
N6-Methyladenosine Methylome Profiling of Muscle and Adipose Tissues Reveals Methylase–mRNA Metabolic Regulatory Networks in Fat Deposition of Rex Rabbits

Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Animals and Tissue Collection
2.2. RNA Extraction and Fragmentation
2.3. M6A Immunoprecipitation and Library Construction
2.4. RNA Extraction and cDNA Synthesis
2.5. Primer Design and Quantitative Real-Time PCR
2.6. KEGG and Gene Screening
2.7. Quality Control, Mapping and Statistical Analysis
3. Results
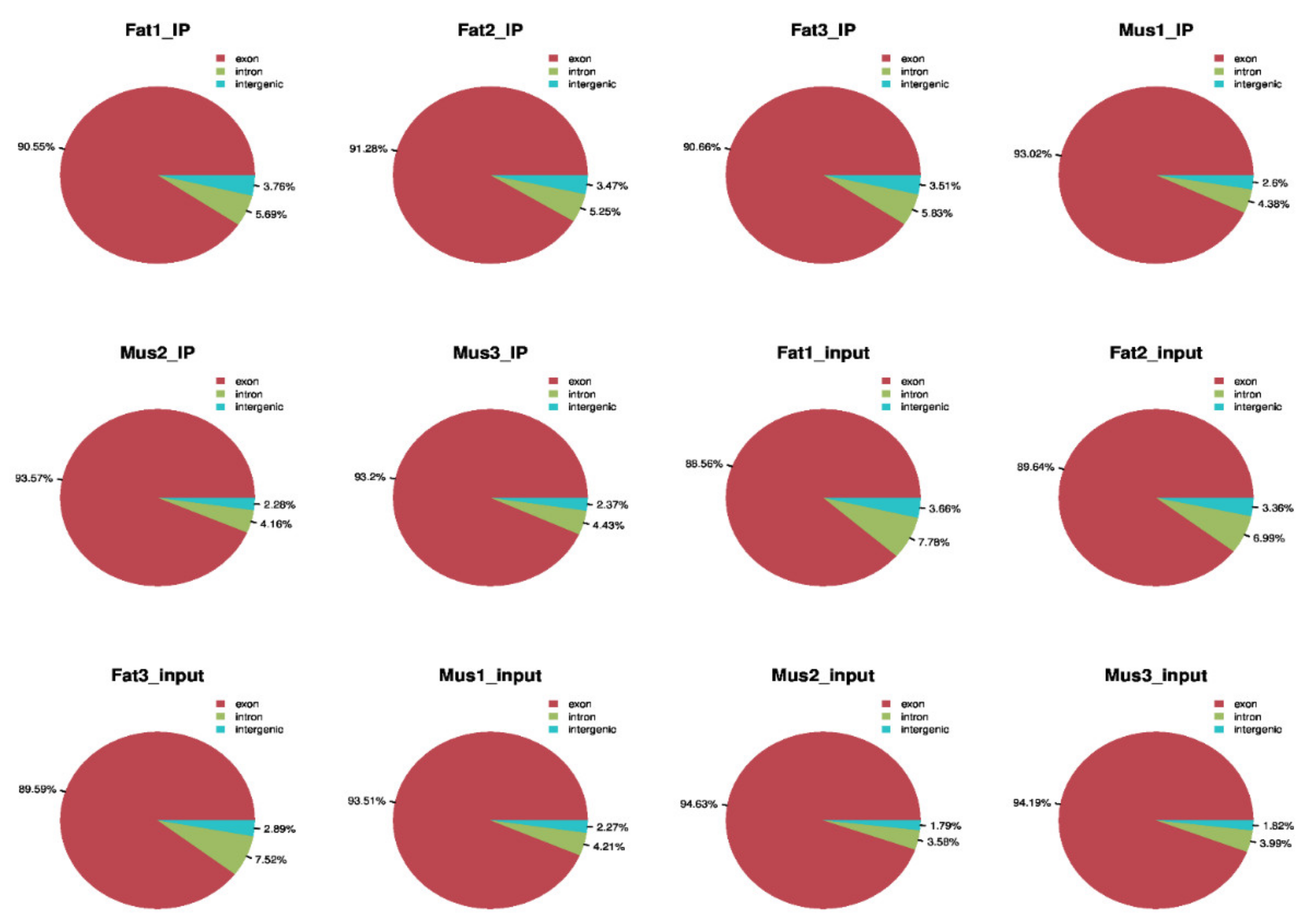
3.1. Sequencing Statistics and Quality Control
3.2. Mapping Reads to the Reference Genome
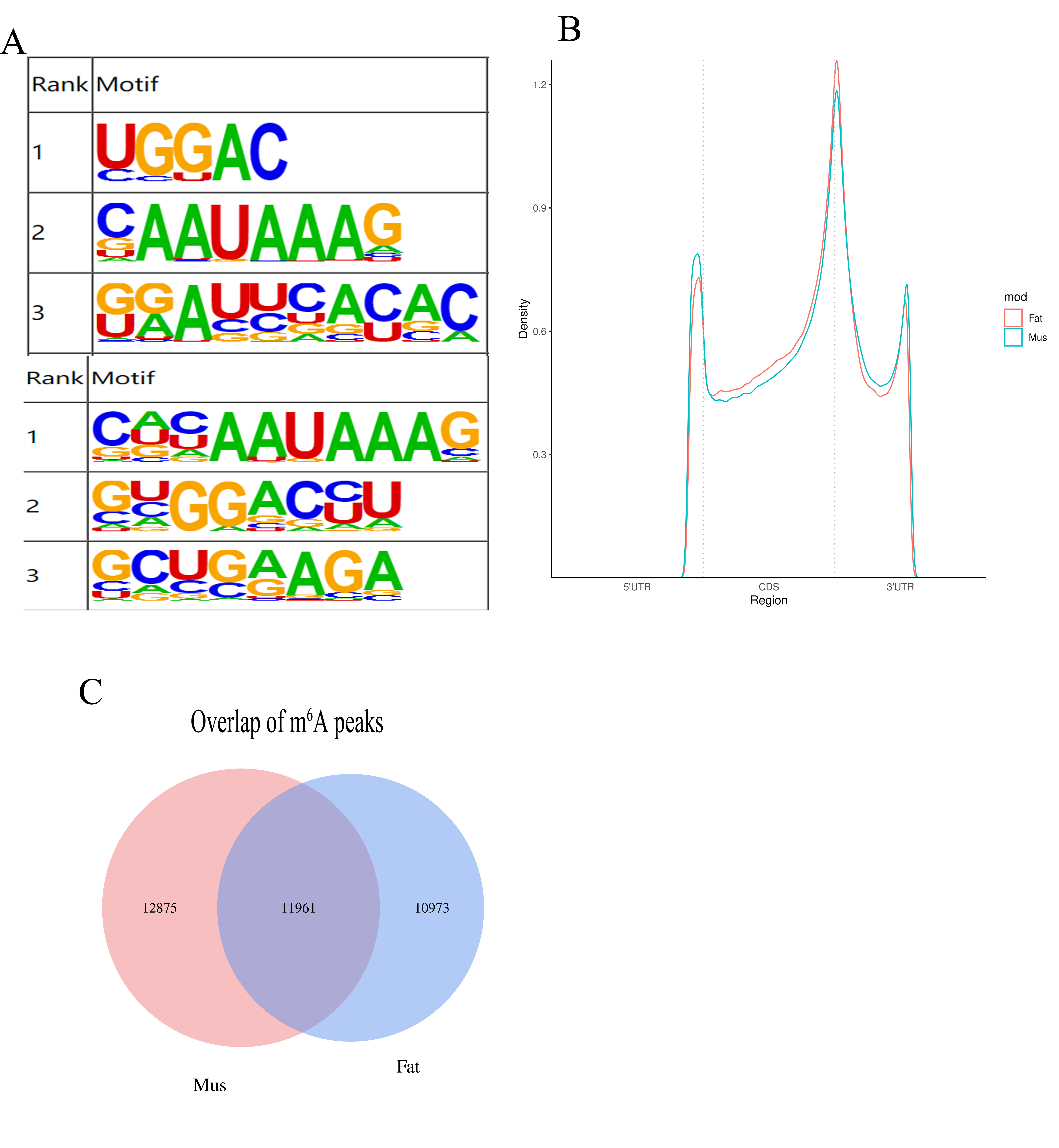
3.3. Transcriptome-Wide Detection and Distribution of m6A Modification in Rex Rabbits
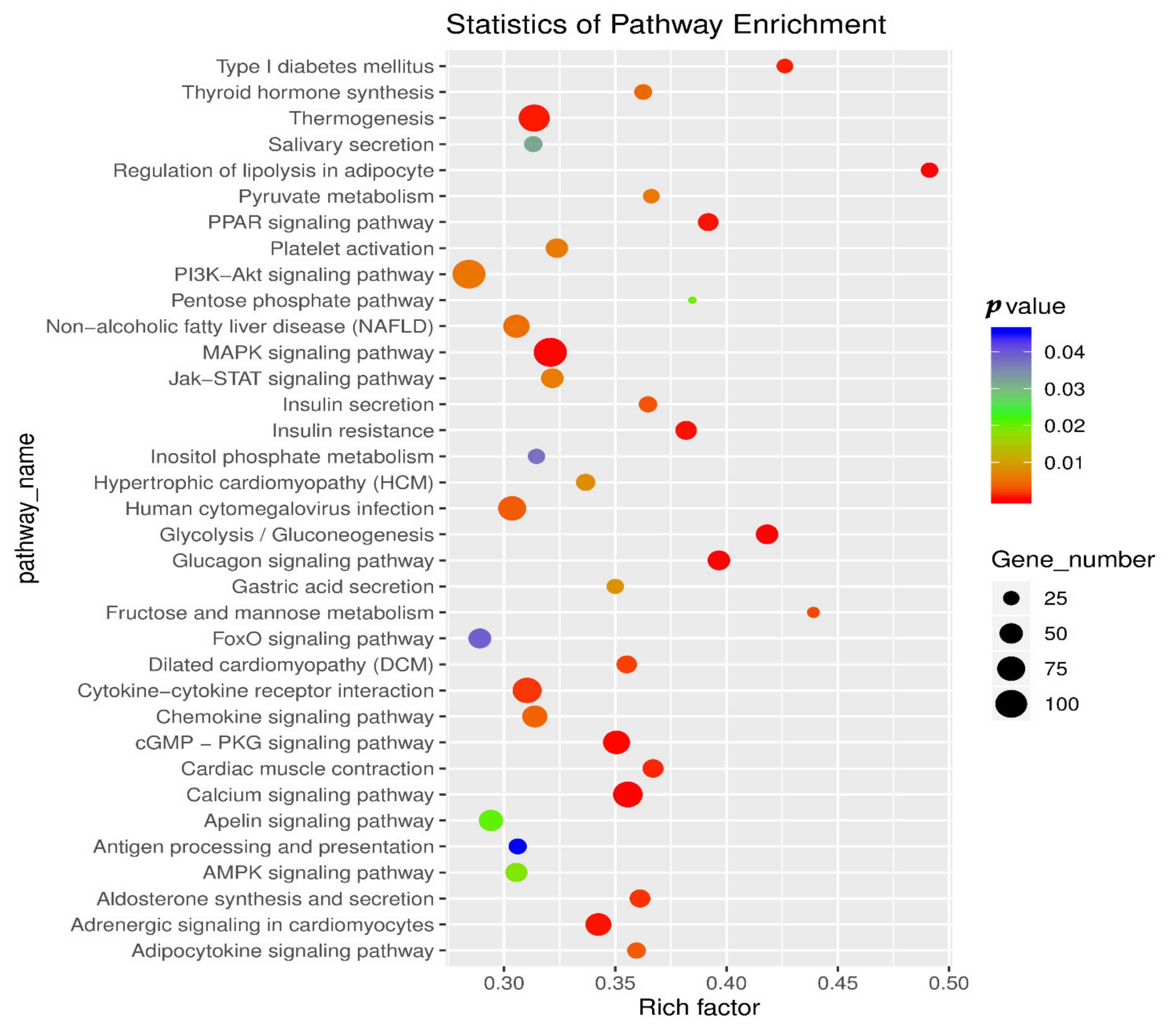
3.4. KEGG Pathway Analysis in Muscle and Adipose Tissue
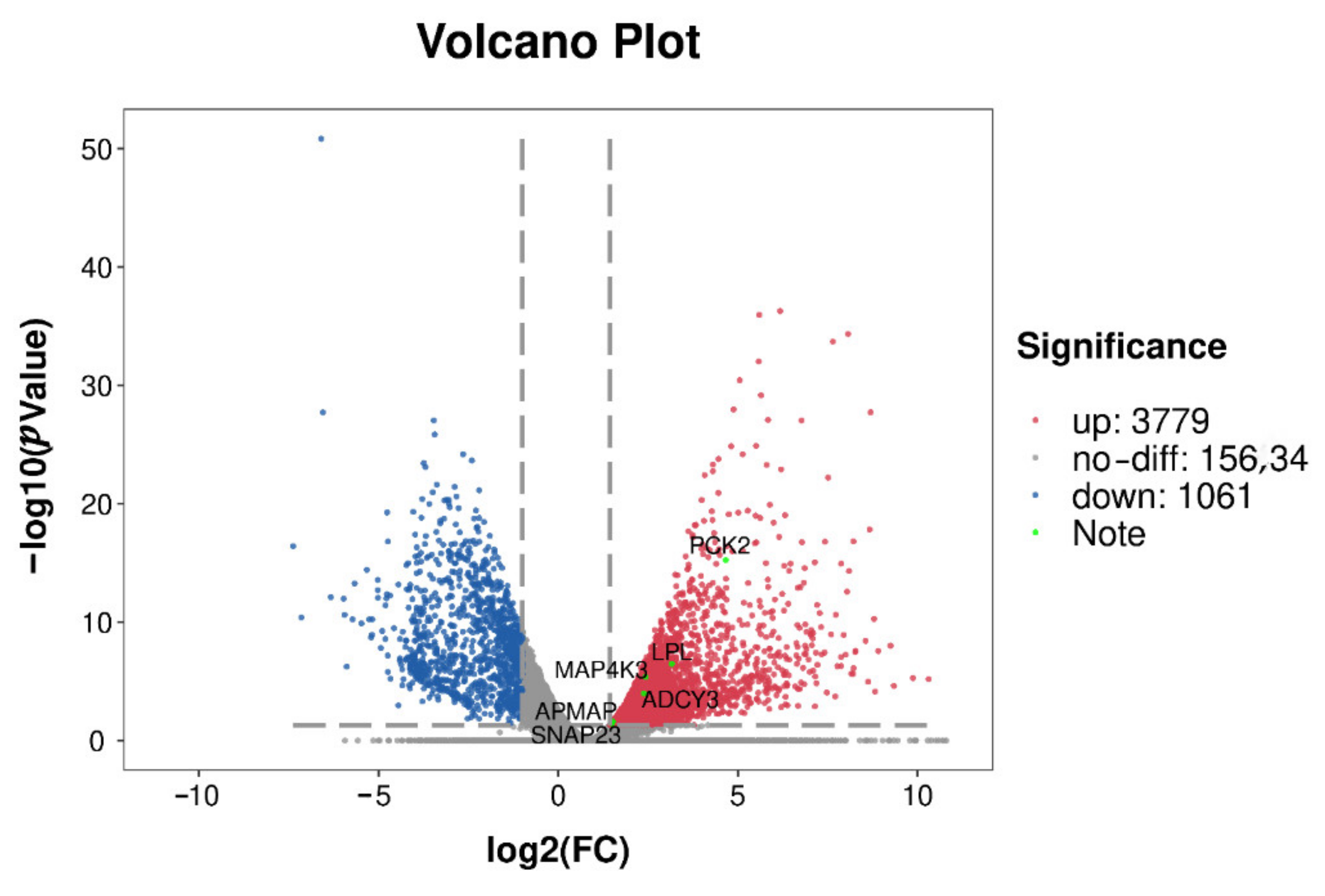
3.5. Gene Screening and Overview of m6A-Modified Genes
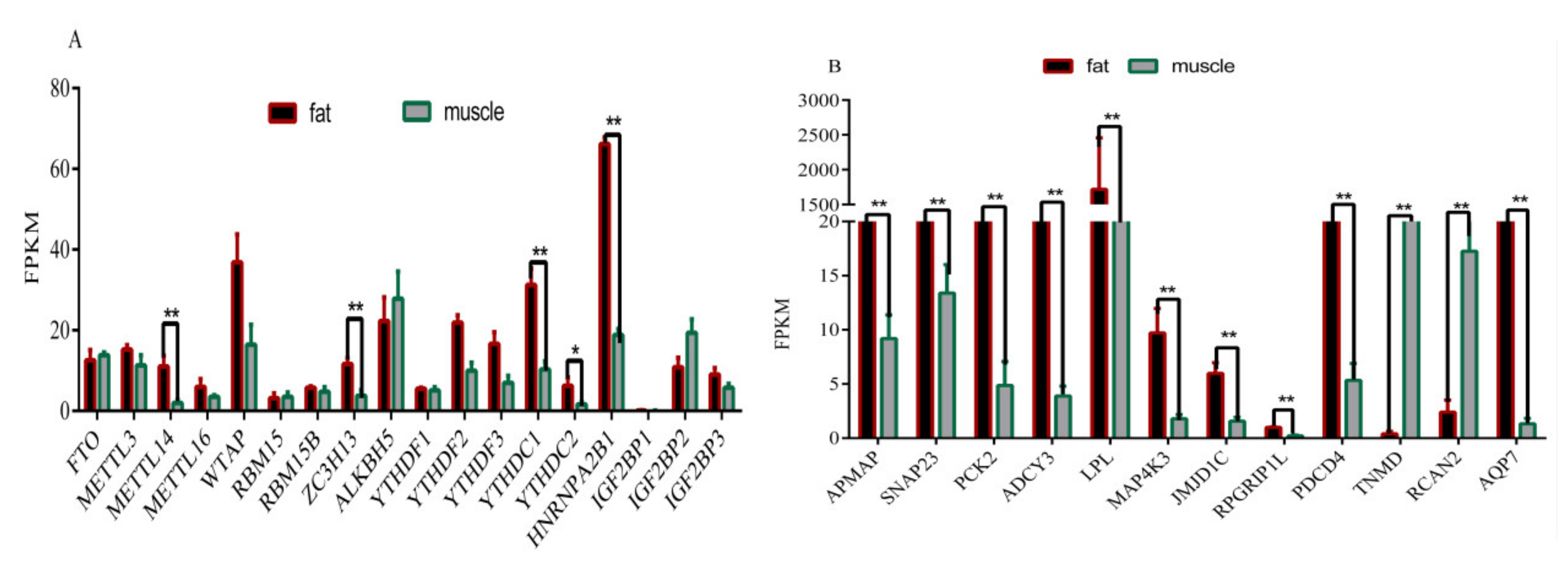
3.6. Overview of Differentially Expressed of Methylase Genes and Genes Related to Fat Deposition and Meat Quality in Muscle and Adipose Tissue Samples
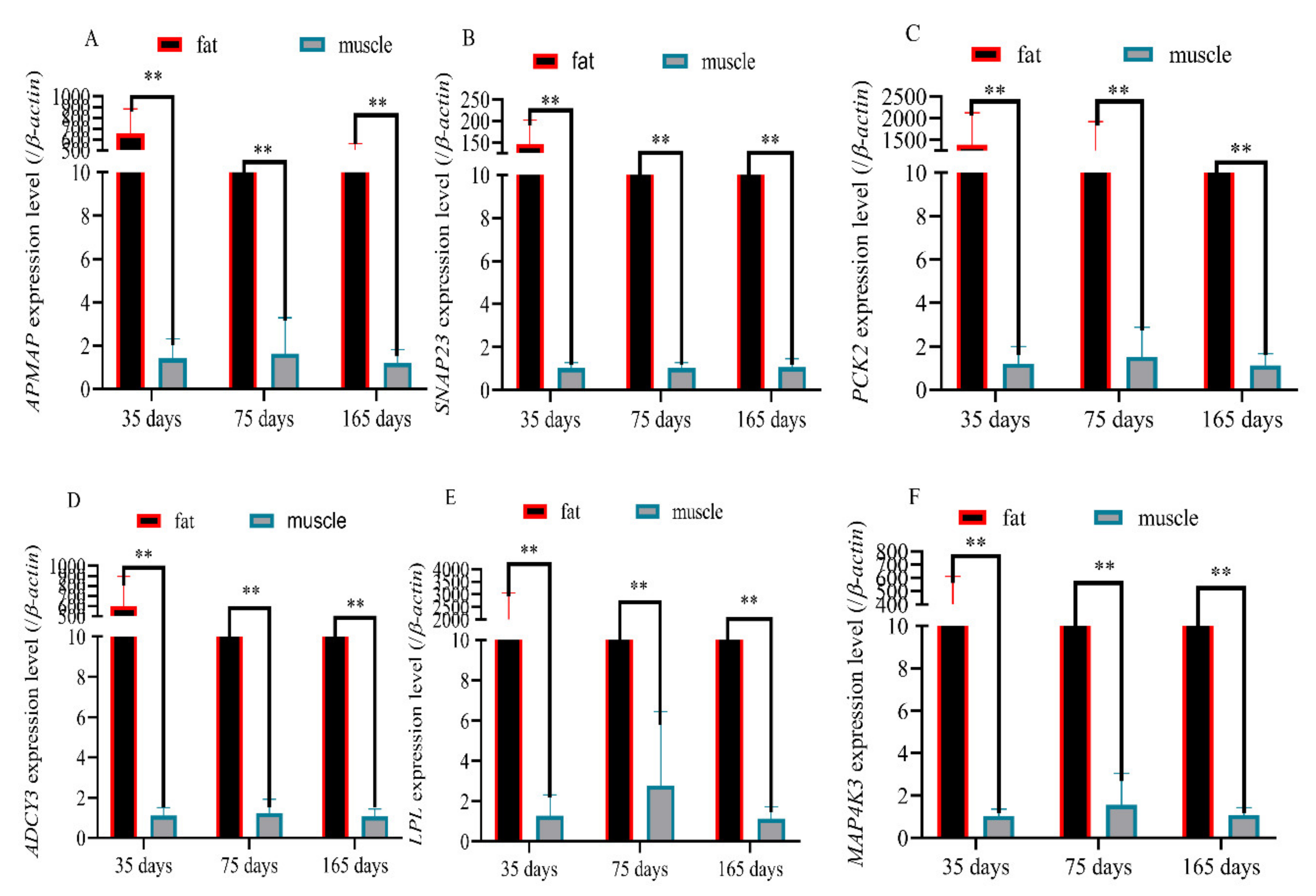
3.7. Validation of Six Randomly Genes Related to Fat Deposition and Meat Quality by RT-qPCR
3.8. Validation of Six Randomly Genes Related to Fat Deposition and Meat Quality by RT-qPCR
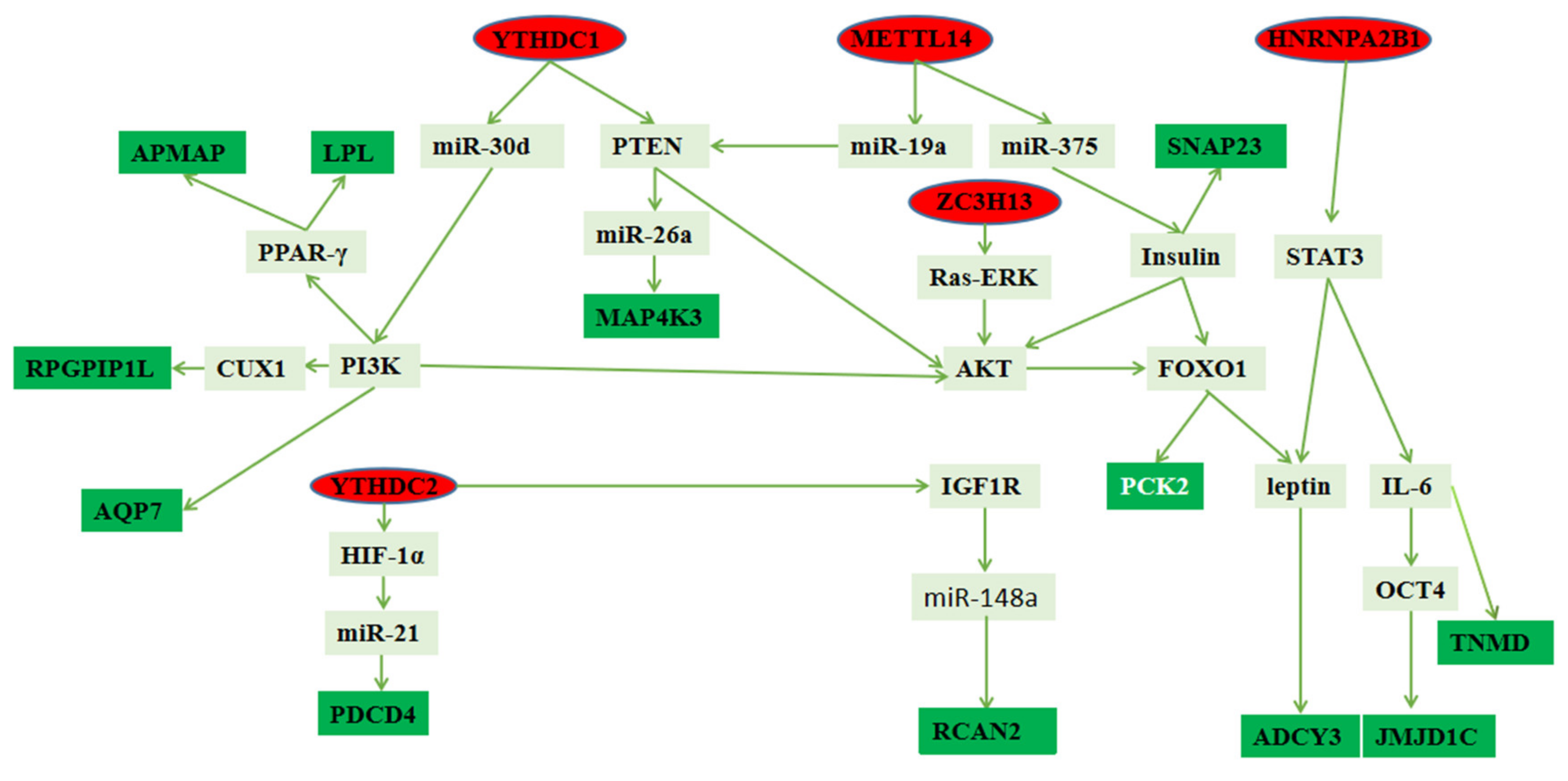
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Luo, G.; Mu, J.; Wang, S.; Dong, X.; Ren, Z. Association of blood APMAP content and meat quality trait in Rex rabbits. Anim. Biotechnol. 2021, 30, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Barnett, R. Obesity. Lancet 2005, 365, 1843–1851. [Google Scholar] [CrossRef]
- Fu, Y.; Dominissini, D.; Rechavi, G.; He, C. Gene expression regulation mediated through reversible m6A RNA methylation. Nat. Rev. Genet. 2014, 15, 293–306. [Google Scholar] [CrossRef]
- Wu, R.; Jiang, D.; Wang, Y.; Wang, X. N(6)-Methyladenosine (m(6)A) Methylation in mRNA with A Dynamic and Reversible Epigenetic Modification. Mol. Biotechnol. 2016, 58, 450–459. [Google Scholar] [CrossRef]
- Saletore, Y.; Meyer, K.; Korlach, J.; Vilfan, I.D.; Jaffrey, S.; Mason, C.E. The birth of the Epitranscriptome: Deciphering the function of RNA modifications. Genome Biol. 2012, 13, 175. [Google Scholar] [CrossRef] [Green Version]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 1, 885–887. [Google Scholar] [CrossRef]
- Yang, Y.; Hsu, P.J.; Chen, Y.S.; Yang, Y.G. Dynamic transcriptomic m(6)A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, S.; Mumbach, M.; Jovanovic, M.; Wang, T.; Maciag, K.; Bushkin, G.G.; Mertins, P.; Ter-Ovanesyan, D.; Habib, N.; Cacchiarelli, D. Perturbation of m6A Writers Reveals Two Distinct Classes of mRNA Methylation at Internal and 5′ Sites. Cell Rep. 2014, 8, 284–296. [Google Scholar] [CrossRef] [Green Version]
- Batista, P.M.B.; Wang, J.; Qu, K.; Zhang, J.; Li, L.; Bouley, D.; Lujan, E.; Haddad, B.; Daneshvar, K. m(6)A RNA modification controls cell fate transition in mammalian embryonic stem cells. Cell Stem Cell 2014, 15, 707–719. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; He, C. Fate by RNA methylation: M 6 A steers stem cell pluripotency. Genome Biol. 2015, 16, 41–43. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Yang, Y.; Sun, B.; Shi, Y.; Yang, X.; Xiao, W.; Hao, Y.; Ping, X.; Chen, Y.; Wang, W.; et al. FTO-dependent demethylation of N6-methyladenosine regulates mRNA splicing and is required for adipogenesis. Cell Res. 2014, 24, 1403–1419. [Google Scholar] [CrossRef]
- Merkestein, M.; Laber, S.; Mcmurray, F.; Andrew, D.; Sachse, G.; Sanderson, J.; Li, M.; Usher, S.; Sellayah, D.; Ashcroft, F.M. FTO influences adipogenesis by regulating mitotic clonal expansion. Nat. Commun. 2015, 6, 6792–6801. [Google Scholar] [CrossRef] [Green Version]
- Ma, C.; Chang, M.; Lv, H.; Zhang, Z.; Zhang, W.; He, X.; Wu, G.; Zhao, S.; Zhang, Y.; Wang, D.; et al. RNA m(6)A methylation participates in regulation of postnatal development of the mouse cerebellum. Genome Biol. 2018, 19, 68. [Google Scholar] [CrossRef] [Green Version]
- Lin, Z.; Hsu, P.J.; Xing, X.; Fang, J.; Lu, Z.; Zou, Q.; Zhang, K.; Zhang, X.; Zhou, Y.; Zhang, T.; et al. Mettl3-/Mettl14-mediated mRNA N6-methyladenosine modulates murine spermatogenesis. Cell Res. 2017, 27, 1216–1230. [Google Scholar] [CrossRef]
- Xu, K.; Yang, Y.; Feng, G.H.; Sun, B.F.; Chen, J.Q.; Li, Y.F.; Chen, Y.S.; Zhang, X.X.; Wang, C.X.; Jiang, L.Y. Mettl3-mediated m6A regulates spermatogonial differentiation and meiosis initiation. Cell Res. 2017, 27, 1100–1114. [Google Scholar] [CrossRef] [Green Version]
- Geula, S.; Moshitch-Moshkovitz, S.; Dominissini, D.; Mansour, A.A.; Kol, N.; Salmon-Divon, M.; Hershkovitz, V.; Peer, E.; Mor, N.; Manor, Y.S. m6A mRNA methylation facilitates resolution of nave pluripotency toward differentiation. Science 2015, 347, 1002–1006. [Google Scholar] [CrossRef]
- Ping, X.-L.; Chen, T.; Wang, M.; Li, M.M.; Zhao, X. m(6)A RNA Methylation Is Regulated by MicroRNAs and Promotes Reprogramming to Pluripotency. Cell Stem Cell 2015, 16, 289–301. [Google Scholar]
- Luo, G.; Hu, S.; Lai, T.; Wang, J.; Lai, S. MiR-9-5p promotes rabbit preadipocyte differentiation by suppressing leptin gene expression. Lipids Health Dis. 2020, 19, 1–11. [Google Scholar] [CrossRef]
- Schmittgen, K.J.L.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME Suite: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Heinz, S.; Benner, C.; Spann, N.; Bertolino, E.; Lin, Y.C.; Laslo, P.; Cheng, J.X.; Murre, C.; Singh, H.; Glass, C.K. Simple Combinations of Lineage-Determining Transcription Factors Prime cis-Regulatory Elements Required for Macrophage and B Cell Identities—ScienceDirect. Mol. Cell 2010, 38, 576–589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robinson, M.D.; Mccarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Biogeosciences 2010, 26, 139–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef]
- Meyer, K.D.; Saletore, Y.; Olivier, Z.P. Comprehensive Analysis of mRNA Methylation Reveals Enrichment in 3′ UTRs and near Stop Codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.Z.; Macqueen, A.; Zheng, G.; Duan, H.; Dore, L.C.; Lu, Z.; Liu, J.; Chen, K.; Jia, G.; Bergelson, J. Unique features of the m6A methylome in Arabidopsis thaliana. Nat. Commun. 2013, 5, 5630–5647. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, X.; Li, C.; Hu, S.; Yu, J.; Song, S. Transcriptome-wide N6-methyladenosine profiling of rice callus and leaf reveals the presence of tissue-specific competitors involved in selective mRNA modification. RNA Biol. 2014, 11, 1180–1188. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Zheng, Y.; Guo, D.; Zhang, X.; Bai, W. m6A Methylation Analysis of Differentially Expressed Genes in Skin Tissues of Coarse and Fine Type Liaoning Cashmere Goats. Front. Genet. 2020, 10, 1318. [Google Scholar] [CrossRef]
- Masatoshi, K.; Mitsuru, O.; Takayoshi, S.; Motoharu, A.; Toshihiro, U.; Aya, I.; Naoki, K.; Yukiko, O.; Naoto, K.; Ryo, S. The RNA methyltransferase complex of WTAP, METTL3, and METTL14 regulates mitotic clonal expansion in adipogenesis. Mol. Cell. Biol. 2018, 38, 116–118. [Google Scholar]
- Zhang, Z.; Wang, Q.; Zhao, X.; Shao, L.; Xu, R. YTHDC1 mitigates ischemic stroke by promoting Akt phosphorylation through destabilizing PTEN mRNA. Cell Death Dis. 2020, 11, 977. [Google Scholar] [CrossRef]
- He, H.; Cai, M.; Zhu, J.; Xiao, W.; Liu, B.; Shi, Y.; Yang, X.; Liang, X.; Zheng, T.; Hu, S. miR-148a-3p promotes rabbit preadipocyte differentiation by targeting PTEN. Vitr. Cell. Dev. Biol. Anim. 2018, 54, 241–249. [Google Scholar] [CrossRef]
- He, J.J.; Li, Z.; Rong, Z.X.; Gao, J.; Sun, L.Q. m6A Reader YTHDC2 Promotes Radiotherapy Resistance of Nasopharyngeal Carcinoma via Activating IGF1R/AKT/S6 Signaling Axis. Front. Oncol. 2020, 10, 1166. [Google Scholar] [CrossRef]
- Yun, S.J.; Kim, E.K.; Tucker, D.F.; Kim, C.D.; Birnbaum, M.J.; Bae, S.S. Isoform-specific regulation of adipocyte differentiation by Akt/protein kinase Balpha. Biochem. Biophys. Res. Commun. 2008, 371, 138–143. [Google Scholar] [CrossRef]
- Hu, Y.; Sun, Z.; Deng, J.; Hu, B.; Yan, W.; Wei, H.; Jiang, J. Splicing factor hnRNPA2B1 contributes to tumorigenic potential of breast cancer cells through STAT3 and ERK1/2 signaling pathway. Tumour Biol. 2017, 39, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Zhou, Y.; Lei, W.; Zhang, K.; Shi, J.; Hu, Y.; Shu, G.; Song, J. Signal transducer and activator of transcription 3 (STAT3) regulates adipocyte differentiation via peroxisome-proliferator-activated receptor γ (PPARγ). Biol. Cell 2010, 102, 1–12. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, Y.; Li, Z.; Zhiqing, Z.; Wang, P. MicroRNA-19a functions as an oncogene by regulating PTEN/AKT/pAKT pathway in myeloma. Leuk. Lymphoma 2017, 58, 932–940. [Google Scholar] [CrossRef]
- Huse, J.T.; Brennan, C.; Hambardzumyan, D.; Wee, B. The PTEN-regulating microRNA miR-26a is amplified in high-grade glioma and facilitates gliomagenesis in vivo. Genes Dev. 2009, 23, 1327–1337. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Yang, C.; Yang, Y. MicroRNA-26a inhibits the growth and invasiveness of malignant melanoma and directly targets on MITF gene. Cell Death Discov. 2017, 3, 17028. [Google Scholar] [CrossRef]
- Poy, M.N.; Hausser, J.; Trajkovski, M.; Braun, M.; Collins, S.; Rorsman, P.; Zavoian, M.; Stoffel, M. miR-375 maintains normal pancreatic α- and β-cell mass. Proc. Natl. Acad. Sci. USA 2009, 106, 5813–5818. [Google Scholar] [CrossRef] [Green Version]
- Wasik, A.A.; Dumont, V.; Tienari, J.; Nyman, T.A.; Fogarty, C.L.; Forsblom, C.; Lehto, M.; Lehtonen, E.; Groop, P.; Lehtonen, S. Septin 7 reduces nonmuscle myosin IIA activity in the SNAP23 complex and hinders GLUT4 storage vesicle docking and fusion—ScienceDirect. Exp. Cell Res. 2017, 350, 336–348. [Google Scholar] [CrossRef]
- Zhu, D.; Zhou, J.; Zhao, J.; Jiang, G.; Dong, M. ZC3H13 suppresses colorectal cancer proliferation and invasion via inactivating Ras–ERK signaling. J. Cell. Physiol. 2019, 234, 8899–8907. [Google Scholar] [CrossRef] [PubMed]
- Cheung, M.; Sharma, A.; Madhunapantula, S.V.; Robertson, G.P. Akt3 and Mutant V600EB-Raf Cooperate to Promote Early Melanoma Development. Cancer Res. 2008, 68, 3429–3439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Valades, A.G.; Mendez-Lucas, A.; Vidal-Alabro, A.; Blasco, F.X.; Chillon, M.; Bartrons, R.; Bermudez, J.; Perales, J.C. Pck1 Gene Silencing in the Liver Improves Glycemia Control, Insulin Sensitivity, and Dyslipidemia in db/db Mice. Diabetes 2008, 57, 2199–2210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitamura, T.; Feng, Y.; Kitamura, Y.I.; Chua, S.C.; Xu, A.W.; Barsh, G.S.; Rossetti, L.; Accili, D. Forkhead protein FoxO1 mediates Agrp-dependent effects of leptin on food intake. Nat. Med. 2006, 12, 534–540. [Google Scholar] [CrossRef] [PubMed]
- Michael, J.K.; Bradford, B.L.; Alastair, S.G. Melanocortin-4 receptor-regulated energy homeostasis. Nat. Neurosci. 2016, 19, 206–219. [Google Scholar]
- Hou, Y.; Zhang, Q.; Pang, W.; Hou, L.; Meng, X. YTHDC1-mediated augmentation of miR-30d in repressing pancreatic tumorigenesis via attenuation of RUNX1-induced transcriptional activation of Warburg effect. Cell Death Differ. 2021, 28, 3105–3124. [Google Scholar] [CrossRef]
- Ye, C.; Yu, X.; Liu, X.; Dai, M.; Zhang, B. Erratum: miR-30d inhibits cell biological progression of Ewing’s sarcoma by suppressing the MEK/ERK and PI3K/Akt pathways in vitro. Oncol. Lett. 2021, 21, 133. [Google Scholar] [CrossRef]
- Mei, G.; Fu, C.; Lin, T.; Peng, Y.; Li, W.; Zhu, X.; Ling, L.; Chen, Y. Apelin-13 Decreases Lipid Storage in Hypertrophic Adipocytes In Vitro Through the Upregulation of AQP7 Expression by the PI3K Signaling Pathway. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2014, 20, 1345–1352. [Google Scholar]
- Ripka, S.; Neesse, A.; Riedel, J.; Bug, E.; Aigner, A.; Poulsom, R.; Fulda, S.; Neoptolemos, J.; Greenhalf, W.; Barth, P. CUX1: Target of Akt signalling and mediator of resistance to apoptosis in pancreatic cancer. Gut 2010, 59, 1101–1110. [Google Scholar] [CrossRef]
- Stratigopoulos, G.; Leduc, C.A.; Cremona, M.L.; Chung, W.K.; Leibel, R.L. Cut-like Homeobox 1 (CUX1) Regulates Expression of the Fat Mass and Obesity-associated and Retinitis Pigmentosa GTPase Regulator-interacting Protein-1-like (RPGRIP1L) Genes and Coordinates Leptin Receptor Signaling. J. Biol. Chem. 2011, 286, 2155–2170. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Gollapudi, S.; Su, H.; Gupta, S. Leptin Activates Human B Cells to Secrete TNF-α, IL-6, and IL-10 via JAK2/STAT3 and p38MAPK/ERK1/2 Signaling Pathway. J. Clin. Immunol. 2011, 31, 472–478. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Kang, J.W.; Song, X.; Kim, B.K.; Yoo, Y.D.; Kwon, Y.T.; Lee, Y.J. Role of the IL-6-JAK1-STAT3-Oct-4 pathway in the conversion of non-stem cancer cells into cancer stem-like cells. Cell. Signal. 2013, 25, 961–969. [Google Scholar] [CrossRef] [Green Version]
- Katoh, M.; Katoh, M. Comparative integromics on JMJD1C gene encoding histone demethylase: Conserved POU5F1 binding site elucidating mechanism of JMJD1C expression in undifferentiated ES cells and diffuse-type gastric cancer. Int. J. Oncol. 2007, 31, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Gaut, L.; Duprez, D. Tendon development and diseases. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 5–23. [Google Scholar] [CrossRef] [Green Version]
- Tanabe, A.; Tanikawa, K.; Tsunetomi, M.; Takai, K.; Ikeda, H.; Konno, J.; Torigoe, T.; Maeda, H.; Kutomi, G.; Okita, K. RNA helicase YTHDC2 promotes cancer metastasis via the enhancement of the efficiency by which HIF-1α mRNA is translated. Cancer Lett. 2016, 376, 34–42. [Google Scholar] [CrossRef]
- Jia, Z.; Lian, W.; Shi, H.; Cao, C.; Han, S.; Wang, K.; Li, M.; Zhang, X. Ischemic Postconditioning Protects Against Intestinal Ischemia/Reperfusion Injury via the HIF-1α/miR-21 Axis. Sci. Rep. 2017, 7, 16190. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z. miR-21 promotes migration and invasion by the miR-21-PDCD4-AP-1 feedback loop in human hepatocellular carcinoma. Oncol. Rep. 2012, 27, 1660–1668. [Google Scholar] [CrossRef] [Green Version]
- Xu, Q.; Liu, L.Z.; Yin, Y.; He, J.; Li, Q.; Qian, X.; You, Y.; Lu, Z.; Peiper, S.C.; Shu, Y. Regulatory circuit of PKM2/NF-κB/miR-148a/152-modulated tumor angiogenesis and cancer progression. Oncogene 2015, 34, 5482–5493. [Google Scholar] [CrossRef]
- Ma, X.; Zhu, G.; Jiao, T.; Shao, F. Effects of circular RNA Ttc3/miR-148a/Rcan2 axis on inflammation and oxidative stress in rats with acute kidney injury induced by sepsis. Life Sci. 2021, 272, 119233. [Google Scholar] [CrossRef]
- Bogner-Strauss, J.G.; Prokesch, A.; Sanchez-Cabo, F.; Rieder, D.; Hackl, H.; Duszka, K.; Krogsdam, A.; di Camillo, B.; Walenta, E.; Klatzer, A.; et al. Reconstruction of gene association network reveals a transmembrane protein required for adipogenesis and targeted by PPARγ. Cell. Mol. Life Ences 2010, 67, 4049–4064. [Google Scholar] [CrossRef]
- Boström, P.; Andersson, L.; Rutberg, M.; Perman, J.; Lidberg, U.; Johansson, B.R.; Fernandez-Rodriguez, J.; Ericson, J. SNARE proteins mediate fusion between cytosolic lipid droplets and are implicated in insulin sensitivity. Nat. Cell Biol. 2007, 9, 1286–1293. [Google Scholar] [CrossRef] [PubMed]
- Modaressi, S.; Brechtel, K.; Christ, B.; Jungermann, K.; Localization, C. Human mitochondrial phosphoenolpyruvate carboxykinase 2 gene. Structure, chromosomal localization and tissue-specific expression. Biochem. J. 1998, 333, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Pitman, J.L.; Wheeler, M.C.; Lloyd, D.J.; Walker, J.R.; Glynne, R.J.; Gekakis, N. A Gain-of-Function Mutation in Adenylate Cyclase 3 Protects Mice from Diet-Induced Obesity. PLoS ONE 2014, 9, e110226. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Shen, C.; Ahmed, M.S. Adenylate cyclase 3: A new target for anti-obesity drug development. Obes. Rev. 2016, 17, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Sorisky, A. From Preadipocyte to Adipocyte: Differentiation-Directed Signals of Insulin from the Cell Surface to the Nucleus. Crc Crit. Rev. Clin. Lab. Sci. 1999, 36, 1–34. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Wang, J.; Xie, W.; Lyu, Q.; Wu, J.; He, J.; Qiu, W.; Xu, N.; Zhang, Y. MiR-378a-3p enhances adipogenesis by targeting mitogen-activated protein kinase 1. Biochem. Biophys. Res. Commun. 2015, 457, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Viscarra, J.A.; Wang, Y.; Nguyen, H.P.; Choi, Y.G.; Sul, H.S. Histone demethylase JMJD1C is phosphorylated by mTOR to activate de novo lipogenesis. Nat. Commun. 2020, 11, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.; Cantagrel, V.; Mariani, L.; Serre, V.; Lee, J.; Elkhartoufi, N.; de Lonlay, P.; Desguerre, I.; Munnich, A.; Boddaert, N.; et al. Identification of a novel ARL13B variant in a Joubert syndrome-affected patient with retinal impairment and obesity. Eur. J. Hum. Genet. 2015, 23, 621–627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Dong, Z.; Liu, X.; Song, X.; Song, Q.; Shang, Q.; Jiang, Y.; Guo, C.; Zhang, L. Programmed cell death-4 deficiency prevents diet-induced obesity, adipose tissue inflammation, and insulin resistance. Diabetes 2013, 62, 4132–4143. [Google Scholar] [CrossRef] [Green Version]
- Tolppanen, A.M.; Pulkkinen, L.; Kolehmainen, M.; Schwab, U.; Lindström, J.; Tuomilehto, J.; Uusitupa, M. Tenomodulin is Associated with Obesity and Diabetes Risk: The Finnish Diabetes Prevention Study. Obesity 2012, 15, 1082–1088. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.Y.; Hayashi, Y.; Xu, S.; Kanou, Y.; Takagishi, Y.; Tang, Y.P.; Murata, Y. Inactivation of the Rcan2 Gene in Mice Ameliorates the Age- and Diet-Induced Obesity by Causing a Reduction in Food Intake. PLoS ONE 2011, 6, e14605. [Google Scholar] [CrossRef]
- Miyauchi, T.; Yamamoto, H.; Abe, Y.; Yoshida, G.J.; Rojek, A.; Sohara, E.; Uchida, S.; Yasui, M. Dynamic subcellular localization of aquaporin-7 in white adipocytes. Febs Lett. 2015, 589, 608–614. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Primer Sequence | Temperature (°C) | Product Size (bp) | Gene ID |
|---|---|---|---|---|
| APMAP | 5’-GCTGCTGGATTCTCCCATAG-3′ | 60 | 163 | 100339857 |
| 5’-AAACATCACGTCCCCGATAT-3′ | ||||
| β-actin | 5’-GGAGATCGTGCGGGACAT-3′ | 61.4 | 318 | 100009272 |
| 5’-GTTGAAGGTGGTCTCGTGGAT-3′ | ||||
| SNAP23 | 5’-CCTGGCAATGTGGTGTCTAA-3′ | 59.5 | 250 | 100008776 |
| 5’-TGGTGTCAGCCTTTTCTGTAAT-3′ | ||||
| PCK2 | 5’-AACAGGAGGTGCGTGACATT-3′ | 60.2 | 250 | 100144327 |
| 5’-GGGACAGGGAGTGTGAGAAG-3′ | ||||
| ADCY3 | 5’-TGGGCGTCATGTCCTACTAC-3′ | 60 | 238 | 100343958 |
| 5’-ACATTCTCGTGGCGGTACAT-3′ | ||||
| LPL | 5’-GACATTGGGGAGTTGCTGAT-3′ | 60.5 | 214 | 100340171 |
| 5’ACTTGTCGTGGCATTTCACA-3′ | ||||
| MAP4K3 | 5’-ATGTGGGGCACTCCAAACTA-3′ | 59.5 | 182 | 100356354 |
| 5’-TGAAGTCTCGCCCTCTACTG-3′ |
| Sample | Raw_Reads | Valid_Reads | Valid% | Q20% | Q30% | GC% |
|---|---|---|---|---|---|---|
| Fat1_IP | 81562394 | 79590262 | 89.94 | 98.07 | 94.17 | 49.20 |
| Fat2_IP | 74268656 | 72697900 | 90.48 | 98.02 | 94.08 | 49.93 |
| Fat3_IP | 75635076 | 74194910 | 90.77 | 97.92 | 93.90 | 50.64 |
| Mus1_IP | 101998102 | 100309020 | 91.02 | 98.17 | 94.45 | 53.73 |
| Mus2_IP | 100858460 | 98910896 | 90.69 | 98.09 | 94.25 | 53.14 |
| Mus3_IP | 77539934 | 76213194 | 90.62 | 98.08 | 94.24 | 53.00 |
| Fat1_input | 67621338 | 66671322 | 90.59 | 98.09 | 94.16 | 48.59 |
| Fat2_input | 72342384 | 71424132 | 90.48 | 98.17 | 94.36 | 49.85 |
| Fat3_input | 72298544 | 71241220 | 90.17 | 98.16 | 94.40 | 51.05 |
| Mus1_input | 78891496 | 78053538 | 91.05 | 98.18 | 94.45 | 52.35 |
| Mus2_input | 102167202 | 100877896 | 90.49 | 98.04 | 94.18 | 53.68 |
| Mus3_input | 92062236 | 90789550 | 90.64 | 98.25 | 94.62 | 53.62 |
| Sample | Valid Reads | Mapped Reads | Unique Mapped Reads | Multi Mapped Reads |
|---|---|---|---|---|
| Fat1_IP | 78626498 | 71401305 (90.81%) | 50564672 (64.31%) | 20836633 (26.50%) |
| Fat2_IP | 71989454 | 65126746 (90.47%) | 49953050 (69.39%) | 15173696 (21.08%) |
| Fat3_IP | 73616646 | 65572313 (89.07%) | 49524600 (67.27%) | 16047713 (21.80%) |
| Mus1_IP | 100013482 | 83125913 (83.11%) | 58211564 (58.20%) | 24914349 (24.91%) |
| Mus2_IP | 98559478 | 81835114 (83.03%) | 59331136 (60.20%) | 22503978 (22.83%) |
| Mus3_IP | 75865960 | 62874564 (82.88%) | 46707280 (61.57%) | 16167284 (21.31%) |
| Fat1_input | 65033698 | 60764764 (93.44%) | 43533958 (66.94%) | 17230806 (26.50%) |
| Fat2_input | 69325074 | 64450485 (92.97%) | 48831496 (70.44%) | 15618989 (22.53%) |
| Fat3_input | 70284120 | 64559775 (91.86%) | 48476357 (68.97%) | 16083418 (22.88%) |
| Mus1_input | 77296586 | 69248163 (89.59%) | 48632494 (62.92%) | 20615669 (26.67%) |
| Mus2_input | 99957966 | 88668893 (88.71%) | 62516927 (62.54%) | 26151966 (26.16%) |
| Mus3_input | 65033698 | 60764764 (93.44%) | 43533958 (66.94%) | 17230806 (26.50%) |
| Gene Name | log2(fc) | Methylation Regulation | Chromosome | Peak Region | Peak Star | Peak End | p-Value |
|---|---|---|---|---|---|---|---|
| APMAP | 1.52 | Hypo-methylation | 65 | 3’ UTR | 1,160,678 | 1,161,123 | 1 × 10−42 |
| SNAP23 | 1.49 | Hypo-methylation | 17 | Exon | 29,652,649 | 29,656,542 | 5.01 × 10−37 |
| PCK2 | 4.67 | Hypo-methylation | 17 | 3’ UTR | 44,153,721 | 44,154,226 | 1.58 × 10−33 |
| ADCY3 | 2.38 | Hypo-methylation | 2 | Exon | 173,934,224 | 173,934,819 | 5.01 × 10−26 |
| LPL | 3.16 | Hypo-methylation | 15 | 5’ UTR | 4,554,062 | 4,554,301 | 0.0041 |
| MAP4K3 | 2.43 | Hypo-methylation | 2 | 5’ UTR | 146,895,441 | 146,925,862 | 0.008 |
| JMJD1C | 1.93 | Hypo-methylation | 18 | Exon | 23,014,029 | 23,014,960 | 0.00017 |
| RPGRIP1L | 2.18 | Hypo-methylation | 5 | 5’ UTR | 10,022,614 | 10,027,508 | 0.0093 |
| PDCD4 | 2.1 | Hyper-methylation | 18 | 5’ UTR | 58,499,213 | 58,499,903 | 0.012 |
| TNMD | −6.59 | Hyper-methylation | 22 | Exon | 88,793,707 | 88,793,990 | 0.05 |
| RCAN2 | −2.85 | Hypo-methylation | 12 | Exon | 35,531,621 | 35,531,680 | 0.027 |
| AQP7 | 5.58 | Hypo-methylation | 1 | 3’ UTR | 20,078,653 | 20,078,802 | 0.022 |
| Gene Name | Gene ID | M6A Regulation | Gene Regulation | Block Count | Block Sizes | Block Starts | Distance To TSS |
|---|---|---|---|---|---|---|---|
| APMAP | 100339857 | Down | up | 1 | 446 | 0 | 41,647 |
| SNAP23 | 100008776 | Down | up | 3 | 27, 145, 37 | 0, 2495, 3857 | 30,071 |
| PCK2 | 100144341 | Down | up | 1 | 506 | 0 | 9040 |
| ADCY3 | 100343958 | Down | up | 2 | 57, 94, | 0, 502 | 72,693 |
| LPL | 100340171 | Down | up | 1 | 240 | 0 | 30 |
| MAP4K3 | 100356354 | Down | up | 2 | 195, 15, | 0, 30407 | 0 |
| JMJD1C | 100350438 | Down | up | 2 | 92, 298 | 0, 634 | 262,502 |
| RPGRIP1L | 100354520 | Down | up | 4 | 58, 92, 44, 47 | 0, 2264, 2782, 4848 | 239 |
| PDCD4 | 100354557 | up | up | 2 | 54, 184 | 0, 507 | 0 |
| TNMD | 100125994 | up | Down | 2 | 72, 48 | 0, 236 | 0 |
| RCAN2 | 100343905 | Down | Down | 1 | 60 | 0 | 303,927 |
| AQP7 | 100350611 | Down | up | 1 | 150 | 0 | 11,448 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luo, G.; Wang, S.; Ai, Y.; Li, J.; Ren, Z. N6-Methyladenosine Methylome Profiling of Muscle and Adipose Tissues Reveals Methylase–mRNA Metabolic Regulatory Networks in Fat Deposition of Rex Rabbits. Biology 2022, 11, 944. https://doi.org/10.3390/biology11070944
Luo G, Wang S, Ai Y, Li J, Ren Z. N6-Methyladenosine Methylome Profiling of Muscle and Adipose Tissues Reveals Methylase–mRNA Metabolic Regulatory Networks in Fat Deposition of Rex Rabbits. Biology. 2022; 11(7):944. https://doi.org/10.3390/biology11070944
Chicago/Turabian StyleLuo, Gang, Shuhui Wang, Yaotian Ai, Jiapeng Li, and Zhanjun Ren. 2022. "N6-Methyladenosine Methylome Profiling of Muscle and Adipose Tissues Reveals Methylase–mRNA Metabolic Regulatory Networks in Fat Deposition of Rex Rabbits" Biology 11, no. 7: 944. https://doi.org/10.3390/biology11070944
APA StyleLuo, G., Wang, S., Ai, Y., Li, J., & Ren, Z. (2022). N6-Methyladenosine Methylome Profiling of Muscle and Adipose Tissues Reveals Methylase–mRNA Metabolic Regulatory Networks in Fat Deposition of Rex Rabbits. Biology, 11(7), 944. https://doi.org/10.3390/biology11070944