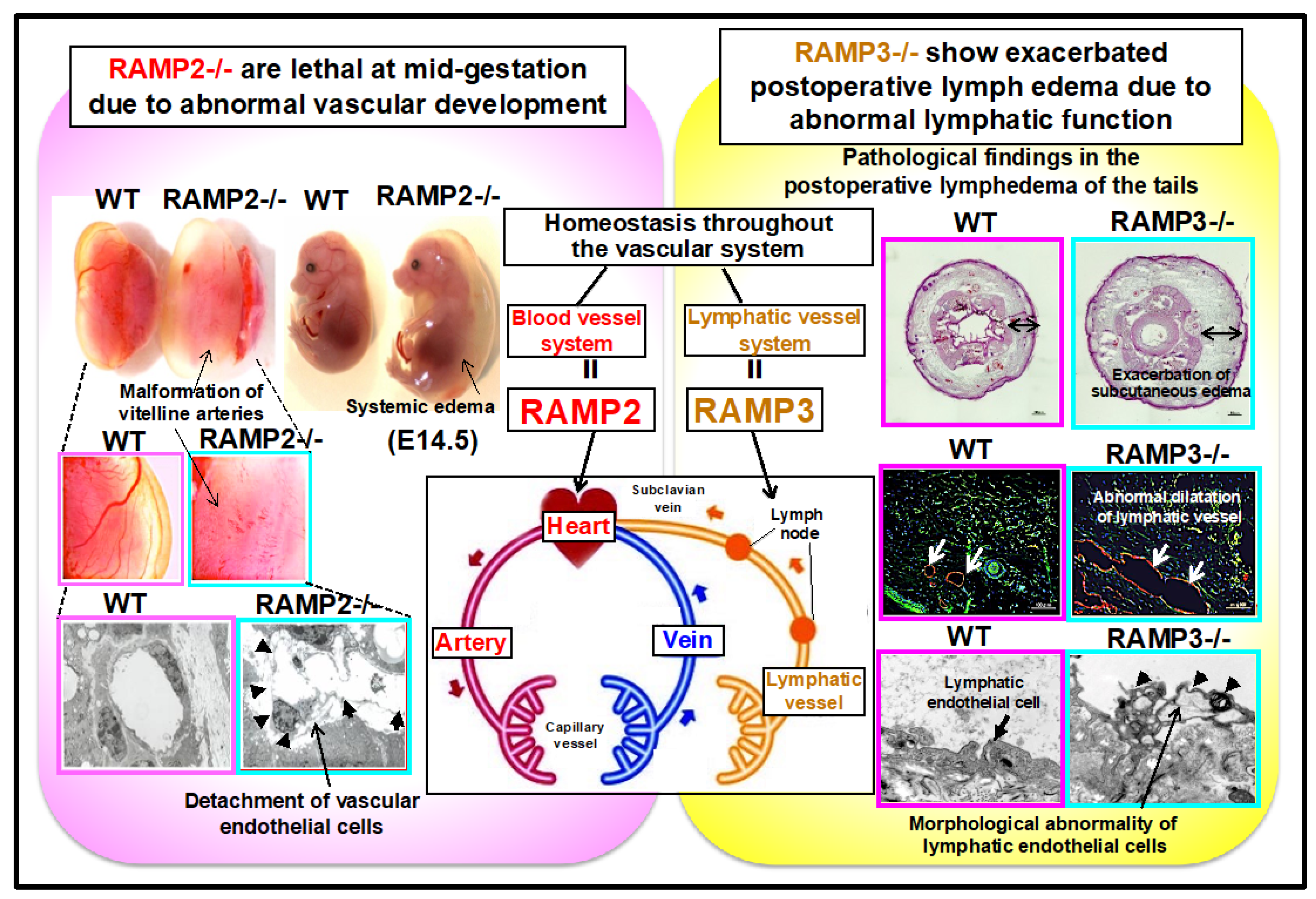
2.1. Regulation of Vascular Homeostasis by the AM-RAMP2 System
2.2. Functional Differentiation of AM-RAMP2 and AM-RAMP3 Systems in the Regulation of Vascular Homeostasis
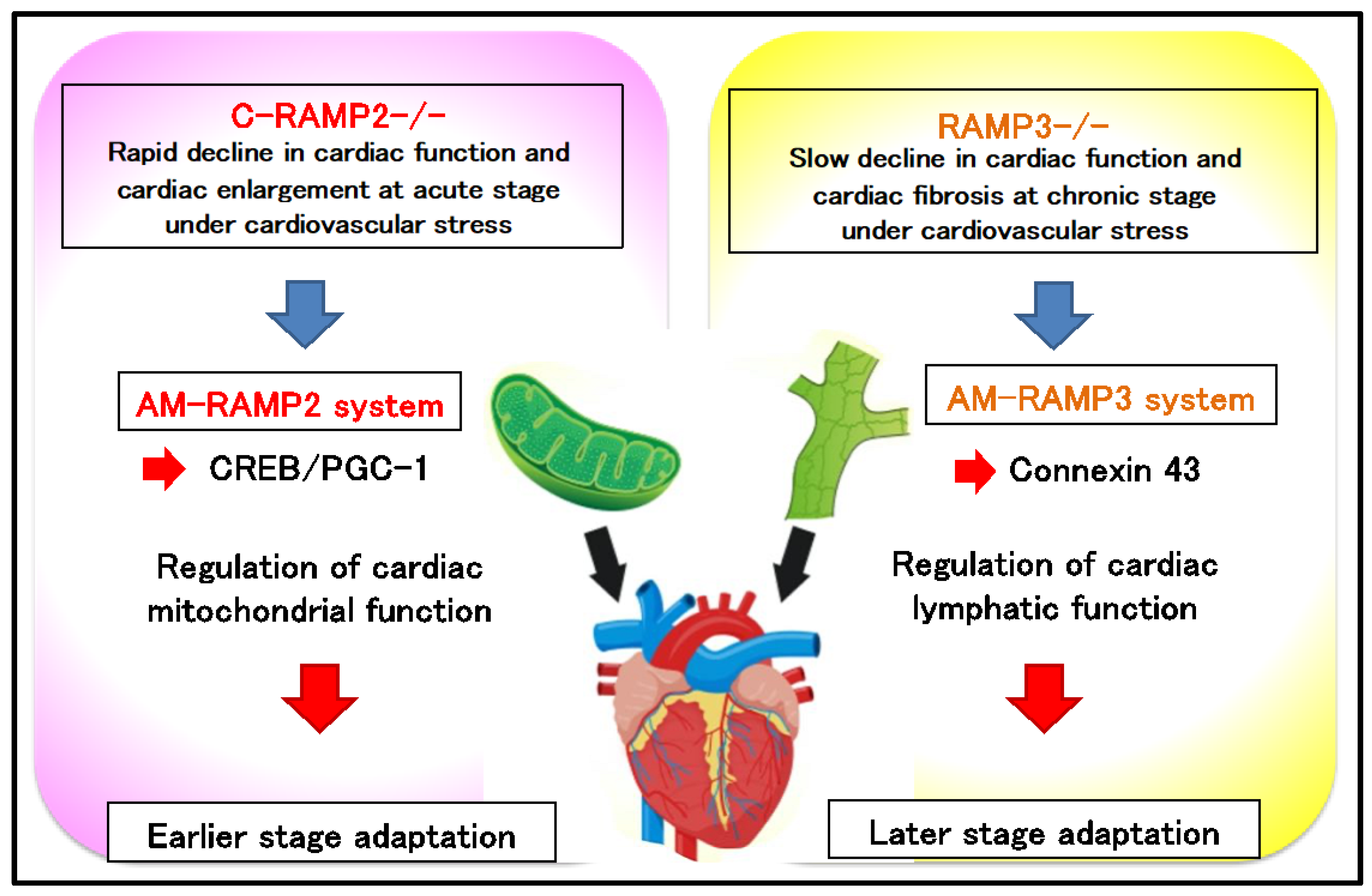
2.3. Functional Differentiation of AM-RAMP2 and AM-RAMP3 Systems in Cardiovascular Stress Response
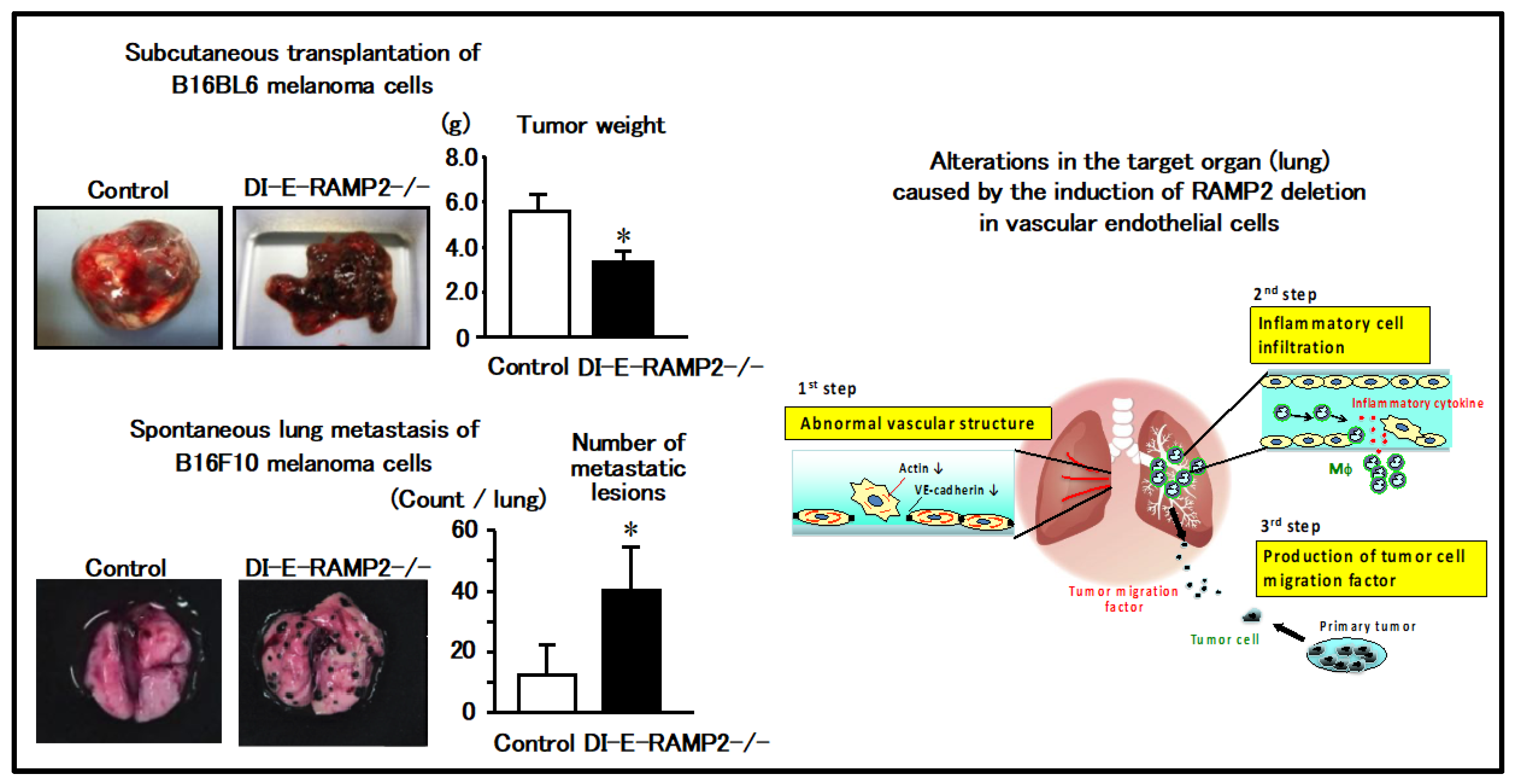
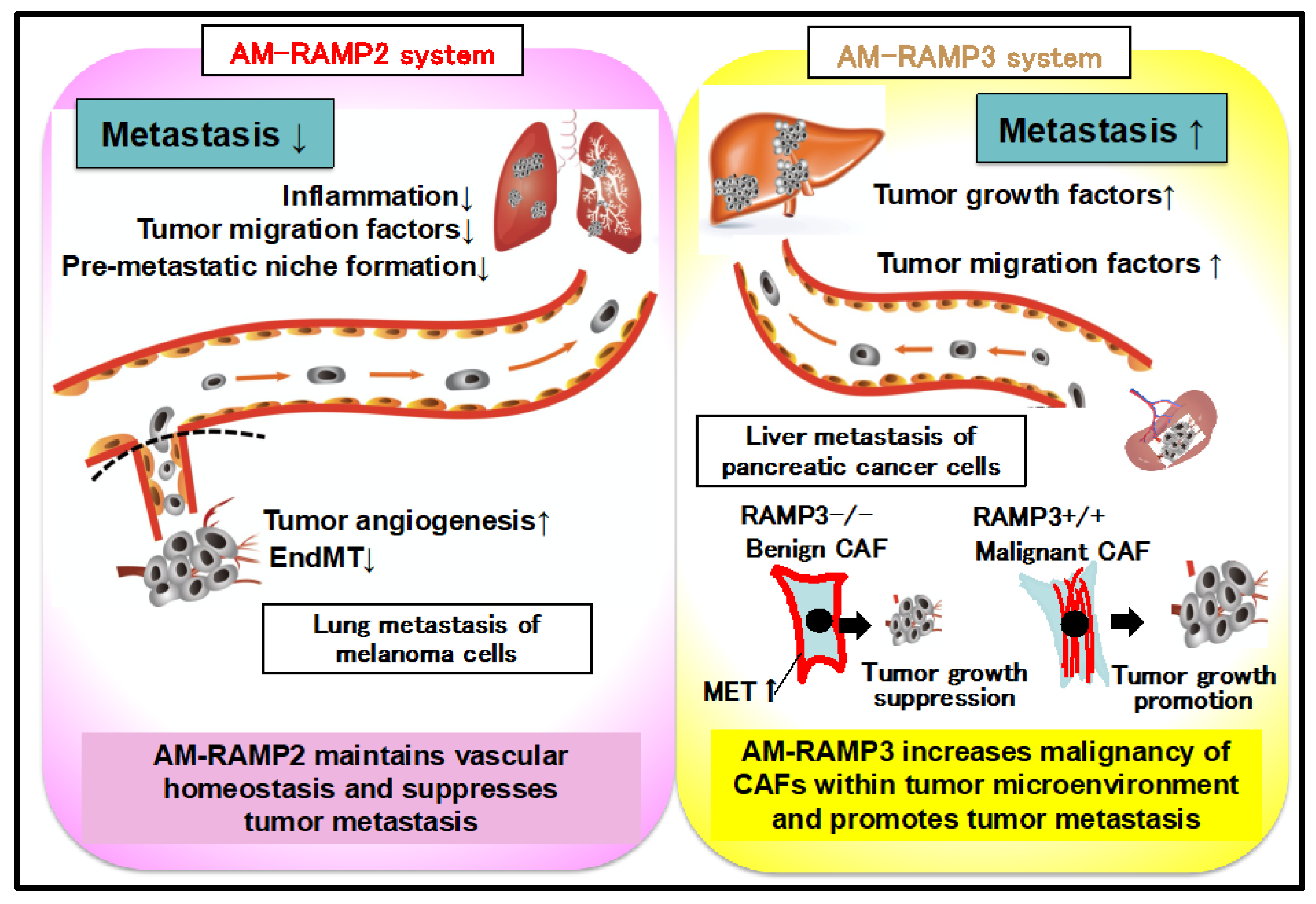
2.4. Regulation of Vascular Homeostasis and Suppression of Cancer Metastasis by the AM-RAMP2 System
2.5. Antagonism between AM-RAMP2 and AM-RAMP3 Systems in Cancer Metastasis
3. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kitamura, K.; Kangawa, K.; Kawamoto, M.; Ichiki, Y.; Nakamura, S.; Matsuo, H.; Eto, T. Adrenomedullin: A Novel Hypotensive Peptide Isolated from Human Pheochromocytoma. Biochem. Biophys. Res. Commun. 1993, 192, 553–560. [Google Scholar] [CrossRef] [PubMed]
- Shindo, T.; Tanaka, M.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Yamauchi, A.; Sakurai, T. Regulation of Cardiovascular Development and Homeostasis by the Adrenomedullin-RAMP System. Peptides 2019, 111, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Ashizuka, S.; Inatsu, H.; Inagaki-Ohara, K.; Kita, T.; Kitamura, K. Adrenomedullin as a Potential Therapeutic Agent for Inflammatory Bowel Disease. Curr. Protein Pept. Sci. 2013, 14, 246–255. [Google Scholar] [CrossRef] [PubMed]
- Ashizuka, S.; Inatsu, H.; Kita, T.; Kitamura, K. Adrenomedullin Therapy in Patients with Refractory Ulcerative Colitis: A Case Series. Dig. Dis. Sci. 2016, 61, 872–880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLatchie, L.M.; Fraser, N.J.; Main, M.J.; Wise, A.; Brown, J.; Thompson, N.; Solari, R.; Lee, M.G.; Foord, S.M. RAMPs Regulate the Transport and Ligand Specificity of the Calcitonin-Receptor-Like Receptor. Nature 1998, 393, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Shindo, T.; Kurihara, Y.; Nishimatsu, H.; Moriyama, N.; Kakoki, M.; Wang, Y.; Imai, Y.; Ebihara, A.; Kuwaki, T.; Ju, K.H.; et al. Vascular Abnormalities and Elevated Blood Pressure in Mice Lacking Adrenomedullin Gene. Circulation 2001, 104, 1964–1971. [Google Scholar] [CrossRef] [Green Version]
- Ichikawa-Shindo, Y.; Sakurai, T.; Kamiyoshi, A.; Kawate, H.; Iinuma, N.; Yoshizawa, T.; Koyama, T.; Fukuchi, J.; Iimuro, S.; Moriyama, N.; et al. The GPCR Modulator Protein RAMP2 Is Essential for Angiogenesis and Vascular Integrity. J. Clin. Investig. 2008, 118, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, T.; Ochoa-Callejero, L.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Iinuma, N.; Arai, T.; Yoshizawa, T.; Iesato, Y.; Lei, Y.; et al. Vascular Endothelial Adrenomedullin-RAMP2 System Is Essential for Vascular Integrity and Organ Homeostasis. Circulation 2013, 127, 842–853. [Google Scholar] [CrossRef]
- Prasain, N.; Stevens, T. The Actin Cytoskeleton in Endothelial Cell Phenotypes. Microvasc. Res. 2009, 77, 53–63. [Google Scholar] [CrossRef] [Green Version]
- Xian, X.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Tanaka, M.; Koyama, T.; Kawate, H.; Yang, L.; Liu, T.; Imai, A.; et al. Vasoprotective Activities of the Adrenomedullin-RAMP2 System in Endothelial Cells. Endocrinology 2017, 158, 1359–1372. [Google Scholar] [CrossRef] [Green Version]
- Yamauchi, A.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Igarashi, K.; Toriyama, Y.; Tanaka, M.; Liu, T.; Xian, X.; et al. Functional Differentiation of RAMP2 and RAMP3 in Their Regulation of the Vascular System. J. Mol. Cell. Cardiol. 2014, 77, 73–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizawa, T.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Iesato, Y.; Koyama, T.; Uetake, R.; Yang, L.; Yamauchi, A.; et al. Novel Regulation of Cardiac Metabolism and Homeostasis by the Adrenomedullin-Receptor Activity-Modifying Protein 2 System. Hypertension 2013, 61, 341–351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, N.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Tanaka, M.; Tanaka, M.; Wei, Y.; Kakihara, S.; Zhao, Y.; et al. Adrenomedullin-RAMP2 and -RAMP3 Systems Regulate Cardiac Homeostasis During Cardiovascular Stress. Endocrinology 2021, 162, bqab001. [Google Scholar] [CrossRef] [PubMed]
- Herzig, S.; Long, F.; Jhala, U.S.; Hedrick, S.; Quinn, R.; Bauer, A.; Rudolph, D.; Schutz, G.; Yoon, C.; Puigserver, P.; et al. CREB Regulates Hepatic Gluconeogenesis Through the coactivator PGC-1. Nature 2001, 413, 179–183. [Google Scholar] [CrossRef]
- Karpinich, N.O.; Caron, K.M. Gap Junction Coupling Is Required for Tumor Cell Migration Through Lymphatic Endothelium. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 1147–1155. [Google Scholar] [CrossRef] [Green Version]
- Trincot, C.E.; Xu, W.; Zhang, H.; Kulikauskas, M.R.; Caranasos, T.G.; Jensen, B.C.; Sabine, A.; Petrova, T.V.; Caron, K.M. Adrenomedullin Induces Cardiac Lymphangiogenesis After Myocardial Infarction and Regulates Cardiac Edema via Connexin 43. Circ. Res. 2019, 124, 101–113. [Google Scholar] [CrossRef]
- Tsuruda, T.; Kato, J.; Kuwasako, K.; Kitamura, K. Adrenomedullin: Continuing to Explore Cardioprotection. Peptides 2019, 111, 47–54. [Google Scholar] [CrossRef]
- Miller, M.J.; Martínez, A.; Unsworth, E.J.; Thiele, C.J.; Moody, T.W.; Elsasser, T.; Cuttitta, F. Adrenomedullin Expression in Human Tumor Cell Lines. Its Potential Role as an Autocrine Growth Factor. J. Biol. Chem. 1996, 271, 23345–23351. [Google Scholar] [CrossRef] [Green Version]
- Siclari, V.A.; Mohammad, K.S.; Tompkins, D.R.; Davis, H.; McKenna, C.R.; Peng, X.; Wessner, L.L.; Niewolna, M.; Guise, T.A.; Suvannasankha, A.; et al. Tumor-Expressed Adrenomedullin Accelerates Breast Cancer Bone Metastasis. Breast Cancer Res. 2014, 16, 458. [Google Scholar] [CrossRef] [Green Version]
- Ramachandran, V.; Arumugam, T.; Hwang, R.F.; Greenson, J.K.; Simeone, D.M.; Logsdon, C.D. Adrenomedullin Is Expressed in Pancreatic Cancer and Stimulates Cell Proliferation and Invasion in an Autocrine Manner via the Adrenomedullin Receptor, ADMR. Cancer Res. 2007, 67, 2666–2675. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Koyama, T.; Sakurai, T.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Liu, T.; Xian, X.; Imai, A.; Zhai, L.; et al. The Endothelial Adrenomedullin-RAMP2 System Regulates Vascular Integrity and Suppresses Tumour Metastasis. Cardiovasc. Res. 2016, 111, 398–409. [Google Scholar] [CrossRef] [PubMed]
- Lin, F.; Wang, N.; Zhang, T.C. The Role of Endothelial-Mesenchymal Transition in Development and Pathological Process. IUBMB Life 2012, 64, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, S.; Watanabe, A.; Aburatani, H.; Maru, Y. Tumour-Mediated Upregulation of Chemoattractants and Recruitment of Myeloid Cells Predetermines Lung Metastasis. Nat. Cell Biol. 2006, 8, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, S.; Watanabe, A.; Sakurai, Y.; Akashi-Takamura, S.; Ishibashi, S.; Miyake, K.; Shibuya, M.; Akira, S.; Aburatani, H.; Maru, Y. The S100A8-Serum Amyloid A3-TLR4 Paracrine Cascade Establishes a Pre-Metastatic Phase. Nat. Cell Biol. 2008, 10, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, S.; Ishibashi, S.; Tomita, T.; Watanabe, A.; Akashi-Takamura, S.; Murakami, M.; Kijima, H.; Miyake, K.; Aburatani, H.; Maru, Y. Primary Tumours Modulate Innate Immune Signalling to Create Pre-Metastatic Vascular Hyperpermeability Foci. Nat. Commun. 2013, 4, 1853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, K.; Tanaka, M.; Kamiyoshi, A.; Sakurai, T.; Ichikawa-Shindo, Y.; Kawate, H.; Cui, N.; Wei, Y.; Tanaka, M.; Kakihara, S.; et al. Deficiency of the Adrenomedullin-RAMP3 System Suppresses Metastasis Through the Modification of Cancer-Associated Fibroblasts. Oncogene 2020, 39, 1914–1930. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Aishima, S.; Ohuchida, K.; Fujiwara, K.; Fujino, M.; Mizuuchi, Y.; Hattori, M.; Mizumoto, K.; Tanaka, M.; Oda, Y. Podoplanin Expression in Cancer-associated Fibroblasts Enhances Tumor Progression of Invasive Ductal Carcinoma of the Pancreas. Mol. Cancer. 2013, 12, 168. [Google Scholar] [CrossRef] [Green Version]
- Inoue, H.; Miyazaki, Y.; Kikuchi, K.; Yoshida, N.; Ide, F.; Ohmori, Y.; Tomomura, A.; Sakashita, H.; Kusama, K. Podoplanin Promotes Cell Migration via the EGF-Src-Cas Pathway in Oral Squamous Cell Carcinoma Cell Lines. J. Oral Sci. 2012, 54, 241–250. [Google Scholar] [CrossRef] [Green Version]
- Edvinsson, L.; Warfvinge, K. Recognizing the Role of CGRP and CGRP Receptors in Migraine and Its Treatment. Cephalalgia 2019, 39, 366–373. [Google Scholar] [CrossRef]
- Ochoa-Callejero, L.; García-Sanmartín, J.; Martínez-Herrero, S.; Rubio-Mediavilla, S.; Narro-Íñiguez, J.; Martínez, A. Small Molecules Related to Adrenomedullin Reduce Tumor Burden in a Mouse Model of Colitis-associated Colon Cancer. Sci. Rep. 2017, 7, 17488. [Google Scholar] [CrossRef] [Green Version]
- Blet, A.; Deniau, B.; Geven, C.; Sadoune, M.; Caillard, A.; Kounde, P.R.; Polidano, E.; Pickkers, P.; Samuel, J.L.; Mebazaa, A. Adrecizumab, a Non-neutralizing Anti-adrenomedullin Antibody, Improves Haemodynamics and Attenuates Myocardial Oxidative Stress in Septic Rats. Intensive Care Med. Exp. 2019, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Kusano, S.; Kukimoto-Niino, M.; Akasaka, R.; Toyama, M.; Terada, T.; Shirouzu, M.; Shindo, T.; Yokoyama, S. Crystal Structure of the Human Receptor Activity-Modifying Protein 1 Extracellular Domain. Protein Sci. 2008, 17, 1907–1914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kusano, S.; Kukimoto-Niino, M.; Hino, N.; Ohsawa, N.; Okuda, K.; Sakamoto, K.; Shirouzu, M.; Shindo, T.; Yokoyama, S. Structural Basis for Extracellular Interactions Between Calcitonin Receptor-Like Receptor and Receptor Activity-Modifying Protein 2 for Adrenomedullin-Specific Binding. Protein Sci. 2012, 21, 199–210. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RAMP2 Knockout Mice | RAMP3 Knockout Mice |
|---|---|
| Conventional RAMP2-/- ref. [7] | Conventional RAMP3-/- ref. [11] |
| Embryonic lethal at midgestation | No abnormalities in vascular development |
| Edema and bleeding | Abnormal lymphatic function in adult |
| Abnormality of endothelial cells | Enhanced postoperative lymph edema |
| Endothelial cell-specific RAMP2-/- ref. [8] | |
| Lethal around perinatal period (95%) | |
| Systemic edema | |
| Survive until adulthood (5%) | |
| Vasculitis within major organs | |
| Organ damages | |
| Drug-inducible endothelial cell-specific RAMP2-/- | |
| Enhanced cancer cell metastasis | Reduced cancer cell metastasis ref. [26] |
| Cardiac myocyte cell-specific RAMP2-/- ref. [13] | Slow decline in cardiac function under cardiovascular stress ref. [13] |
| Rapid decline in cardiac function under cardiovascular stress | Reduced cardiac lymphatic vessels |
| Mitochondrial dysfunction | Reduced expression of connexin 43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shindo, T.; Tanaka, M.; Kamiyoshi, A.; Ichikawa-Shindo, Y.; Kawate, H.; Sakurai, T. Receptor Activity Modifying Protein RAMP Sub-Isoforms and Their Functional Differentiation, Which Regulates Functional Diversity of Adrenomedullin. Biology 2022, 11, 788. https://doi.org/10.3390/biology11050788
Shindo T, Tanaka M, Kamiyoshi A, Ichikawa-Shindo Y, Kawate H, Sakurai T. Receptor Activity Modifying Protein RAMP Sub-Isoforms and Their Functional Differentiation, Which Regulates Functional Diversity of Adrenomedullin. Biology. 2022; 11(5):788. https://doi.org/10.3390/biology11050788
Chicago/Turabian StyleShindo, Takayuki, Megumu Tanaka, Akiko Kamiyoshi, Yuka Ichikawa-Shindo, Hisaka Kawate, and Takayuki Sakurai. 2022. "Receptor Activity Modifying Protein RAMP Sub-Isoforms and Their Functional Differentiation, Which Regulates Functional Diversity of Adrenomedullin" Biology 11, no. 5: 788. https://doi.org/10.3390/biology11050788
APA StyleShindo, T., Tanaka, M., Kamiyoshi, A., Ichikawa-Shindo, Y., Kawate, H., & Sakurai, T. (2022). Receptor Activity Modifying Protein RAMP Sub-Isoforms and Their Functional Differentiation, Which Regulates Functional Diversity of Adrenomedullin. Biology, 11(5), 788. https://doi.org/10.3390/biology11050788